小麦穗顶部和基部小穗结实粒数的全基因组关联分析
2021-04-19马婕龙史雨刚曹亚萍孙黛珍王曙光
马婕龙,张 政,史雨刚,曹亚萍,孙黛珍,王曙光
(1.山西农业大学农学院,山西太谷030801;2.山西农业大学小麦研究所,山西临汾041000)
小麦(Triticum aestivumL.)作为主要粮食作物,为世界上1/3 以上的人口提供了能量来源[1-2]。随着人口不断增长,粮食短缺将继续是人类面临的难题,所以,提高籽粒产量仍是未来小麦育种的主要目标之一[3]。穗作为小麦地上部最后一个发育器官,它的发育情况将直接影响穗粒数,进而影响小麦籽粒产量[4]。而穗顶部和基部结实粒数作为穗粒数重要组成部分,对穗粒数影响很大,但在小麦育种中往往容易被忽视[5]。
小麦花序呈复穗状,多个互生的小穗在穗轴两侧交错排列,且每个小穗可分化出3~9 朵小花,但小穗上部小花由于发育时间短,往往会退化,而基部的2~5 朵小花则可以正常发育,同一穗中小花的发育顺序取决于小花穗位,先穗中部小花,其次为穗基部和顶部小花[6-7]。显然小麦穗部小花的发育并不是同步进行的,且最先发育的小花可以继续发育并形成籽粒,而后发育的小花更容易丧失生活力和生育力[8]。研究表明,小花的生育力与其在穗部位置密切相关[9],环境因素和栽培措施也会影响小花的分化和发育以及结实率[10]。
穗顶部和基部小花低生育率在禾谷类作物中是非常常见的现象,如玉米的“秃头”[11-12]。研究发现,在小麦中小穗结实粒数与穗粒数呈极显著正相关,穗顶部和基部小花低生育力会造成顶部和基部小穗结实率降低,将直接影响整穗穗粒数[13-14]。在水稻中已经发现影响穗顶部和基部小花发育的相关基因[15]。目前在小麦中关于穗顶部和基部结实粒数的相关研究报道较少,但是有较多关于不育小穗的研究。MA 等[16]通过利用重组自交系在13 条染色体上均检测到与小花不育相关的QTL 位点;XU 等[17]利用不同氮磷水平下生长的重组自交系群体,在2D 染色体上的Xwmc112~Xbarc168区间内检测到1 个与小花不育相关的QTL。
全基因组关联分析(Genome-wide association study,GWAS)是基于连锁不平衡,对多个个体的目标性状与全基因组范围内的标记进行关联分析,进而挖掘出与目标性状相关的基因,可用来鉴定与复杂性状相关联的基因变异[18]。
本研究通过分析236 份小麦种质资源在4 个环境下的穗顶部结实粒数和穗基部结实粒数,并与106 对SSR 标记进行关联分析,筛选出相关的优异等位变异,为分子标记辅助选择来提高小麦穗顶部结实粒数和基部结实粒数,进而提高小麦产量提供参考。
1 材料和方法
1.1 试验材料
本研究中所用的236 份小麦种质资源由我国北方冬小麦育成品种和骨干亲本组成,包括225 份国内品种、5 份国外品种以及6 份来源未知的品种。
1.2 试验方法
试验材料于2016 年9 月和2017 年9 月在山西农业大学农作站小麦试验基地种植,包括雨养(DS)和灌溉(WW)2 种水分处理,共4 种环境,即2017DS、2017WW、2018DS 和2018WW。其中,干旱条件为在小麦生长发育期间只依靠自然降雨补充水分,不增加人工灌溉;灌溉是指除自然降雨外,还根据田间墒情分别于越冬前、返青期、拔节期和灌浆中期增加人工灌溉。每个处理均为随机区组设计,重复3 次,播种时,每份材料挑选70 粒饱满无损伤的种子,均匀点播在行长2 m、行间距20 cm 的2 行小区内。
成熟时每个材料选取10 个长势一致的穗子,分别检测麦穗从顶部数第1、2、3 小穗结实粒数(分别简称顶1(T1)、顶2(T2)、顶3(T3))以及穗子从基部数第1、2、3 小穗结实粒数(分别简称基1(B1)、基2(B2)、基3(B3)),取平均值作为表型值。
1.3 数据分析
所用SSR 标记的多态性筛选以及群体结构分析参照张东等[19]的方法进行,所有SSR 位点的等位变异数、等位基因频率、多态性信息指数(PIC)参照LIU 等[20]的方法计算。参照EVANNO 等[21]利用K+Q 的模型进行亚群数量分析。使用TASSEL 3.0 软件中的混合线性模型(MLM),根据计算出的Q 值和K 值进行标记与表型性状数据的关联分析。群体表型数据利用SPSS 20.0 进行统计分析。
2 结果与分析
2.1 不同环境下小麦穗顶部和基部小穗结实粒数的变化
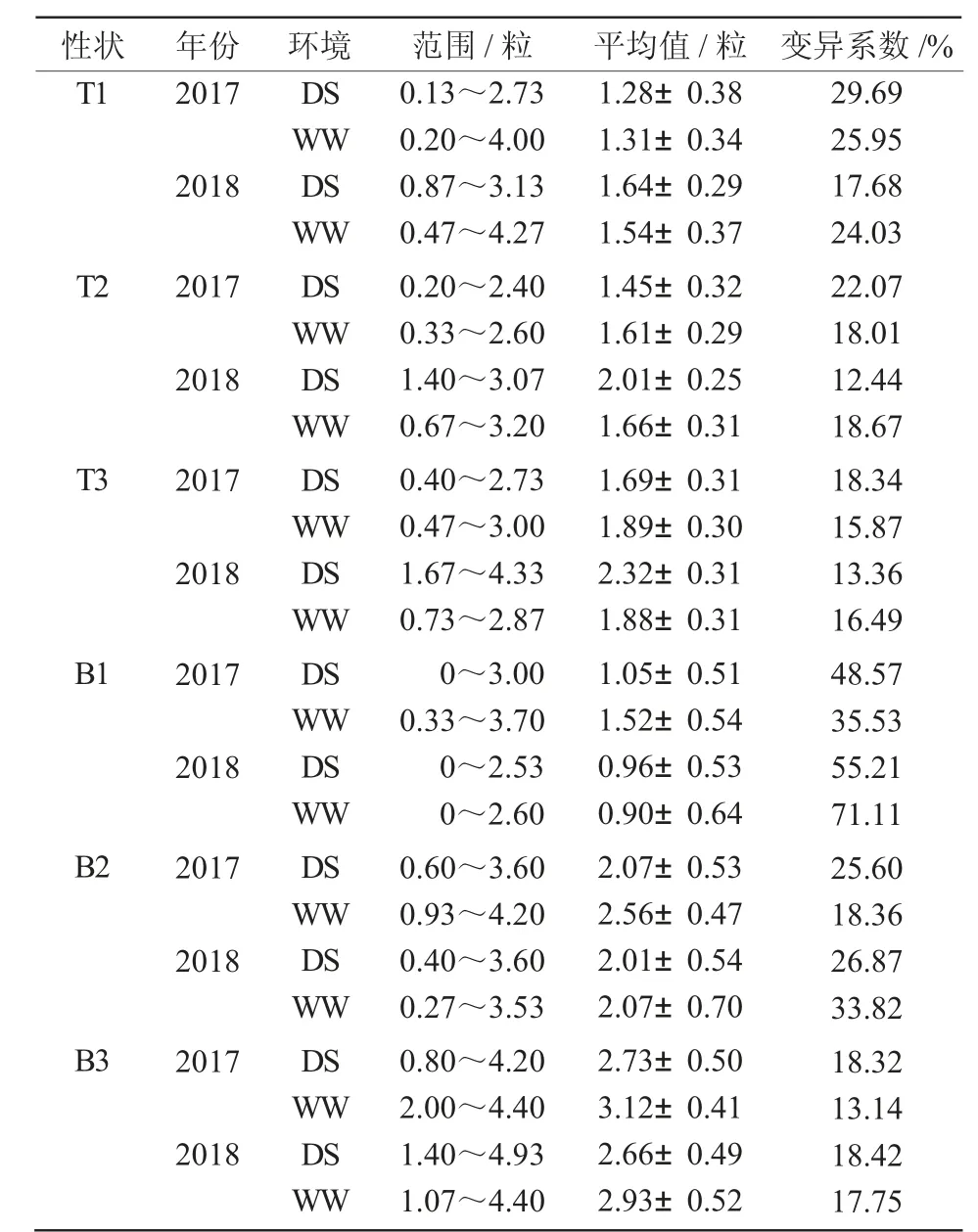
表1 不同环境下小麦穗顶部和基部小穗结实粒数的变异比较
通过调查2 a 共4 个环境下236 份小麦品种资源的穗顶部小穗结实粒数和基部小穗结实粒数,结果表明(表1),所有性状都存在较大的变异,穗顶部小穗结实粒数的变异系数为12.44%~29.69%,穗基部小穗结实粒数的变异系数为13.14%~71.11%。比较发现,穗顶部小穗结实粒数的变异系数大多在20%左右,整体要低于穗基部小穗结实粒数,穗基部小穗结实粒数在多个环境条件下的变异系数都要高于20%。
在雨养条件下,穗顶部第1、2、3 小穗结实粒数分别为0.13~3.13、0.20~3.07、0.40~4.33 粒,穗基部第1、2、3 小穗结实粒数分别为0~3.00、0.40~3.60、0.80~4.93 粒。在灌溉条件下,穗顶部第1、2、3 小穗结实粒数分别为0.20~4.27、0.33~3.20、0.47~3.00 粒,穗基部第1、2、3 小穗结实粒数分别为0~3.70、0.27~4.20、1.07~4.40 粒。结果表明,小麦穗顶部结实粒数和基部结实粒数在群体品种间存在较大的遗传变异。
2.2 小麦穗顶部和基部小穗结实粒数与标记间的关联分析
利用穗顶部小穗结实粒数和基部小穗结实粒数与SSR 标记进行全基因组关联分析,结果表明(表2),在P<0.01 水平下,共检测到6 个位点与穗顶部第1 个小穗结实粒数显著相关,表型解释率为5.87%~30.73%,分别位于染色体1A、1D、2B、3B、6B和6D 上,其中,标记Xgwm55(6D)关联到2 个环境,即17DS 和17WW,表型解释率分别为11.69%和20.62%;且除了Xgwm232(1D)和Xwmc361(2B)外,其他标记的表型解释率均高于10%,其中,Xgwm164(1A)的表型解释率为30.73%,P值为1.15×10-9,均高于其他位点。检测到9 个与穗顶部第2 个小穗结实粒数显著相关的位点,表型解释率为5.92%~20.56%,分别位于染色体2A、2D、3A、4D、5B 和5D 上,其中,Xgwm539(2D)和Xgwm484(2D)的表型解释率高于其他各显著关联位点,分别为20.56%和20.00%。检测到7 个与穗顶部第3 个小穗结实粒数显著相关的位点,表型解释率为5.73%~15.34%,分别位于染色体1A、2A、2D、3B、4B、5D 和6D 上,除了标记Xgwm102(2D)和Xgwm538(4B)外,其他标记的表型解释率均高于10%。与穗基部小穗结实粒数显著相关的位点共检测到5 个,其中与穗基部第1、2 个小穗结实粒数显著相关的位点各有1 个,分别为Xgwm55(6D)和Xgwm5(3A),表型解释率分别为11.50%和10.66%;与穗基部第3 个小穗结实粒数显著关联的位点有3 个,即Xgwm285(3B)、Xgwm161(3D)与Xcfe172(3D),表型解释率分别为14.96%、16.22%和6.84%。
2.3 小麦穗顶部和穗基部小穗结实粒数优异等位变异及其效应分析
对每个关联位点优异等位变异及其效应进行分析,结果表明(表3),与穗顶部第1 个小穗的结实粒数显著相关的6 个位点中,Xgwm132-6B128等位变异效应最大,可增加结实粒数0.14 粒;Xwmc361-2B216等位变异频率最高,为74.63%,可增加结实粒数0.02 粒。与穗顶部第2 个小穗的结实粒数显著相关的9 个位点中,Xgwm102-2D142等位变异效应最大,可增加结实粒数0.19 粒;Xgwm609-4D111等位变异频率最高,为47.32%,可增加结实粒数0.08 粒。与穗顶部第3 个小穗的结实粒数显著相关的7 个位点中,Xgwm102-2D142等位变异效应最大,可增加结实粒数0.20 粒;Xcfd266-5D165等位变异频率最高,为50.24%,可增加结实粒数0.04 粒。与穗基部小穗结实粒数显著相关的位点中,Xgwm55-6D128可增加基部第1 个小穗的结实粒数0.07 粒;等位变异频率最高的是Xgwm5-3A168,为31.22%,可增加穗基部第2 个小穗结实粒数0.17 粒;Xcfe172-3D144等位变异在18WW 环境下可增加穗基部第3 个小穗结实粒数0.34 粒。
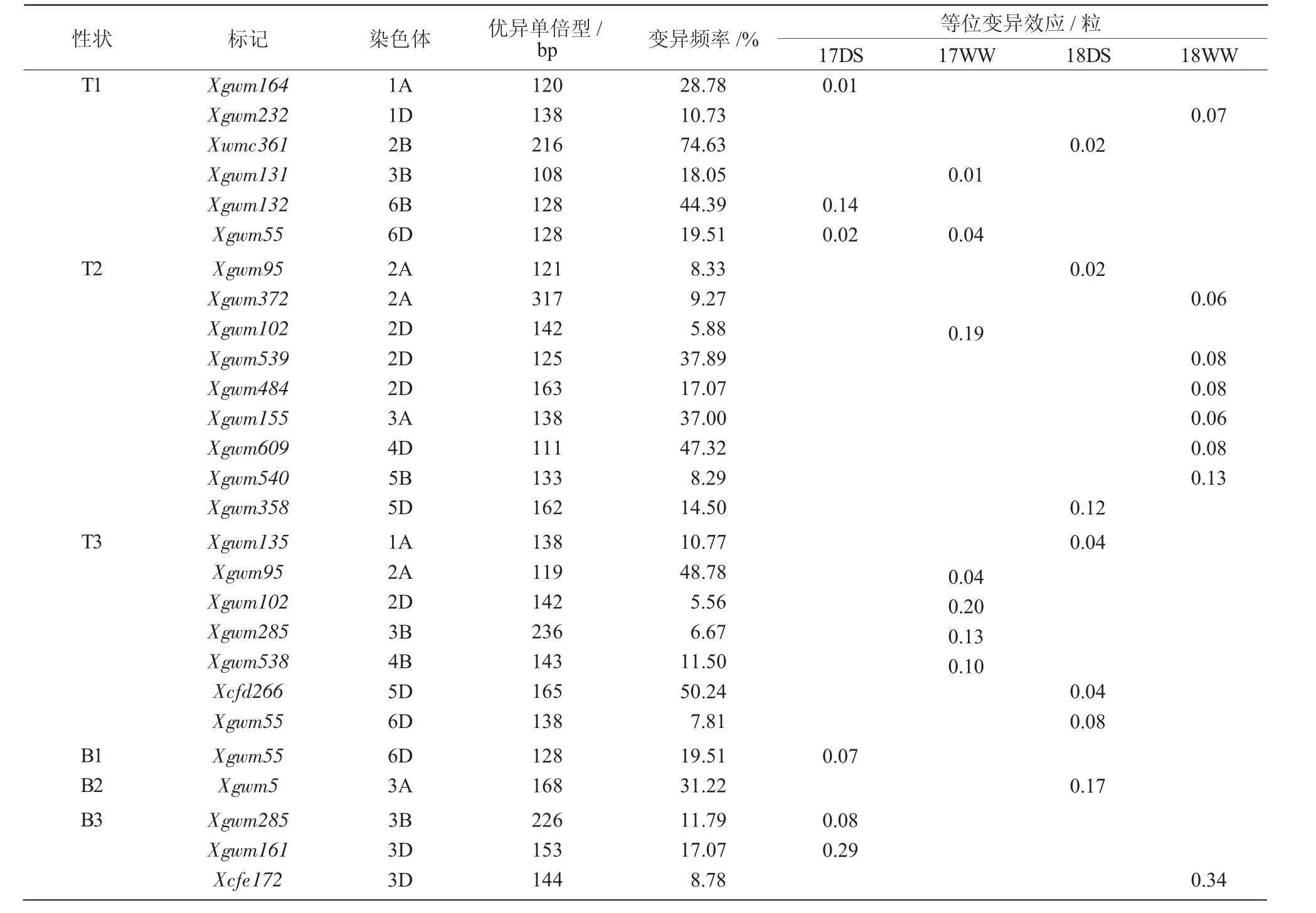
表3 关联标记的优异等位变异及其效应
3 讨论
3.1 多环境下关联的位点及其优异等位变异效应
本研究在2 a 共4 种环境下对小麦穗顶部和基部第1、2、3 小穗结实粒数进行关联分析,共关联到分布在1A、1D、2A、2B、2D、3A、3B、3D、4B、4D、5B、5D、6B 和6D 染色体上的27 对SSR 标记,共检测到28 个显著关联位点,表型解释率为5.73%~30.73%,且均检测到对小穗结实粒数具有正向效应的优异等位变异。在与穗顶部结实粒数相关的位点中,Xgwm132-6B128等位变异可以增加穗顶部第1 个小穗结实粒数0.14 粒;Xgwm102-2D142等位变异可以分别增加17WW 环境条件下的穗顶部第2、3 个小穗结实粒数0.19、0.20 粒;在与穗基部结实粒数相关的位点中,Xgwm5-3A168、Xgwm161-3D153和Xcfe172-3D144等位变异在不同环境条件下可以分别增加穗基部第1、2、3 小穗结实粒数0.17、0.29、0.34 粒;同时检测到Xgwm55-6D128等位变异可分别增加穗顶部和基部第1 小穗结实粒数0.04、0.07 粒。这些优异等位变异对于利用标记辅助选择,增加小穗结实粒数,进而增加穗粒数具有重要的应用价值。
3.2 关联位点的稳定性和一因多效性
本研究中发现,Xgwm55(6D)在17DS 和17WW条件下,同时与穗顶部第1 个小穗结实粒数显著关联,在17DS 和18DS 条件下分别与穗基部第1 个小穗结实粒数和穗顶部第3 个小穗结实粒数显著关联,而郭杰[22]研究发现,Xgwm55与穗基部第3 个小穗结实粒数显著关联;张东等[19]研究发现,Xgwm55在多个环境条件下与株高和穗长显著关联;PERRETANT 等[23]研究发现,Xgwm55与一个籽粒硬度相关的QTL 位点连锁。本研究在17WW 条件下,检测到标记Xgwm95(2A)与穗顶部第3 个小穗结实粒数显著关联,在18DS 条件下与穗顶部第2 个小穗结实粒数显著关联,且Xgwm95-2A119等位变异频率相对较高,为48.78%。魏添梅[24]在水、旱2 种环境下研究发现,Xgwm95与株高极显著关联;王晖等[25]检测到一个千粒质量QTL 位于2A 染色体的Xgwm372~Xgwm95标记区间;张颖君等[26]利用洋小麦/中优9507 建立的重组自交系群体,将与籽粒休眠相关的标记定位在Xgwm372~Xgwm95标记区间,与Xgwm95遗传距离为1.4 cM;张娜等[27]研究发现,标记Xgwm95与抗叶锈病基因Lr45连锁。本研究发现,在17WW 条件下,Xgwm102(2D)同时与穗顶部第2、3 个小穗结实粒数显著关联。张政等[28]研究发现,该位点与小麦开花期极显著关联。本研究发现,17WW 和17DS 条件下,Xgwm285(3B)分别与穗顶部和穗基部第3 个小穗结实粒数显著关联。常成等[29]研究发现,Xgwm285与除草剂抗药性显著相关。显然,以上检测到的这4 个位点具有稳定性和一因多效性,为今后分子标记辅助育种提供了重要参考价值。
4 结论
本研究利用4 个环境条件下的小麦穗顶部和基部结实粒数与106 对SSR 标记进行关联分析,共检测到28 个显著关联位点,发现Xgwm132-6B128、Xgwm102-2D142、Xgwm5-3A168、Xgwm161-3D153以及Xcfe172-3D144为增加小麦穗顶部或基部小穗结实粒数的优异等位变异位;发现Xgwm55(6D)、Xgwm95(2A)、Xgwm102(2D)和Xgwm285(3B)为多环境下稳定检测到具有一因多效性的标记位点,这些发现对于改善小麦穗顶部和基部结实粒数,增加籽粒产量具有重要参考价值。
