G蛋白偶联雌激素受体及其对免疫系统疾病的影响
2019-05-27王韫秀顾兆伟郝丽英
王韫秀,顾兆伟,郝丽英
(1.中国医科大学附属盛京医院 a.医疗保险工作部,b.耳鼻咽喉科,沈阳 110004; 2.中国医科大学药学院药物毒理教研室,沈阳 110122)
雌激素是一种类固醇激素,最主要的作用是促进女性的第二性征发育,建立并保持女性生殖功能,对子宫和乳腺的发育有重要的调控作用;雌激素还对大脑的发育至关重要,可减少卒中后神经元损伤,增加神经元连接,并在许多疾病甚至肿瘤中起作用。雌激素与细胞中并不活跃的雌激素受体结合后变得异常活跃,进而发挥其生物学效应。雌激素受体主要分为经典的核受体(包括雌激素受体α和雌激素受体β)和膜受体两大类[1]。随着对受体认识的深入,雌激素信号的复杂性也随之增加,核受体通过经典的雌激素受体通路,触发雌激素受体-雌激素复合物作为配体门控转录因子移位至细胞核,调节特异性靶基因的转录而发挥基因型调节效应;膜受体通过第二信使系统介导快速非基因效应。近年来,G蛋白偶联雌激素受体(G protein-coupled estrogen receptor,GPER)作为一种新型的雌激素膜受体成为国内外学者关注的热点。越来越多的研究发现,GPER是一种介导雌激素,是在免疫系统、神经系统、肾脏系统、血管系统和生殖系统发挥作用的重要受体[2-6]。现就GPER的发现、信号转导、配体及其对免疫系统疾病的影响予以综述。
1 GPER的发现
雌激素受体和盐皮质激素受体是类固醇激素核受体亚家族成员。有研究推测,雌激素的快速血管效应与第二信使系统有关,并发现了盐皮质激素醛固酮在平滑肌收缩中的快速效应[4,7]。有研究显示,活化盐皮质激素受体可以调节环腺苷酸的代谢、钙信号通路以及Src蛋白和促分裂原活化的蛋白激酶,以上信号机制与心脏收缩力、血管反应性、心肌纤维化、细胞增殖和凋亡的调节有重要关系[8]。在某种情况下,盐皮质激素和雌激素的快速效应与膜相关的经典类固醇受体有关(如雌激素受体和盐皮质激素受体)。但有研究推测,另有受体触发这些快速类固醇效应[9]。
有研究提示,G蛋白偶联受体30是通过非经典受体系统调节雌激素效应的主要物质。此前,G蛋白偶联受体30只是一种无已知内源性配体,且不被认识的孤立G蛋白偶联受体。随后将雌激素作为其主要配体,被命名为GPER。但有研究显示,醛固酮是GPER更有效的激动剂,将其命名为GPER可能属于误称,但尚未检测到醛固酮与GPER结合,故无证据支持GPER作为醛固酮受体行使作用[10-13]。
GPER可广泛表达,在生殖器官、胰腺、肝脏、心脏、血管、乳腺、肺脏、神经组织以及白细胞均有表达[14]。在新鲜离体鼠主动脉平滑肌细胞和内皮细胞均能检测到GPER的表达[8,10,15]。近年来对GPER作用分子机制的认识大多由其功能分析的推断得出。对GPER活化机制了解的欠缺由多种因素造成,主要与细胞膜上GPER的低表达以及GPER主要在细胞内表达与GPER选择性激动剂和拮抗剂的相对低效能结合和高水平的非特异性结合等有关[16]。
2 GPER的信号转导通路
雌激素受体α和雌激素受体β是参与雌激素基因效应的主要核受体。有证据表明,快速调节细胞信号通路可通过一种细胞膜上定位的雌激素受体介导,由此引发人们对GPER信号通路的研究[17]。GPER的定位主要在细胞内,与此前报道描述的细胞膜GPER内化相一致[18-19]。GPER活化介导的信号通路包括上皮生长因子受体继发反式活化通路、磷脂酰肌醇-4,5-二磷酸3-激酶通路和细胞外信号调节激酶通路,以上通路均与盐皮质激素受体和雌激素受体活化介导的非基因信号通路有关[20-21]。GPER信号通路通过表皮生长因子受体、上皮生长因子受体反式活化,同时涉及Src家族的非受体酪氨酸激酶,目前已被其他G蛋白偶联受体接受,GPER的刺激活化了金属蛋白酶并诱导肝素结合表皮生长因子的释放,进一步结合并活化上皮生长因子受体,导致下游信号分子(如细胞外信号调节激酶1和细胞外信号调节激酶2)的活化[22-23]。此外,17β-雌二醇介导的GPER活化触发了环腺苷酸的产生、细胞内钙动员以及磷脂酰肌醇-3-激酶活化[24]。对人乳腺癌细胞的深入研究显示,鞘氨醇激酶以及活化的整合素α5β对17β-雌二醇介导的上皮生长因子受体的反式活化起调节作用,雌激素刺激后,通过GPER调节整合素α5β在纤连蛋白集合过程中的作用[25]。此外,GPER还可通过活化环腺苷酸、细胞外信号调节激酶和磷脂酰肌醇-3-激酶的信号机制间接调节转录活性,GPER调控的基因包括编码c-Fos的FOS,并与其他蛋白质形成异二聚体和转录因子激活蛋白1;同时,这些信号通路也激活其他转录因子(如类固醇因子1),诱导其他基因的表达[26]。
GPER还可触发通过G蛋白偶联途径的信号通路,由于GPER对各种G蛋白亚型的选择性,与介导GPER调节作用的非G蛋白偶联通路相比,对G蛋白偶联途径的信号通路重要性的认识仍存在争议。基于对乳腺癌HEK293细胞和SkBr3细胞的相关研究,最初GPER的活化被认为是通过G蛋白与腺苷酸环化酶/蛋白激酶A的连接而活化[24],可检测到与Gαi和(或)Gαo活化一致的百日咳毒素敏感并依赖酪氨酸激酶的GPER介导的信号通路,但这种与G蛋白连接的推测并未得到认可。Filardo等[27]对MDA-MB-231乳腺癌细胞中GPER依赖的细胞外信号调节激酶活化的研究发现,含有细胞外信号调节激酶活化抑制蛋白依赖的信号通路可能导致G蛋白偶联受体激酶依赖的GPER效应。以上研究可作为G蛋白β和G蛋白γ亚单元依赖GPER介导细胞外信号调节激酶活化的证据,揭示了GPER活化中多种G蛋白依赖信号通路。
目前,由雌激素和醛固酮触发的GPER快速分子事件是研究的难点,但触发GPER所致的下游效应也具有重要的生理意义。GPER不仅参与介导雌激素的非基因作用,还参与醛固酮介导的非基因作用,GPER活化所致的基因影响被逐渐认识。基因作用范围较广,包括转录因子和辅助活化因子的调节,如c-Jun/c-Fos、TIF2、SF-1、YAP和TAZ。GPER的作用还包括对细胞周期调节蛋白的调节以及与程序化细胞死亡有关的蛋白调节;在某种情况下,还包括对经典的类固醇受体活性的调节作用。
3 GPER的配体
GPER和经典雌激素受体共享一个共同配体,导致两种受体类型对雌激素物质的相对选择性问题。雌激素作为配体,无构象灵活性,立体特异性识别位点有限,因此可以预测两种受体都能结合雌激素,结合类似的结构相关化合物谱,证实了大量与经典雌激素受体结合的化合物也能结合或激活GPER。GPER配体包括内源性激素(如雌激素、醛固酮以及各种合成的配体、药物、植物性雌激素)和环境激素[28]。
3.1雌激素 雌激素是促进雌性动物第二性征发育和器官成熟的重要物质,主要由卵巢和胎盘产生,少量由肝、肾上腺皮质和乳房分泌。孕期胎盘可大量分泌雌激素,男性睾丸也可少量分泌雌激素。雌激素受体分布在子宫、阴道、乳房、盆腔以及皮肤、膀胱、尿道、骨骼和大脑等组织,具有广泛而重要的生理作用,还对内分泌、心血管、代谢系统、骨骼的生长和成熟等有显著影响。天然雌激素主要包含17β-雌二醇、17α-雌二醇、雌酮、雌三醇及其二羟基和2-甲基雌二醇代谢产物[29]。雌激素通过与其受体的结合发挥生物效应。
3.2醛固酮 醛固酮通过各种在体和离体模型活化GPER依赖的功能通路,此效应并非醛固酮与GPER的直接结合,但可反映盐皮质激素受体与GPER的相互作用[30]。在盐皮质激素缺乏的受体模型中发现了GPER依赖的醛固酮效应[15]。在GPER表达而无雌激素受体表达的细胞模型中,未证实预测的醛固酮和雌激素均与GPER相互作用而导致醛固酮与雌激素竞争的结合,但与G蛋白偶联受体结合的基础条件(如饱和度、选择性、解离动力学以及类似于饱和实验的解离常数)并未得到证实[11-12]。醛固酮可通过GPER介导部分非盐皮质激素受体依赖效应,但GPER结合并激活醛固酮的分子机制仍待进一步研究证实。
3.3合成的配体 GPER特异性激动剂G-1以及拮抗剂配体G15和G36能有效地选择性结合GPER,触发GPER介导的生物学效应,主要用于区分GPER介导的雌激素效应和传统雌激素受体介导的雌激素效应。
GPER特异性激动剂和GPER特异性拮抗剂是目前GPER研究领域的重要配体,有效推进了对GPER生物意义的探究,使其在相关研究中的应用得到广泛认可,但对GPER活化或抑制的确切机制尚不清楚[7]。
3.4药物、植物雌激素和环境激素 除上述生理配体和合成配体外,还有很多化合物与GPER相互作用,主要包括选择性雌激素受体调节因子(如他莫昔芬)、雌激素受体拮抗剂(如ICI 182,780)和烟酸[31-32]。植物性雌激素(如染料木黄酮、白藜芦醇、橡黄素、绿茶儿茶素)以及环境激素或内分泌干扰物(如壬基酚和双酚A)也可能与GPER相互作用[33]。有研究证明,亚砷酸盐和镉可与GPER相互作用[34]。已有研究对以上化合物与GPER的相互作用进行回顾,但与GPER相互作用的重要性及其生物效应和治疗效应仍不清楚[29]。
4 GPER对免疫系统疾病的影响
雌激素治疗和抗雌激素治疗能够调节免疫系统的发育、成熟和功能[35-36]。有研究发现,GPER可在多种免疫细胞(T细胞、B细胞、单核巨噬细胞、中性粒细胞)中表达,可见GPER可调节雌激素在免疫系统中的某些特定作用[37-39]。
4.1GPER对胸腺的影响 在妊娠或长期雌激素暴露下,胸腺质量、细胞结构减少,并伴随CD4、CD8细胞亚群的改变[40]。前期雌激素受体α敲除鼠的研究提示,雌激素受体α只能部分调节雌激素对胸腺的作用,而雌激素受体β的作用更小[41]。GPER敲除鼠并未表现严重的免疫缺陷,但GPER在雌激素介导的胸腺萎缩中的作用明确。对雌激素受体α和GPER敲除鼠的直接比较中,雌激素介导的胸腺萎缩不仅在雌激素受体α敲除鼠中减弱,在GPER敲除鼠中也减弱。进一步分析提示,雌激素介导的胸腺萎缩包括发育阻断和凋亡[42]。雌激素受体α仅介导胸腺细胞发育过程中的早期发育阻断(尤其是CD4/CD8双阴选择胸腺细胞),GPER则在双阳选择胸腺细胞凋亡中发挥作用,并通过GPER特异性激动剂G-1得到进一步证实,即GPER特异性激动剂G-1可通过非雌激素受体α诱导的发育阻断而诱导胸腺凋亡[42]。有报道称,GPER的β半乳糖苷酶基因报告鼠的初始T细胞在无雌激素刺激时也可增加其凋亡率,G-1对雌激素诱导的胸腺萎缩无治疗作用,此外,GPER对雌激素诱导的胸腺萎缩无作用[43]。目前,导致以上结果的显著差异尚不明确。
4.2GPER对自身免疫性疾病的影响 雌激素作为抗炎治疗药也逐渐受到关注,尤其对多发性硬化的作用。妊娠期多发性硬化症状得到明显缓解提示雌激素的保护作用[44]。有关雌二醇对多发性硬化治疗的临床试验正在进行[45]。自身免疫性脑脊髓炎鼠实验模型的研究展现了雌激素的保护和治疗作用,通过临床评估发现,GPER基因敲除鼠的雌激素介导保护作用降低,而雌激素介导的脑白质损伤的保护作用完全缺失[46]。GPER的作用在G-1治疗脑脊髓炎的模型中得到进一步证实,在临床表现和组织学上均有保护作用,并与此前观察到的雌激素的保护作用相似[38,46]。G-1对GPER基因敲除鼠的作用被完全阻断,可见G-1的作用需要GPER的参与,且G-1对GPER具有选择性,但雌激素本身的保护作用仅部分阻断,提示雌激素发挥作用的机制包括雌激素受体α和GPER。但G-1单独激活GPER的野生鼠可完全复制雌激素的效应,可见G-1在GPER活化时发挥保护作用,而不需雌激素受体α的活化[46]。雌激素受体α基因敲除鼠中G-1的效应尚未得到证实,G-1介导的保护作用是否需要雌激素受体α的表达或基础活化仍需进一步研究确认。有研究报道,GPER可通过上调程序化死亡来增强调节T细胞的抑制活化和抑制巨噬细胞炎症介质产生,从而发挥保护作用[38,46]。除雌激素本身的保护作用外,GPER基因敲除鼠中,炔雌醇对脑脊髓炎无治疗作用;但在雌激素受体α基因敲除鼠中,炔雌醇对脑脊髓炎仍有治疗作用,这与白细胞介素(interleukin,IL)-10的产生增加有关[47]。GPER基因敲除脑脊髓炎鼠的雌激素依赖的维生素D3的保护作用消失,与降低的抗炎因子有关,再次证实脑脊髓炎模型中GPER是维生素D3介导雌激素依赖的保护效应中的关键因素[48]。与GPER活化的脑脊髓炎的抗炎作用一致,大龄GPER基因敲除鼠可能继发于其他代谢功能障碍而表现出促炎状态[49]。此外,Brunsing等[50]研究也证实,在体外促炎反应的Th17细胞极化条件下,G-1在自身免疫性疾病中的免疫抑制作用能够刺激Foxp3表达(调节性T细胞的标志)和IL-10的产生,并在体内G-1治疗时得到验证。总之,GPER活化具有抗炎效应。
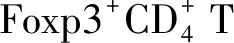
有研究评估了哮喘中GPER对嗜酸粒细胞功能的影响发现,人外周血嗜酸粒细胞中存在GPER信使RNA和蛋白表达;GPER特异性受体激动剂G-1预培养可增强嗜酸粒细胞趋化因子对人嗜酸粒细胞的定向趋化作用;抑制嗜酸粒细胞自发凋亡和胱天蛋白酶-3的活化,且不受环腺苷酸磷酸二酯酶抑制剂或磷脂酰肌醇-3-激酶抑制剂的影响;嗜酸粒细胞与IL-5共培养较静止期嗜酸粒细胞,可增加G-1诱导凋亡和胱天蛋白酶3的活化,但并未观察到G-1对嗜酸粒细胞的脱颗粒作用[52]。另有研究显示,雌激素可通过GPER/环腺苷酸/蛋白激酶A/环磷腺苷效应元件结合蛋白信号通路调节脊椎动物嗜酸粒细胞功能,体外GPER活化能轻微减少嗜酸粒细胞的呼吸爆发,并显著改变编码主要促炎和抗炎介质的基因表达谱[39]。
4.4GPER对造血系统的影响 GPER可在前体B细胞急性淋巴细胞白血病中通过CC趋化因子配体18[chemokine (C-C motif) ligand 18,CCL18]减弱CXC家族趋化因子受体4反应[53]。CCL18和CXC趋化因子配体12[chemokine (C-X-C motif) ligand 12,CXCL12]是血清中具有较高水平的稳态同源趋化因子。多种疾病可出现CCL18的升高(如儿童急性淋巴细胞白血病),但功能尚不清楚。CXCL12是造血过程和B细胞回巢过程中的关键趋化因子。研究显示,CCL18干扰了CXCL12介导的前体B细胞急性淋巴细胞白血病细胞活化[53]。CCL18显著降低前体B细胞急性淋巴细胞白血病细胞株中CXCL12诱导的钙动员、趋化性和细胞增殖,在前体B细胞急性淋巴细胞白血病患者的原始细胞中也观察到此结果,但在普通急性淋巴细胞白血病患者中并没有发现;CCL18对通过GPER调节CXCL12诱导的反应,CCL18与表达GPER的细胞结合后,GPER特异性拮抗剂可阻断这种结合并消除由CCL18介导的功能效应。CCL18可以通过干扰雌二醇或其他化学激动剂等配体活化GPER。由此可见,CCL18经由与GPER的相互作用在前体B细胞急性淋巴细胞白血病细胞CXC家族趋化因子受体4依赖反应中起重要的调节因子作用[53]。
雌激素对双向神经内分泌免疫的相互作用起重要作用,通过雌激素受体α、雌激素受体β和(或)GPER发挥作用。有研究发现,雌激素受体α、雌激素受体β和GPER可在鲤鱼淋巴组织以及某些肾脏单核巨噬细胞、中性粒细胞和淋巴细胞表达,雌激素受体β也在一些头肾淋巴细胞中表达,但在原始外周血淋巴细胞中无表达;免疫刺激可改变细胞类型和受体表达,取决于活化和成熟两个阶段,提示白细胞对雌激素刺激的直接反应,并由此确定体外雌激素对单核巨噬细胞活性氧类产生的作用;在佛波酯刺激细胞中,短期雌激素培育可增加活性氧类的产生,结果与GPER调节一致,部分通过磷脂酰肌醇-3-激酶活化发挥作用,进一步证明了雌激素受体介导的神经内分泌免疫沟通是一种进化保护[54]。
5 小 结
GPER作为新发现的雌激素膜相关受体,在各领域的作用越来越受到关注。目前,GPER对免疫系统的影响和重要作用也逐渐引起关注,如GPER对胸腺等重要免疫器官的影响,对多发性硬化等自身免疫性疾病的影响,对哮喘等过敏性疾病的影响以及通过与CCL18的相互作用对前体B细胞等造血细胞发育的影响等。但以上研究还局限于体外实验和动物实验阶段,仍需更多深入的基础性研究其在免疫系统疾病中的作用,以便对免疫系统疾病发病机制的深入了解,进而指导相关治疗的研究,期待GPER的相关研究为免疫相关疾病甚至肿瘤免疫的研究提供新思路。
