LncRNA HOTAIR在肿瘤中转录调控及作用机制的研究进展
2019-05-27马旭兰代荫梅
马旭兰,代荫梅
(首都医科大学附属北京妇产医院妇科,北京 100026)
在人类基因组中,稳定转录的蛋白质编码基因比例仅占2%,高达90%的真核基因组中DNA转录产物均为非编码RNA[1]。非编码RNA包括短链非编码RNA(长度为20~50个核苷酸,如微RNA、小干扰RNA等),中等链非编码RNA(长度为50~200个核苷酸,如核小RNA、核仁小RNA等)和长链非编码RNA(long non-coding RNAs,lncRNAs)[长度为200~100 000个核苷酸,如同源异型盒基因转录反义基因间RNA(HOX transeript antisense intergenitic RNA,HOTAIR)、肺腺癌转移相关转录本1等][2]。其中,lncRNAs可在不同层面广泛参与基因组的调控过程,如表观遗传学、转录水平及转录后水平等[3]。HOTAIR是第一个被发现的反义转录lncRNAs,长为2.2 kb,定位于第12号染色体的HOXD基因座,由Rinn等[4]在人成纤维细胞中发现,是目前探究的热点lncRNAs之一。有研究证实,HOTAIR在乳腺癌、子宫内膜癌、结直肠癌、宫颈癌等多种肿瘤中高表达,并均可促进肿瘤的增殖、侵袭、转移、复发和耐药等恶性生物学行为的进展[5-8]。现对HOTAIR在肿瘤中转录调控及作用机制的研究进展予以综述。
1 HOTAIR的转录调控机制
HOTAIR的转录调控机制异常复杂,受许多因素调控,但目前各因素在不同肿瘤中的研究较少,有待进一步研究。Bhan等[5]研究发现,HOTAIR转录受雌激素正向调控,应用17β-雌二醇刺激雌激素受体阳性的人乳腺癌MCF7细胞后发现,HOTAIR被诱导转录,且在暴露于雌激素受体抑制剂他莫昔芬后转录水平受到抑制。研究表明,HOTAIR转录由核受体共抑制因子维持在基础状态,暴露于雌激素时,雌激素通过雌激素-雌激素受体-雌激素受体反应元件途径与多种雌激素受体共调节因子共同结合在HOTAIR启动子上,解除核受体共抑制因子对HOTAIR的转录抑制,诱导HOTAIR转录[5,9]。此外,在子宫内膜癌[10]、前列腺癌[11]等肿瘤中,雌激素诱导HOTAIR转录水平上调也得到了证实。Bhan等[12]进一步实验发现,雌激素内分泌干扰物双酚A和己烯雌酚也可发挥类雌激素作用,诱导HOTAIR转录水平上调。
原癌基因c-Myc也是影响HOTAIR转录的因素。Ma等[13]对胆囊癌组织和细胞的研究发现,HOTAIR是c-Myc的直接作用靶点,c-Myc正向调控HOTAIR的表达。具体作用机制为c-Myc通过识别并作用于HOTAIR上游区域的假定E-box元素(c-Myc靶标反应元件),导致HOTAIR表达上调,敲减c-Myc可降低HOTAIR的表达及其启动子活性,上调c-Myc则可增加HOTAIR表达及其启动子活性。此外,Li等[14]对乳腺癌的研究证实,HOTAIR是c-Myc激活其靶基因转录的关键效应因子。转化生长因子-β1也可刺激HOTAIR转录上调。Ren等[15]研究发现,癌相关成纤维细胞通过分泌转化生长因子-β1激活HOTAIR转录,促进乳腺癌细胞的转移,表明转化生长因子-β1表达增加可显著增强HOTAIR表达,而转化生长因子-β1抑制剂减弱HOTAI活化,由此可见,转化生长因子-β1/HOTAIR轴有望成为乳腺癌治疗的靶标。
Yang等[16]对多种癌细胞系进行实验研究发现,重组人骨桥蛋白可以时间和剂量依赖方式诱导HOTAIR转录,而骨桥蛋白可以下调重组人干扰素调节因子1的表达,并通过染色质免疫沉淀和荧光素酶活性测定实验发现,干扰素调节因子1可以与HOTAIR启动子区域结合并降低其转录活性,且细胞过度表达的干扰素调节因子1可下调HOTAIR的表达。可见,骨桥蛋白作为细胞外基质蛋白可以通过减弱干扰素调节因子1的抑制作用来刺激HOTAIR转录,从而促进癌细胞的侵袭和转移。此外,信号转导及转录激活因子3信号通路也可调控HOTAIR的转录。Sun等[17]对头颈部鳞状细胞癌细胞的体内外实验研究发现,HOTAIR和组蛋白甲基转移酶zeste基因增强子同源物2(enhancer of zeste homolog 2,EZH2)是头颈部鳞状细胞癌中信号转导及转录激活因子3信号转导的下游效应分子,信号转导及转录激活因子3可通过与HOTAIR编码基因的启动子结合上调HOTAIR转录,从而增强EZH2介导的表观遗传沉默,表明信号转导及转录激活因子3/HOTAIR信号轴以依赖EZH2的方式调控头颈部鳞状细胞癌生长,为靶向信号转导及转录激活因子3/HOTAIR/EZH2信号转导治疗头颈部鳞状细胞癌提供了理论基础。
2 HOTAIR的作用机制
2.1HOTAIR通过与多梳蛋白抑制复合体2(polycomb repressive complex 2,PRC2)和赖氨酸特异性组蛋白去甲基化酶1结合发挥作用 LncRNA HOTAIR最经典的作用机制是发挥lncRNAs支架分子的功能,招募并结合PRC2和赖氨酸特异性组蛋白去甲基化酶1复合体,形成组蛋白修饰复合物,定位至特定的HOX基因位点,使该位点发生表观遗传学沉默,促进肿瘤恶性生物学行为的进展。其中,HOTAIR的3′端结构域招募赖氨酸特异性组蛋白去甲基化酶1复合体,发挥去除染色体组蛋白H3第4位赖氨酸二甲基化状态(即去甲基化)的作用,通过调控靶基因表达发挥促癌作用。而PRC2复合体与其5′端结构域结合,具有组蛋白甲基化转移酶活性,其核心成分主要由EZH2、EED、SUZ12亚基构成,通过介导染色体组蛋白H3第27位赖氨酸三甲基化导致一系列细胞生长所必需基因的沉默[18-19]。Betancur和Tomari[20]对细胞中纯化出的PRC2的各个组分的研究发现,EED负责调节EZH2、SUZ12的RNA结合活性,是PRC2的伴侣蛋白,而EZH2和SUZ12可以直接与RNA结合,其中EZH2是发挥主要作用的亚基。EZH2的失调与多种肿瘤的表观遗传畸变都存在联系。多项研究表明,HOTAIR通过与EZH2结合在多种肿瘤中发挥促癌作用[21-24]。Gonzalez等[25]的研究证明,在转录激活因子中,EZH2还具有PRC2非依赖性功能,也可单独发挥作用。
2.2HOTAIR作为竞争性内源RNA发挥作用 竞争性内源RNA是在转录后水平相互调控的一类RNA的总称,具有相同的微RNA(microRNA,miRNA)应答元件,可通过该应答元件竞争同一种miRNA,从而实现自身间的相互作用及参与靶基因的表达调控[26]。多项研究表明,lncRNA HOTAIR可作为竞争性内源RNA,通过“海绵效应”招募并结合miRNA抑制其表达,从而促进肿瘤的发生与发展(表1)[10,23,27-35]。以上发现有助于对肿瘤发生与发展机制的理解,并为治疗该肿瘤提供新的研究方向和治疗靶点。
2.3HOTAIR通过激活信号通路发挥作用 HOTAIR可通过调节Wnt/β联蛋白信号通路的活性来调控肿瘤的进展。Guo等[36]纳入北京协和医院病理科标本库中48例接受顺铂作为新辅助化疗的非小细胞肺癌患者的组织进行研究发现,HOTAIR通过调节Wnt信号通路对非小细胞肺癌顺铂耐药发挥重要作用,可能是影响非小细胞肺癌患者临床预后的重要因素。Cheng等[37]经过一系列实验发现,敲减HOTAIR可上调miR-34a抑制磷脂酰肌醇-3-激酶/蛋白激酶B和Wnt/β联蛋白信号通路,从而抑制胃癌细胞对顺铂的耐药性。Xiao等[30]的研究发现,HOTAIR可通过调节miR-203a-3p的表达水平及Wnt/β联蛋白信号通路的活性调节结直肠癌的进展和化学耐药性。
此外,HOTAIR还通过其他信号通路发挥作用。Yu等[38]研究表明,HOTAIR可通过调节p53/蛋白激酶B/c-Jun氨基端激酶信号通路调节乳腺癌细胞的增殖、凋亡、迁移和侵袭。Liu等[39]研究显示,慢病毒介导的HOTAIR沉默通过激活凋亡相关基因Bax/胱天蛋白酶3抑制肺腺癌中的转化生长因子-α/表皮生长因子受体信号转导通路,从而恢复肿瘤细胞对吉非替尼的敏感性。
2.4HOTAIR通过上皮-间质转化途径发挥作用 HOTAIR可通过参与上皮-间质转化途径调控肿瘤的发生。Dasgupta等[40]研究发现,miR-203可抑制HOTAIR表达,并通过上皮-间质转化途径调节肾细胞癌的发生。Lu等[41]研究显示,在口腔鳞状细胞癌中,HOTAIR过度表达增强了癌细胞的转移潜能和上皮-间质转化特征,还发现HOTAIR的表达与间充质标记呈正相关,与临床样本中的上皮标记呈负相关。此外,Kaplan-Meier生存分析表明,高水平的HOTAIR是口腔鳞状细胞癌患者生存率低的强预测因子。综上所述,HOTAIR介导的癌症干性(具有能够自我更新和分化的干细胞特性)和转移与上皮-间质转化的调节相关。
转录因子Snail是细胞特性和上皮-间质转化的主要调节因子,直接抑制广泛的上皮基因库。Battistelli等[24]研究显示,HOTAIR介导Snail和EZH2的增强子之间的相互作用,将EZH2募集到特定的基因组位点。Snail活性抑制取决于Snail/HOTAIR/EZH2HOTAIR:同源异型盒基因转录反义基因间RNA;miRNA:微RNA;NPM1:核仁磷酸蛋白1;notch3:notch3信号通路;Ki67:细胞核增殖抗原;PCNA:增殖细胞核抗原;Wnt/β-catenin:Wnt/β联蛋白;BCL2:B淋巴细胞瘤2基因;NRP2:神经纤毛蛋白质2;GPC5:磷脂酰肌醇蛋白聚糖家族成员5;PTPN14:蛋白酪氨酸磷酸酶14;GLS:谷氨酰胺酶三联复合物的形成。以上结果证明,HOTAIR在Snail介导的上皮-间质转化中起关键作用。
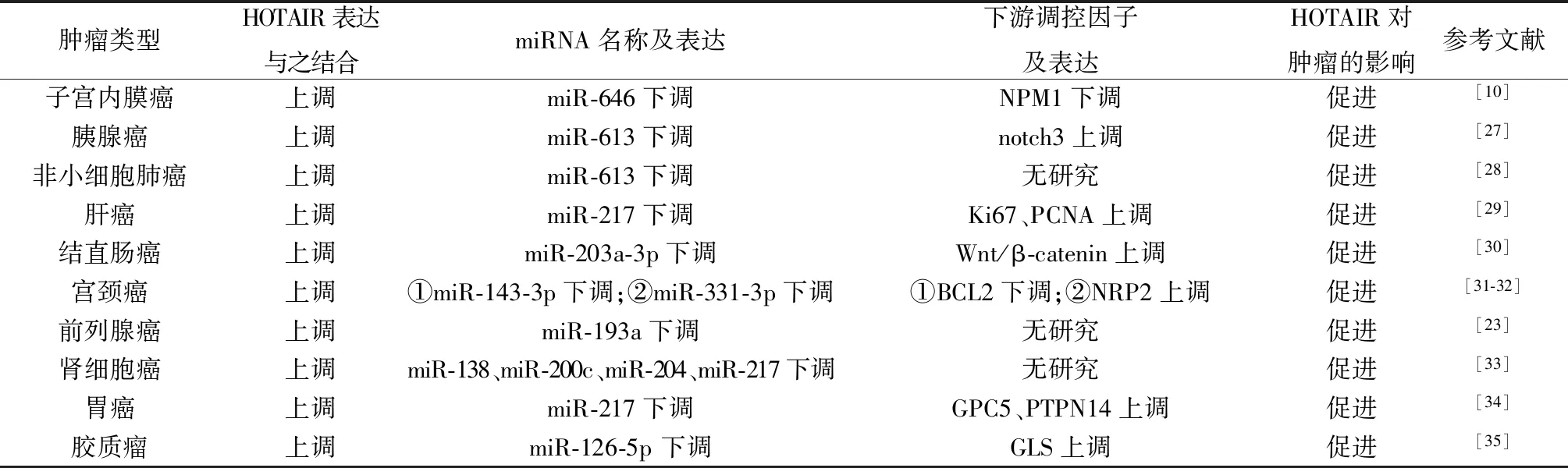
表1 HOTAIR在肿瘤中作为竞争性内源RNA发挥作用的机制
2.5HOTAIR促进蛋白质泛素化 HOTAIR还可通过促进蛋白质泛素化来调控肿瘤的发展。Yoon等[42]对人HeLa细胞、人成纤维细胞IDH4和WI-38以及小鼠胚胎成纤维细胞的实验研究显示,HOTAIR可作为蛋白质泛素化的平台,帮助组装E3泛素连接酶与其各自底物相结合,促进该复合物泛素化,并加速其降解。Mex3b和Dzip3为E3泛素连接酶,具有与HOTAIR结合的特定RNA结合域。其中,Mex3b存在于细胞核和细胞质中,相应的泛素化底物为Snurportin-1蛋白。而Dzip3只存在于细胞质的囊泡中,可促进Ataxin-1蛋白的泛素化。Xue等[43]应用RNA-蛋白质相互作用预测软件筛选可能与Runt相关转录因子3(Runt-related transcription factor 3,Runx3)(一种在胃癌中发挥抑癌作用的重要转录因子)结合的lncRNAs发现,HOTAIR与Runx3蛋白结合并鉴定HOTAIR的相关片段后,通过RNA免疫沉淀和RNA敲减实验验证了HOTAIR与Runx3之间的相互作用,通过免疫共沉淀评估E3泛素连接酶Mex3b和Dzip3在HOTAIR诱导的Runx3泛素化中的作用,利用Pearson相关性分析HOTAIR信使RNA表达与Runx3蛋白表达之间的相关性,结果发现,Mex3b可作为E3泛素连接酶参与HOTAIR诱导的Runx3普遍降解过程,沉默HOTAIR或Mex3b的表达减弱Runx3的降解。由此可见,HOTAIR与Mex3b相互作用可诱导Runx3蛋白的泛素化,增强胃癌细胞的侵袭能力,并为胃癌的转移提供潜在的治疗靶点。目前,关于HOTAIR促进蛋白质泛素化的相关研究较少,HOTAIR在不同肿瘤中参与不同蛋白质的泛素化过程仍存在很大的研究潜力和意义,有待进一步研究。
2.6HOTAIR调控自噬的发生 自噬也称为细胞的“自我消化”,是一种高度保守的生物学行为,有助于维持细胞新陈代谢的稳定,不仅在正常细胞中发挥作用,还在肿瘤细胞适应缺氧、耐受外界刺激和诱导肿瘤化疗耐药性中发挥重要作用,已成为近年来研究的焦点[44]。Wang等[44]利用小干扰RNA干扰口腔鳞状细胞癌细胞中HOTAIR的表达,并通过透射电镜、Western blotting法及流式细胞术检测细胞的自噬及凋亡,评估HOTAIR对中位致死剂量顺铂敏感性影响的研究表明,HOTAIR在口腔鳞状细胞癌细胞中作为致癌基因可加速细胞自噬作用,减少细胞凋亡,并增加口腔鳞状细胞癌对顺铂的耐药性。Yu等[45]研究发现,卵巢癌细胞中沉默HOTAIR可抑制自噬相关基因7和微管相关蛋白1轻链3Ⅱ/Ⅰ的表达,从而抑制自噬过程,而自噬相关基因7和微管相关蛋白1轻链3Ⅱ/Ⅰ的表达水平可随顺铂浓度的增加而增加,下调自噬相关基因7可抑制顺铂诱导的自噬,随后下调HOTAIR表达联合顺铂刺激后发现,卵巢癌细胞增殖和自噬水平受到抑制,抗凋亡蛋白Bcl-2的表达下降,而促凋亡蛋白Bax表达升高,细胞凋亡增加,可见下调HOTAIR的表达可通过抑制顺铂诱导的自噬提高顺铂对卵巢癌的敏感性,从而提高顺铂治疗卵巢癌的效果。此外,HOTAIR可促进自噬的发生,导致非小细胞肺癌[46]、子宫内膜癌[47]、乳腺癌[48]、软骨肉瘤[49]、肝细胞癌[50]等肿瘤细胞对化疗药物耐药性增加。
3 小 结
近年来,有关lncRNA HOTAIR的研究不仅局限于HOTAIR对多种肿瘤组织和细胞的表达水平以及对肿瘤细胞增殖、凋亡、侵袭和迁移的影响,还深入研究了HOTAIR在不同肿瘤中的具体转录调控及作用机制等,取得了显著的成果。但还有待对整个HOTAIR复杂的调控机制进行深入研究,尤其是一些相对冷门的肿瘤中HOTAIR作用机制的研究。因此,进一步阐明HOTAIR在不同肿瘤中的具体作用机制是实现肿瘤精准治疗的理论基础。
目前,尚未将HOTAIR具体作用机制的研究成果应用于临床。开发通过血液、尿液或其他体液等检测HOTAIR表达水平的试剂盒等,可实现多种肿瘤的早发现、早诊断、早治疗以及准确评估预后等,还可将能够敲低HOTAIR或影响其发挥作用的上下游分子制成靶向药物应用于临床,下调HOTAIR水平,从而治疗肿瘤和解决化疗耐药性等。随着HOTAIR的相关研究思路、方法、技术等的不断更新与完善,积极开发HOTAIR的临床应用潜力,将使其成为判定恶性肿瘤发生和预后的生物标志物以及治疗恶性肿瘤的新靶点。
