丹参酮ⅡA注射液对缺血缺氧性脑损伤新生大鼠皮质神经元自噬及Akt-mTOR通路的影响*
2019-02-26苏仁意许细平孙乾朕
朱 波 杨 艳 苏仁意 许细平 孙乾朕
(湖北医药学院附属襄阳市第一人民医院,湖北 襄阳 441000)
缺氧缺血性脑损伤(HIBD)是造成新生儿多种神经功能障碍的疾病之一,严重威胁新生儿生命健康[1]。据调查,围生期HIBD新生儿死亡率高达20%,经治疗幸存患儿中约40%存在脑神经损害等后遗症[2]。目前,HIBD主要依靠亚低温疗法进行治疗,但其预后效果并不理想。相关研究指出,病理性神经元自噬是造成多种神经元损伤凋亡的重要原因之一[3]。因此,寻找调节神经元自噬药物的研究是目前治疗HIBD的新思路。丹参酮ⅡA是丹参主要有效成分之一,难溶于水,易溶于有机溶剂。丹参酮ⅡA因含有醌型结构,具有多种药理药效,例如改善冠状动脉循环、修复心肌损伤、抗细胞凋亡、神经保护等[4]。相关研究显示,丹参酮ⅡA对多种脏器的缺氧缺血性损伤具有保护作用[5-6]。任陈等研究发现丹参酮ⅡA能明显减轻放射线在体外对海马神经元细胞的放射损伤作用,并推测其保护机制可能与调节放疗过程中神经元自噬相关[7]。因此,本研究拟通过建造新生大鼠缺血缺氧性脑损伤模型,观察丹参酮ⅡA对HIBD新生大鼠皮质神经元自噬及Akt-mTOR通路影响并探讨其机制。现报告如下。
1 材料与方法
1.1 实验动物 清洁级7日龄SD(Sprague-Dawley)大鼠80,雌雄不计,体质量11.6~12.17 g,平均体质量(11.98±0.32)g,由河南省实验动物中心提供,许可证号:SYXK(豫)2015-0003。
1.2 试剂与仪器 丹参酮ⅡA磺酸钠购自上海上药第一生化药业有限公司,国药准字H31022558,批号160916。蛋白酶抑制剂购自上海子起生物科技有限公司,货号E429-10MG。RIPA裂解液购自北京君诺德生物技术有限公司,货号7011A。BCA蛋白浓度检测试剂盒购自上海经科化学科技有限公司,货号JK-201。LC3-Ⅰ、LC3-Ⅱ、Beclin-1、p-Akt、p-mTOR、p-S6、GADPH蛋白抗体及HRP缀合的二抗均购自药明康德。透射电子显微镜购自尼康仪器(上海)有限公司。酶标仪购自美国伯腾仪器。
1.3 分组与造模 将SD幼鼠随机分为5组,每组16只,分别为假手术组、模型组、高剂量组(0.1 mg/g)、中剂量组(0.05 mg/g)、低剂量组(0.02 mg/g)。假手术组仅左颈部切口处理,其余各组进行造模。3%乙醚麻醉幼鼠,手术暴露左侧颈主动脉,分离神经,结扎血管,缝合伤口。放回原饲养环境,恢复3 h后,将幼鼠置于常压缺氧舱内,温度37℃,湿度45%~55%,持续通入8%O2和92%N2缺氧处理2.5 h,幼鼠出现左旋即为造模成功。
1.4 给药方法 高剂量组、中剂量组、低剂量组幼鼠分别给予不同剂量的丹参酮ⅡA注射液,于出现左旋现象时腹腔注射1次,24 h后重复注射1次。假手术组与模型组分别按照上述给药方式给予等体积0.9%氯化钠注射液。
1.5 标本采集与检测 1)大脑皮质超微结构观察。缺氧缺血处理32 h后,取幼鼠断头取左脑,利用解剖镜迅速剥离大脑皮质,置于冰上盛有2.5%戊二醛容器中进行处理,用1%锇酸对脑组织进行固定,乙醇和丙酮不同浓度由低至高进行脱水,环氧树脂包埋,切片,利用乙酸双氧铀和硝酸铅染色,置于透射电子显微镜下观察拍片。2)TTC染色观察脑梗死面积。缺氧缺血处理36 h,取幼鼠断头,取左脑置于冰上。Rat Brain Matrix由前之后切片,厚度5 μm。室温条件下,将切片置于2%TTC中,孵育35 min,PBS缓冲液洗涤,10%中性甲醛固定。数码相机采集图片,正常脑组织为红色,梗死组织为白色。利用Image J对采集图片急性分析,统计梗死面积百分比。3)蛋白免疫印迹法检测脑组织自噬蛋白表达。缺氧缺血处理48 h,取幼鼠进行断头取脑,取左侧脑组织,例用含有蛋白酶抑制剂的RIPA裂解液提取脑组织蛋白。4℃下10 000×g离心8 min后取上清,BCA蛋白浓度检测试剂盒检测总蛋白,10%SDS-PAGE分离胶分离蛋白,半干法转移蛋白至PVDF膜上。PBS配制的5%脱脂乳对膜进行封闭,4℃过夜。弃封闭液,PBS缓冲液清洗膜5次,每次4 min。LC3-Ⅰ、LC3-Ⅱ、Beclin-1、p-Akt、p-mTOR、p-S6 蛋白抗体及内参GADPH蛋白抗体作为一抗,室温孵育3 h。弃溶液,PBS缓冲液洗涤5次,每次4 min。室温条件下,HRP缀合的二抗孵育1.5 h。利用Image J软件分析条带灰度,评估蛋白相对表达量。
1.6 统计学处理 应用SPSS20.0统计软件。计量资料以(x±s)表示,两组间比较行t检验。P<0.05为差异具有统计学意义。
2 结 果
2.1 各组新生大鼠大脑皮质超微结构 见图1,表1。假手术组神经元结构正常,细胞核、线粒体、粗面内质网、高尔基体及溶酶体均正常存在。模型组中观察到皮质神经元中存在大量的自噬体和自噬溶酶体,其泡状自噬体中包裹有可见的细胞质结构。相较于模型组,高剂量组、中剂量组、低剂量组中自噬体数量明显减少,且呈一定的剂量依赖性(P<0.05)。

图1 各组新生大鼠大脑皮质超微结构(乙酸双氧铀和硝酸铅染色,5000倍)

表1 各组新生大鼠大脑皮质超微结构(x±s)

图2 各组新生大鼠脑梗死组织切片
2.2 各组新生大鼠脑梗死比较 与假手术组比较,模型组中幼鼠大脑皮质、海马等区域呈现明显的大面积白色梗死。与模型组比较,高剂量组、中剂量组、低剂量组中幼鼠脑梗死程度均有所改善。见图2。对各组中幼鼠脑梗死面积进行定量,发现与假手术组比较,模型组中幼鼠脑梗死面积显著增加 (P<0.05)。与模型组比较,高剂量组、中剂量组、低剂量组脑梗死面积显著减小,且呈一定的剂量依赖性(P<0.05)。见表2。
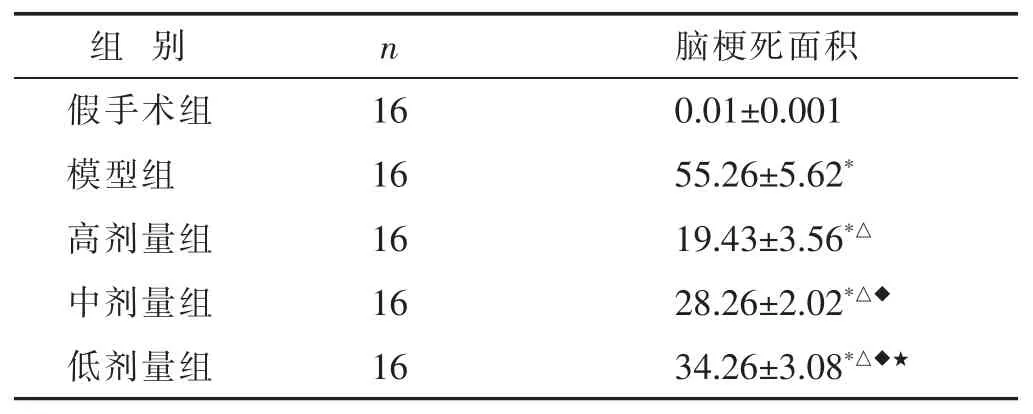
表2 各组新生大鼠脑梗死情况比较(%,x±s)
2.3 各组新生大鼠脑组织自噬蛋白表达情况比较
见图3,表3。与假手术组比较,模型组新生大鼠脑组织中 LC3-Ⅰ/(LC3-Ⅰ+LC3-Ⅱ)的比值显著增高,差异具有统计学意义(P<0.05)。与模型组比较,高剂量组、中剂量组、低剂量组新生大鼠脑组织中LC3-Ⅰ/(LC3-Ⅰ+LC3-Ⅱ)的比值显著降低,呈剂量依赖性(P<0.05)。模型组新生大鼠脑组织中Beclin-1蛋白表达水平与假手术组比较显著上升(P<0.05)。与模型组比较,高剂量组、中剂量组、低剂量组新生大鼠脑组织中Beclin-1蛋白表达水平显著下降,且呈剂量依赖性(P<0.05)。
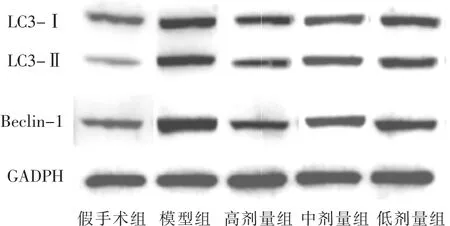
图3 各组新生大鼠脑组织自噬蛋白表达情况

表3 各组新生大鼠脑组织自噬蛋白表达情况比较(x±s)
2.4 各组新生大鼠脑组织中p-Akt表达情况比较
见图4,表4。与假手术组比较,模型组新生大鼠脑组织中p-Akt蛋白的表达水平显著降低(P<0.05)。与模型组比较,高剂量组、中剂量组、低剂量组新生大鼠脑组织中p-Akt蛋白的表达水平显著升高,呈剂量依赖性(P<0.05)。

图4 各组新生大鼠脑组织中p-Akt表达情况

表4 各组新生大鼠脑组织中p-Akt表达情况比较(x±s)
2.5 各组新生大鼠脑组织中p-mTOR表达情况比较见图5,表5。与假手术组比较,模型组新生大鼠脑组织中p-mTOR蛋白的表达水平显著降低(P<0.05)。相较于模型组,高剂量组、中剂量组、低剂量组新生大鼠脑组织中p-mTOR蛋白的表达水平呈显著升高,且呈剂量依赖性(P<0.05)。

图5 各组新生大鼠脑组织中p-mTOR表达情况

表5 各组新生大鼠脑组织中p-mTOR表达情况比较(x±s)
2.6 各组新生大鼠脑组织中p-S6表达情况比较 见图6,表6。相较于假手术组,模型组新生大鼠脑组织中p-S6蛋白的表达水平显著降低(P<0.05)。相较于模型组,高剂量组、中剂量组、低剂量组新生大鼠脑组织中p-S6蛋白的表达水平呈显著升高,且呈剂量依赖性(P<0.05)。

图6 各组新生大鼠脑组织中p-S6表达情况

表6 各组新生大鼠脑组织中p-S6表达情况比较(x±s)
3 讨 论
HIBD的发生机制极其复杂,与氧自由基、神经元自噬、神经元凋亡等密切相关[9]。相关研究指出,大脑发生缺氧缺血早期便可引起细胞凋亡及神经元病理性自噬等[10]。本研究实验显示,与假手术组比较,模型组中幼鼠大脑皮层、海马等区域呈现明显的大面积白色梗死,表明造模成功;高剂量组、中剂量组、低剂量组中脑梗死面积较模型组显著减小,且呈一定的剂量依赖性,表明丹参酮ⅡA能有效改善脑梗死情况。
自噬是降解细胞内蛋白和损伤细胞器,稳定细胞环境的主要方式[11]。相关研究指出,当自噬处于稳态平衡时对机体具有保护作用,但当自噬平衡状态打破时,将导致机体出现损伤[12]。 Lu 等[13]研究中发现,新生大鼠缺血缺氧性脑损伤模型中自噬体数量明显增加,自噬标志蛋白表达水平明显上升,当自噬抑制剂作用于模型可显著降低自噬体数量,抑制神经细胞凋亡,其认为自噬的激活对新生大鼠HIBD具有促进作用。相关研究发现,LC3(微管相关蛋白轻链3)可作为自噬形成标志物,当细胞受到某种刺激,位于细胞内的LC3-Ⅰ将脂质化为LC3-Ⅱ,参与形成自噬体,因此可根据分析LC3-Ⅱ/(LC3-Ⅰ+LC3-Ⅱ)的比值是否上调判断自噬体的形成[14]。还有研究证实,Beclin-1为除 LC3外与自噬密切相关的蛋白,主要参与自噬过程启动[15]。本研究结果发现,模型组中观察到皮质神经元中存在大量的自噬体和自噬溶酶体,其中泡状自噬体中包裹有可见的细胞质结构,同时与模型组比较,高剂量组、中剂量组、低剂量组中自噬体数量明显减少,且呈一定的剂量依赖性。同时本研究结果还发现,模型组中LC3-Ⅱ/(LC3-Ⅰ+LC3-Ⅱ)的比值和Beclin-1蛋白表达水平较假手术组均显著降低,表明缺氧缺血破坏神经元内稳定环境,造成蛋白错误修饰及细胞器损伤,从而通过上调Beclin-1蛋白表达水平启动自噬,LC3-Ⅰ脂质化,形成自噬体。与模型组比较,高剂量组、中剂量组、低剂量组新生大鼠脑组织中LC3-Ⅱ/(LC3-Ⅰ+LC3-Ⅱ)的比值和Beclin-1蛋白表达水平显著下降,且呈剂量依赖性,提示丹参酮ⅡA可通过抑制LC3-Ⅱ/(LC3-Ⅰ+LC3-Ⅱ)的比值和Beclin-1蛋白表达水平,抑制自噬过程。
Akt-mTOR信号转导通路是哺乳动物肿瘤免疫中的重要信号通路,其对调节细胞的生长、增殖、自噬以及凋亡有重要的作用[16]。Akt被相关因子激活磷酸化,发生空间转移,由胞浆转移至细胞核,进而调控下游如mTOR、Caspase-9、cyclin-D1等相应的信号分子,其中mTOR发生磷酸化激活下游p70S6K,促进核糖体蛋白S6发生磷酸化,从而调控细胞自噬,此调节过程与自噬的形成具有负相关性[17]。Makhov等通过荜茇明碱对mTOR信号影响的研究中发现,荜茇明碱能有效抑制细胞中Akt靶蛋白的磷酸化,下调Akt可有效抑制下游mTORC1的活性与细胞自噬[18]。本研究实验发现,与假手术组比较,模型组新生大鼠脑组织中p-Akt、p-mTOR、p-S6蛋白的表达水平均显著降低,提示Akt-mTOR信号转导通路参与调节神经元自噬,且呈负调节作用。与模型组比较,高剂量组、中剂量组、低剂量组新生大鼠脑组织中p-Akt、p-mTOR、p-S6蛋白表达水平均升高,并呈剂量依赖性,提示丹参酮ⅡA对Akt-mTOR信号转导通路具有正调节作用。
综上所述,丹参酮ⅡA对新生大鼠缺血缺氧性脑损伤具有显著的改善作用,大脑皮质神经元自噬受到显著的抑制,推测其机制可能为通过激活Akt-mTOR信号转导通路,抑制神经元自噬,从而改善脑损伤情况。然而,缺血缺氧性脑损伤是一个极其复杂的过程,其具体调节机制还有待进一步研究。
