脯氨酸引发提高烟草种子和幼苗抗逆性及其与抗氧化系统的关系
2019-01-15BOUDMYXAYKhampheng孙艳芝杨慧芹
BOUDMYXAY Khampheng,沈 镭 ,钟 帅 ,孙艳芝 ,杨慧芹
(1.中国科学院地理科学与资源研究所,北京100101;2.中国科学院大学,北京100049;3.云南师范大学马铃薯科学研究院,云南昆明650500)
植物在自然环境中会受到各种因子影响,如干旱、低温、高温、高盐等。不良环境作用于植物,将会引发植物体内一系列生理代谢反应,表现为生长和代谢受到抑制,严重时还会导致植株死亡,造成不可逆的伤害[1]。干旱和低温对世界植物产量的影响,在众多逆境因子中占首位,其危害程度相当于其他自然灾害之和[2]。
干旱和低温是影响植物正常生长发育的一个重要逆境[3]。前人研究发现,在干旱胁迫下糜子的光合色素含量和叶片MDA含量明显升高[4];在干旱胁迫下的木荷幼苗游离脯氨酸和MDA含量有所上升,抗旱性优良[5];随低温胁迫时间的延长,番茄叶片MDA,DHA,ASA含量呈现上升趋势,同时APX活性也有显著升高[6];在250 mmol/L NaCl胁迫下,加工番茄幼苗的MDA含量降低[7]。抗坏血酸(AsA)是一种普遍存在于植物组织的抗氧化物质,在活细胞中,抗坏血酸氧化还原系统由还原型的AsA和脱氢抗坏血酸(DHA)组成。AsA的氧化还原状态反映了植物细胞内外环境的氧化还原状况,当遭遇逆境胁迫时,AsA/DHA比率、相关代谢物和酶类会发生相应的变化。愈创木酚过氧化物酶(GPX)广泛存在于各种动、植物细胞中,它利用愈创木酚为电子供体清除H2O2。GPX存在于细胞的各个部位,它与SOD,CAT等酶类密切配合,相互协调清除植物体内过多的自由基,提高植物的抗逆性[6,8]。
尽管种子引发技术目前已在多种植物种类中得到成功应用,但在不同物种甚至同种不同批次的种子中,引发效果往往受到引发剂种类的影响。脯氨酸(Proline)是植物中主要的有机渗透调节物质之一,许多植物在受到逆境胁迫时都能大量积累脯氨酸,对细胞的渗透调节起重要作用,是植物体的防脱水良剂[9-10]。在干旱条件下,脯氨酸能维持渗透平衡和膜结构的稳定性,起到调节氧化还原状态和清除活性氧的作用[11],同时也能中和干旱胁迫产生的氨,起到解毒作用,还可作为复水后植物直接利用的氮源[12]。目前,尚未有关于脯氨酸引发提高烟草种子和幼苗抗逆性及其与抗氧化系统关系的报道。
本试验选100 mmol/L浓度的脯氨酸引发,研究其对常温条件、干旱、低温和干旱+低温交叉胁迫下烟草种子发芽和幼苗生长的影响,并深入了解抗氧化系统在脯氨酸引发提高烟草种子及幼苗抗逆性中的作用,期望从研究中获得有用信息,为提高烟草种子及幼苗的抗逆性提供参考依据。
1 材料和方法
1.1 材料
供试材料为云烟203裸种,由玉溪中烟种子有限责任公司提供。
1.2 种子引发处理
烟草种子以0.5%NaClO消毒5 min后,用蒸馏水漂洗干净,放于100mmol/L的Pro溶液中,在26℃黑暗条件下引发24 h。引发结束后,用蒸馏水快速冲洗种子,吸干表面水分,26℃下回干到原始含水量,于低温低湿贮藏柜中保存备用。未引发的干种子(萌发前用0.5%NaClO进行表面消毒)记为对照 1(CK1),蒸馏水引发作为对照 2(CK2)。
1.3 种子萌发试验
1.3.1 种子在正常培养条件下的萌发试验 将Pro引发的种子、未引发的干种子(CK1)及蒸馏水引发的种子(CK2)置于下铺1层海绵、上垫2张滤纸的直径为9 cm的培养皿中,每个培养皿中用播种器均匀播种100粒种子,加盖培养,发芽期间用自来水保持滤纸湿润。在26℃、空气相对湿度75%的人工气候箱中萌发,每天光照12 h(正常培养)。在第8,10,12,14,16 天测定幼苗的抗氧化系统。
1.3.2 种子在干旱胁迫条件下的萌发试验 以30%(W/V)PEG—6000作为模拟干旱胁迫条件。正常培养至第8天,然后换用30%的PEG—6000溶液浸润培养,处理2,4 d;处理4 d后恢复为用自来水培养2,4 d。在第10,12天测定干旱处理的抗氧化系统,第14,16天测定恢复处理的抗氧化系统。
1.3.3 种子在低温胁迫条件下的萌发试验 正常培养至第8天,然后置于4℃下低温胁迫处理培养2,4 d;处理4 d后恢复于26℃下培养2,4 d。在第10,12天测定低温处理的抗氧化系统,第14,16天测定恢复处理的抗氧化系统。
1.3.4 种子在干旱+低温交叉胁迫条件下的萌发试验 以25%(W/V)PEG—6000作为模拟干旱+低温交叉胁迫条件。正常培养至第8天,然后换用25%的PEG—6000溶液浸润培养并于4℃下模拟干旱+低温胁迫处理培养2,4 d;处理4 d后恢复为用自来水培养于26℃处理2,4 d。于第10,12天测定干旱+低温交叉胁迫处理的抗氧化系统;第14,16天测定恢复处理的抗氧化系统。
1.4 测定项目及方法
1.4.1 抗坏血酸过氧化物酶的测定 抗坏血酸过氧化物酶(APX)的测定参照李忠光等[13]的方法进行。反应混合液为pH值7.0的Tris-HCl缓冲液,内含0.1 mmol/L EDTA,0.1 mmol/L H2O2。测定时反应混合液和30 mmol/L AsA先于25℃水浴锅中预热,在试管中加入反应混合液2.9 mL,粗酶提取液50 μL,摇匀后调零,封口后于25℃水浴锅中保温5 min,然后加入AsA溶液50 μL,迅速混匀置于分光光度计中,每隔10 s读出290 nm处吸光度的减少值。APX的活力以每分钟消耗1 μmol AsA为一个酶活力单位来计算。
1.4.2 愈创木酚过氧化物酶的测定 愈创木酚过氧化物酶(GPX)的测定参照CHANCE等[14]和李忠光等[15]的方法进行。反应混合液为pH值7.0的Tris-HCl缓冲液,内含0.1 mmol/L EDTA,10 mmol/L愈创木酚,5 mmol/L H2O2。测定时反应混合液先于25℃水浴中预热,然后在试管中加入反应混合液2.95 mL,加入粗酶提取液50 μL,迅速混匀置于分光光度计中,每隔30 s读出470 nm处吸光度的减小值,取3 min反应时间来计算酶活。GPX的酶活力以每分钟消耗1 μmol愈创木酚的酶量为一个酶活力单位来计算。
1.4.3 丙二醛含量的测定 丙二醛(MDA)含量的测定参考张志良等[16]的方法,并对其进行了修改。具体步骤为:取0.2 g幼苗样品于研钵中加入液氮研磨,然后加5%的三氯乙酸(TCA)10 mL充分研磨后转入离心管中,用5 mL10%TCA清洗研钵,合并提取液并于4 000×g离心15 min。取2.0 mL上清液加入2.0 mL 0.25%硫代巴比妥酸(TBA)溶液(用10%TCA配制),100℃水浴锅加热30 min,冷却后在450,450,600 nm波长下测定吸光值。
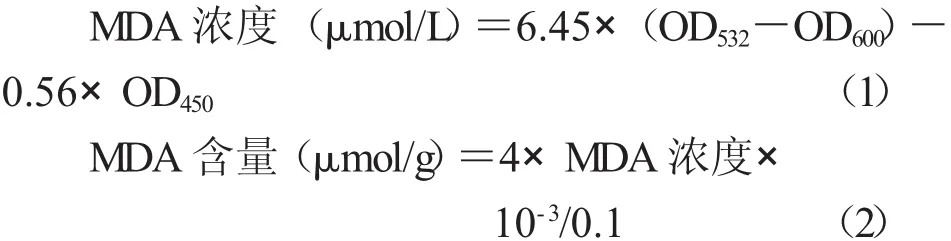
1.4.4 抗坏血酸和谷胱甘肽含量的测定
1.4.4.1 抗氧化剂的提取 抗坏血酸(AsA)和谷胱甘肽(GSH)的提取是按照李忠光等[13]的方法,取样品0.2g于研钵中,加入预冷的5%磺基水杨酸2.5mL和少许石英砂,置于冰上进行充分研磨,转入离心机于4℃下20 000×g离心20 min,将上清液分装于-20℃保存或直接进行抗氧化剂分析。
1.4.4.2 抗坏血酸含量的测定 AsA含量测定参照李忠光等[13]和JIANG等[17]的方法作如下修改:取上清液100 μL,加入20 μL1.84 mol/L三乙醇胺中和样液,加入pH值7.5的50 mmol/L磷酸缓冲液(内含2.5 mmol/LEDTA)250 mL,加入50 mmol/LDTT 50 μL,25℃保温10 min,再加入0.5%乙基马来酰亚胺50 μL,混匀后分别加入10%TCA,44%磷酸、4%双吡啶(用70%乙醇配制)各200 μL,混匀后加入3%FeCl3100 μL,混匀于40℃水浴锅中加热1 h,在525 nm处读出吸光度。还原型AsA的测定需把上述DTT和乙基马来酰亚胺用等体积的蒸馏水代替即可,用5%磺基水杨酸为溶剂,采取同样方法制作AsA标准曲线。
1.4.4.3 谷胱甘肽含量的测定 GSH含量的测定参照李忠光等[13]和NAGALASHMI等[18]的方法作如下修改:取上清液50μL,加入5%磺基水杨酸50μL,加入24μL1.84mol/L三乙醇胺中和样液,加入50μL 10%乙烯吡啶(用70%乙醇配制),25℃水浴1 h,然后加入706mL50mmol/L磷酸缓冲液(内含2.5mmol/L EDTA),再加入 20 μL10 mmol/LNADPH 和 80 μL 12.5 mmol/LDTNB(二硫硝基苯甲酸)混匀,于25℃保温 10 min,之后加入 20 μL 50 U/mol GR,立即混匀后读出3 min内412 nm吸光度的变化值。上述方法为GSSG含量的测定,总谷胱甘肽含量(GSH+GSSG)的测定将上述乙烯吡啶用等体积的蒸馏水代替即可,GSH含量用总谷胱甘肽含量(GSH+GSSG)减去GSSG含量即可得出。
1.5 统计分析
所有试验进行3次独立试验重复,每次独立试验有3个平行重复,每个重复有100粒种子。试验原始数据用SPSS 11.5统计软件分析,用Sigma plot 12.5作图。
2 结果与分析
2.1 脯氨酸引发对烟草幼苗抗氧化系统的影响
2.1.1 脯氨酸引发对干旱胁迫下烟草幼苗中APX和GPX活性的影响 图1-A显示,经100 mmol/L Pro引发后,烟草幼苗APX活力高于对照。对培养至第8天的幼苗进行胁迫处理,随着胁迫时间的延长,APX活力逐渐下降,明显低于正常培养条件下各对照。但经Pro引发的烟草幼苗APX活力仍显著高于经过胁迫的对照CK1和CK2;胁迫4 d后的烟草幼苗APX活力比经过胁迫处理后的对照CK1和CK2分别高出711%和30%,恢复后的幼苗APX活力逐渐升高,恢复生长4 d后APX活力比对照CK1和CK2分别高出383%和26%。从图1-B可以看出,经Pro引发后在正常条件的烟草幼苗GPX活力明显高于CK1和CK2(分别高58%和18%)。在干旱胁迫过程中,GPX活力随胁迫时间的延长而逐渐降低,明显低于正常培养条件各对照;胁迫4 d后,经Pro引发幼苗的GPX活性比经过胁迫处理后的对照CK1和CK2分别高出173%和44%;对胁迫处理的幼苗进行恢复,在恢复生长过程中Pro引发的烟草幼苗GPX活力逐渐升高,恢复生长4 d后,GPX活力比对照CK1和CK2分别明显高出449%和116%(P<0.05)。
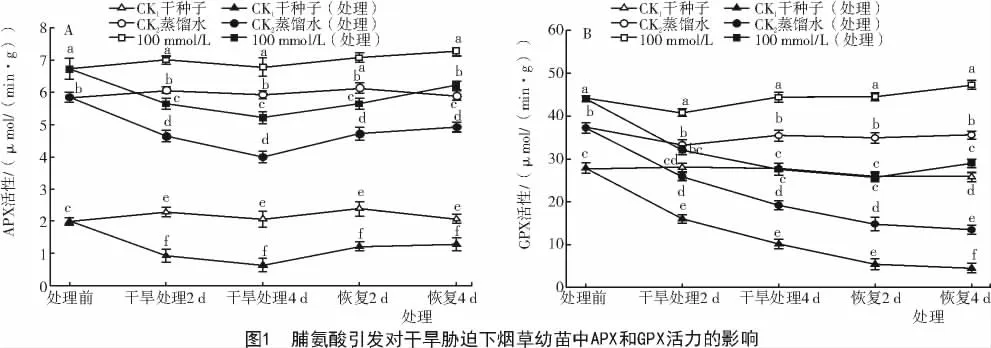
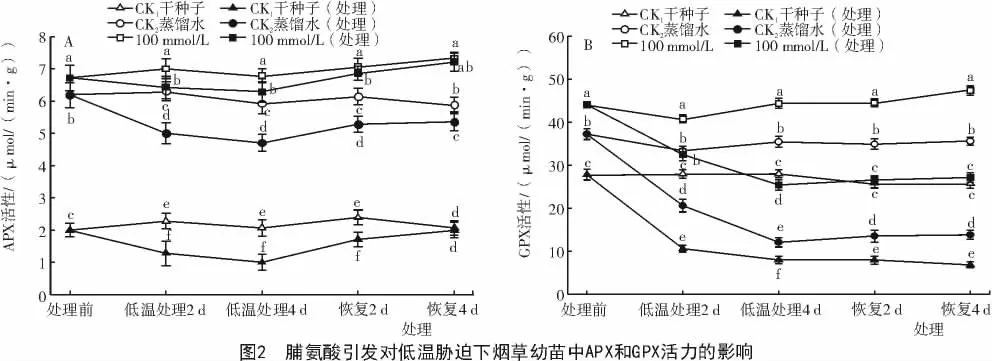
2.1.2 脯氨酸引发对低温胁迫下烟草幼苗中APX和GPX活性的影响 从图2-A可以看出,在正常萌发条件下,经Pro引发后的烟草幼苗APX活性明显高于对照。对培养至第8天的烟草幼苗进行低温胁迫处理,随着胁迫时间的延长APX活力逐渐下降。但经Pro引发过的幼苗APX活力仍明显高于经过胁迫的对照CK1和CK2,低温胁迫4 d后,APX活力比CK1和CK2分别高529%和33%;恢复生长4 d后,APX活力达到最高,分别比CK1和CK2高出261%和35%(P<0.05)。由图2-B可知,经Pro引发后的烟草幼苗GPX活力明显高于对照,在低温胁迫过程中GPX活力逐渐降低,但经Pro引发过种子的幼苗APX活力仍明显高于经过胁迫的对照CK1和CK2;胁迫4 d后,幼苗的GPX比对照CK1和CK2分别高出218%和110%;对胁迫处理后的烟草幼苗进行恢复,恢复4 d后,GPX活力明显提高,Pro引发的幼苗GPX活性分别比对照CK1和CK2高296%和 97%(P<0.05)。
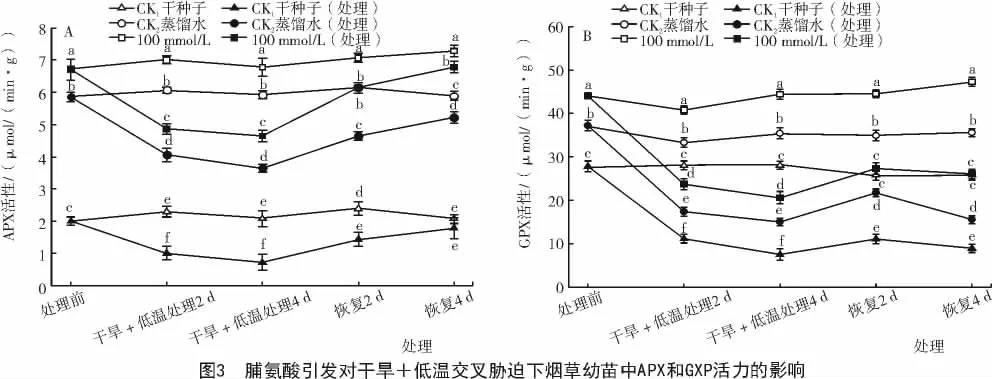
2.1.3 脯氨酸引发对干旱+低温交叉胁迫下烟草幼苗中APX和GPX活性的影响 由图3-A可知,在干旱+低温交叉胁迫下,烟草幼苗APX活性略有下降,但经Pro引发过种子的幼苗APX活力仍明显高于经过胁迫的对照CK1和CK2,干旱+低温交叉胁迫4 d后,APX活力明显提高,比经过胁迫的对照CK1和CK2分别高出550%和27%。恢复后,APX活性继续升高,恢复4 d后APX活力明显高于对照,分别比对照CK1和CK2高出280%和30%。由图3-B可知,在正常条件萌发条件下,经Pro引发后的烟草幼苗GPX活力都明显高于各对照。在干旱+低温交叉胁迫过程中GPX活力略有下降,在胁迫4 d后,经Pro引发幼苗GPX比对照CK1和CK2分别高出175%和37%;对胁迫处理后的烟草幼苗进行恢复,GPX活力逐渐升高,恢复4 d后,GPX活力比对照CK1和CK2分别高出192%和68%(P<0.05)。
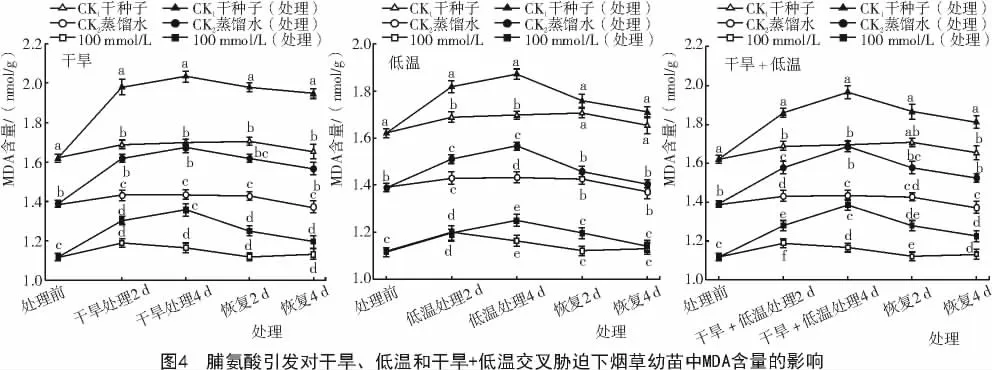
2.1.4 脯氨酸引发对干旱、低温和干旱+低温交叉胁迫下烟草幼苗中MDA含量的影响 由图4可知,在正常萌发条件下,经脯氨酸引发后的烟草幼苗MDA含量比对照干种子和蒸馏水引发的种子分别低42%和21%(P<0.05)。在受干旱、低温和干旱+低温交叉胁迫时,烟草幼苗中MDA含量快速增加,但经Pro引发的MDA含量仍低于对照。经恢复生长后,MDA含量逐渐降低。通过这些分析可知,用100 mmol/L Pro引发可以提高烟草幼苗在干旱、低温和干旱+低温交叉胁迫环境中抗氧化酶活性。
2.2 脯氨酸引发对烟草幼苗中AsA含量变化的影响
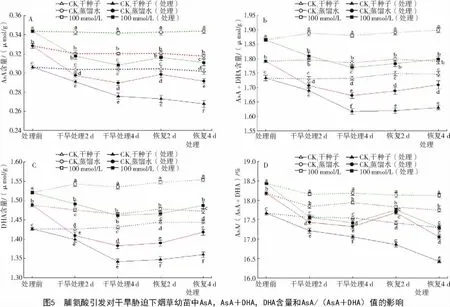
2.2.1 脯氨酸引发对干旱胁迫下烟草幼苗中AsA含量的影响 AsA是一种普遍存在于植物组织中的高丰度小分子抗氧化物质,它可以直接与ROS反应,在植物抵抗氧化胁迫中具有重要作用[19]。在正常萌发条件下,经Pro引发后的烟草幼苗AsA含量比对照 CK1和 CK2分别高 12.3%和 4.7%(P<0.05)。经过干旱胁迫后烟草幼苗AsA含量均明显低于各对照组,但经Pro引发的烟草幼苗AsA含量仍明显高于经过胁迫的对照CK1和CK2。从图5-A可以看出,经干旱胁迫的烟草幼苗还原型抗坏血酸(AsA)的含量显著增加,干旱胁迫4 d时,烟草幼苗的AsA含量逐渐降低,但经Pro引发的ASA仍比对照CK1和CK2分别高出11.8%和6.4%。对胁迫处理4 d后的幼苗进行恢复,恢复2 d后烟草幼苗的AsA含量逐渐升高,但恢复到4 d时,AsA含量又呈降低趋势,但经Pro引发的AsA含量仍比经过胁迫的对照 CK1和 CK2分别高出16.1%和 6.7%(P<0.05)。经过干旱胁迫的烟草幼苗氧化性抗坏血酸(DHA)和总抗坏血酸含量AsA+DHA也随之下降,但经Pro引发的DHA,AsA+DHA含量仍明显高于经过胁迫的对照CK1和CK2(图5-B,C)。经干旱胁迫2 d的烟草幼苗在遭遇后续干旱胁迫4 d时,AsA/(AsA+DHA)明显高于对照(图5-D);对幼苗恢复生长4 d时,还原型抗坏血酸在总含量中的比值分别高出对照CK1和CK25.3%和1.4%。
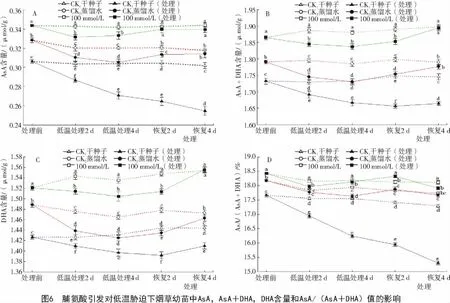
2.2.2 脯氨酸引发对低温胁迫下烟草幼苗中AsA含量的影响 从图6-A可以看出,经低温胁迫的烟草幼苗AsA含量都明显高于对照,并且4℃胁迫4 d时,Pro引发的作用效果明显高于经过胁迫的对照CK1和CK2。在低温胁迫4 d后经Pro引发的烟草幼苗AsA含量比CK1和CK2分别高出23%和9.3%;恢复生长4 d后,AsA含量也高出CK1和CK233%和8.1%。此外,经低温胁迫的DHA含量和总含量也高出对照(图6-B,C)。从图6-D可以看出,低温胁迫处理4 d后,烟草幼苗AsA/(AsA+DHA)的值比CK1和CK2分别高出12%和2.9%;恢复生长4d后,AsA/(AsA+DHA)的值比对照CK1和CK2明显高出 17%和 1.4%(P<0.05)。
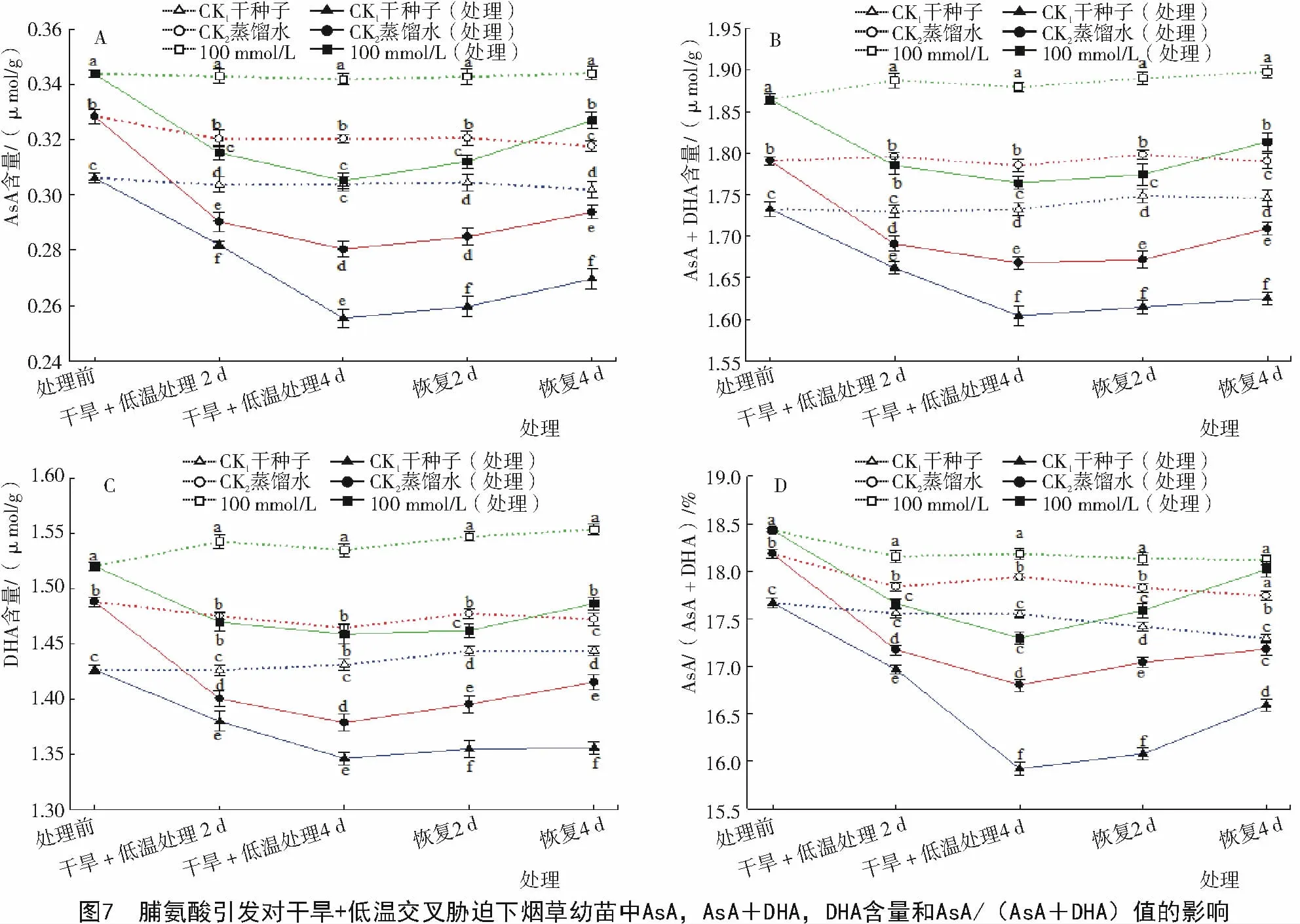
2.2.3 脯氨酸引发对干旱+低温交叉胁迫下烟草幼苗中AsA含量的影响 从图7可以看出,经干旱+低温交叉胁迫诱导,经Pro引发的烟草幼苗AsA,DHA含量和总含量明显高于对照(图7-A,B,C)。干旱+低温交叉胁迫前,烟草幼苗还原型抗坏血酸在总含量中的比值AsA/(AsA+DHA)最高(图7-D);干旱+低温交叉胁迫处理4 d时,经Pro引发的烟草幼苗AsA/(AsA+DHA)的值比CK1和CK2分别高出8.6%和2.9%,恢复生长4 d时,AsA/(AsA+DHA)的值比对照CK1和CK2分别高出8.6%和4.9%。
2.3 脯氨酸引发对烟草幼苗GSH含量变化的影响
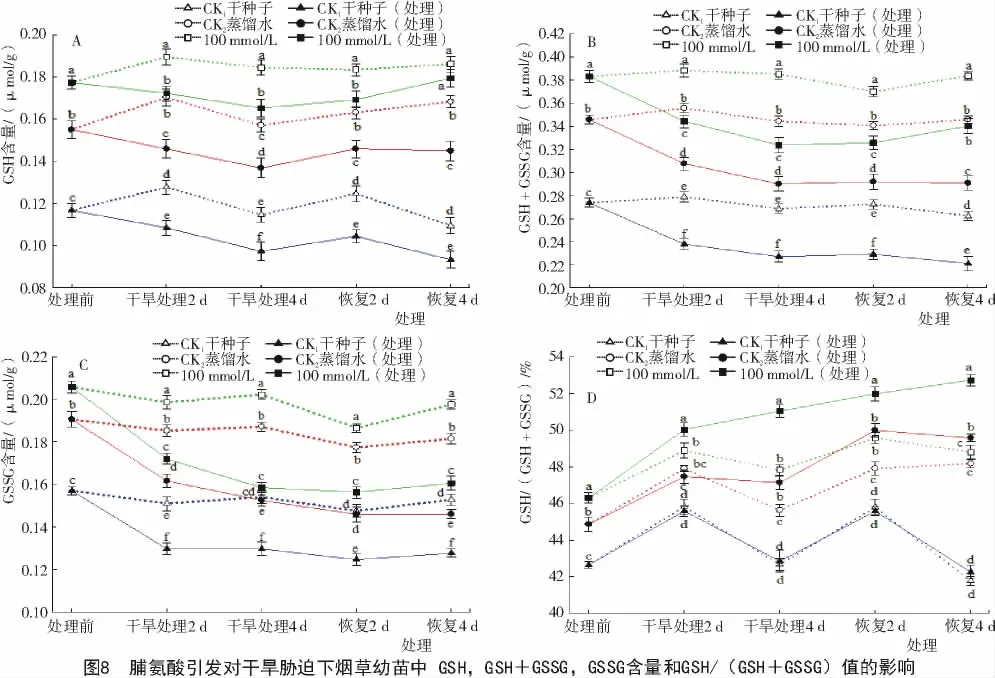
2.3.1 脯氨酸引发对干旱胁迫下烟草幼苗中GSH含量变化的影响 GSH广泛分布于哺乳动物、植物和微生物细胞内,是最主要的、含量最丰富的含巯基的低分子肽,它可以直接或间接与ROS反应[20]。由图8可知,在正常萌发条件下,经100 mmol/L Pro引发后烟草幼苗GSH,SGH+GSSG,GSSG含量比对照CK1和CK2高。经干旱胁迫,Pro引发的烟草幼苗GSH,SGH+GSSG和GSSG含量均明显低于各对照组,但比经过胁迫的对照CK1和CK2要高(图8-A,B,C)。经引发后的烟草种子在干旱胁迫下的GSH/(GSH+GSSG)值明显增大(图8-D),经干旱胁迫时烟草幼苗GSH/(GSH+GSSG)值明显高出各对照组。干旱胁迫处理4 d时,Pro引发的烟草幼苗GSH/(GSH+GSSG)的值比经过胁迫的对照CK1和CK2分别高出19%和8.1%(P<0.05);恢复生长4d时,GSH/(GSH+GSSG)的值明显高出CK1和CK2(24.8%和5.7%),可以看出,100 mmol/LPro引发的烟草幼苗在干旱胁迫下还原性谷胱甘肽的含量明显增加。
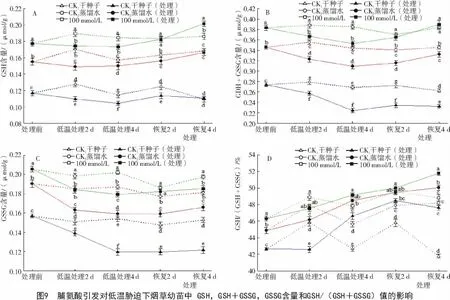
2.3.2 脯氨酸引发对低温胁迫下烟草幼苗中GSH含量变化的影响 从图9可以看出,经低温胁迫后烟草幼苗GSH,GSH+GSSG,GSSG含量均明显低于各对照组,但比 CK1和 CK2要高(图 9-A,B,C)。经100 mmol/L Pro引发的烟草幼苗在低温胁迫下的 GSH/(GSH+GSSG)值明显提高(图 9-D),高于各对照组。低温胁迫处理4 d后,经Pro引发的烟草幼苗GSH/(GSH+GSSG)的值比经过胁迫的对照CK1和 CK2分别高出 5.4%和 1.4%(P<0.05);恢复生长4 d时,GSH/(GSH+GSSG)的值比经过胁迫的对照CK1和CK2分别高出8.7%和3.4%(P<0.05),表明Pro引发能提高烟草幼苗在低温胁迫下还原性谷胱甘肽的含量。
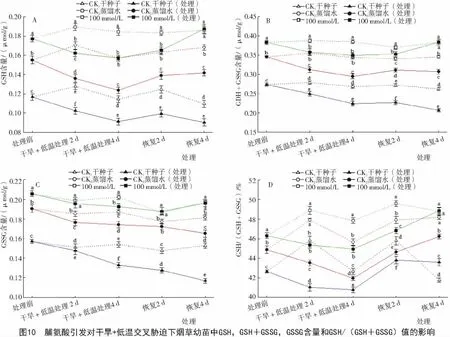
2.3.3 脯氨酸引发对干旱+低温交叉胁迫下烟草幼苗中GSH含量的影响 由图10可知,在正常萌发条件下,经Pro引发后烟草幼苗的谷胱甘肽含量比对照CK1和CK2高。经干旱+低温交叉胁迫的烟草幼苗谷胱甘肽含量均明显低于各对照组,但经Pro引发的烟草幼苗谷胱甘肽含量仍明显高于经过胁迫的对照CK1和CK2。分析结果说明,100 mmol/L Pro引发能提高烟草幼苗在干旱+低温交叉胁迫下的谷胱甘肽含量。
3 结论与讨论
抗坏血酸和谷胱甘肽在活性氧脱毒过程中起重要作用,它们可直接同ROS反应,将其还原;又可作为酶的底物,在活性氧的清除过程中扮演重要角色[2]。抗坏血酸、还原型谷胱甘肽及APX,GR,SOD和MDHAR都参与了植物细胞中抗氧化剂的再生过程。抗坏血酸的第3种功能是在叶绿体类囊体表面作为还原剂,参与抗坏血酸过氧化物酶介入的过氧化氢的清除,在这一过程中,抗坏血酸被氧化成单脱氢抗坏血酸。在完全氧化的脱氢抗坏血酸产生的情况下,还原型谷胱甘肽作为还原剂参与抗坏血酸的再生,产生的氧化型谷胱甘肽可以通过谷胱甘肽还原酶得到还原。由于抗坏血酸在清除活性氧的过程中发挥了重要作用,故而在叶绿体中,氧的摄入、APX的功能及分子氧还原等相关的过程都有其参与[2]。
本试验研究结果表明,在正常萌发条件下,用100 mmol/L Pro引发后,烟草幼苗抗氧化酶活性(APX和GPX)和抗氧化剂(AsA和GSH)都明显高于对照。对烟草幼苗进行干旱、低温或干旱+低温交叉胁迫处理,随着胁迫时间的延长,抗氧化酶活性逐渐降低,抗氧化剂含量逐渐下降,但经Pro引发的烟草幼苗抗氧化酶活性和抗氧化剂含量仍明显高于经过胁迫的对照CK1和CK2。以上结果表明,Pro引发可明显改善烟草幼苗抗氧化系统,包括抗氧化剂含量和抗氧化酶活性,使幼苗在干旱、低温或干旱+低温交叉胁迫下有较强的抗旱和抗寒性,抗氧化系统在这个过程中发挥了重要的作用。
植物体内的抗氧化酶活性越高,MDA含量越低[7,21]。烟草幼苗在干旱胁迫、低温或干旱+低温交叉胁迫下,MDA含量快速增加,表明幼苗体内细胞发生了膜脂过氧化反应,细胞膜的完整性受到破坏,但经100 mmol/L Pro引发的烟草幼苗MDA含量明显低于对照,说明Pro引发在很大程度上减轻了逆境胁迫引起的伤害,提高了烟草的抗旱和抗旱能力。
