基于农艺性状和品质性状的燕麦属物种遗传多样性分析
2018-12-05,,,,,,
, , , , , ,
(四川农业大学小麦研究所, 成都 611130)
六倍体栽培燕麦(AvenasativaL.)隶属于禾本科(Poaceae),燕麦族(Aveneae),燕麦属(AvenaL.)是一种重要的粮草兼用型作物,富含β-葡聚糖、蛋白质、膳食纤维以及不饱和脂肪酸,具有很高的营养价值,对心脑血管疾病有积极作用[1-4]。同时,燕麦具有较强的抗逆能力,不与小麦稻谷等主要粮食作物争夺耕地,以及农业风险系数低等优点,有较大的发展潜质。我国是燕麦的起源中心之一,燕麦种植历史悠久,至今仍是内蒙古、山西、河北等地区大面积种植的粮食作物之一。
燕麦野生种质资源是燕麦育种的重要基因库。燕麦属中包含大约29个物种[5-7],这些物种分布范围较广,遗传多样性十分丰富,许多种质资源具有极强的抗病能力,如抗燕麦白粉病[8],同时在许多农艺性状上表现出丰富的变异,适用于不同育种目标的燕麦新品种选育。尽管野生燕麦资源中含有许多优异基因,但目前应用于育种的材料还不多。这主要归咎于不同物种间存在生殖隔离,以及严重的连锁累赘效应。同时,缺乏对收集到的野生燕麦种质资源相关农艺性状和品质性状的系统评价也是造成利用较少的重要原因。目前对于野生燕麦种质资源的系统评价主要集中于抗逆性与抗病性方面,而在其它农艺性状以及品质性状的系统评价还未多见。
因此,本研究选择代表燕麦属24个物种的116份材料,针对11个农艺性状和3个品质性状进行系统分析和评价,为燕麦育种亲本选择提供依据。
1 材料与方法
1.1 试验材料
本试验收集了来自世界各地的代表燕麦属24个物种(二倍体13个,四倍体7个,六倍体4个)共计116份材料用于本研究(表1),所有供试材料连续2年(2010—2011年,2011—2012年)种植于成都温江四川农业大学实验基地(30°41′N,103°49′E),进行相关农艺性状和品质性状调查。所有材料均于10月底播种次年6月初收获。田间试验采用随机区组设计,3次重复,双行区,行长2 m,行距0.30 m,田间栽培管理与大田一致。材料的具体信息见表1。
表1 供试116份燕麦材料
续表1

种名种型基因组样品号a来源地A.barbata野生种ABPI296229以色列PI337802土耳其PI337826希腊PI282723以色列PI337731希腊PI367322葡萄牙A.brevis野生种AsCiav1783德国:萨克森Ciav9113欧洲PI258545葡萄牙A.canariensis野生种AcCN23017加那利群岛CN23029加那利群岛CN25442加那利群岛CN26172加那利群岛CN26195加那利群岛A.clauda野生种CpCN19205伊朗CN19242土耳其CN21378希腊CN21388阿尔及利亚CN24695土耳其A.damascena野生种AdCN19457叙利亚CN19458叙利亚A.eriantha野生种CpCiav9050英国PI367381西班牙:马德里A.fatua野生种ACDPI447299中国:甘肃PI544659美国PI545459墨西哥PI560776土耳其A.hirtula野生种AsCN19739阿尔及利亚CN21674法国:科西嘉CN21703摩洛哥CN22633突尼斯A.hispanica野生种AsCN25676葡萄牙CN25727葡萄牙CN25766葡萄牙CN25778葡萄牙CN25787葡萄牙A.longiglumis野生种AsCiav9087阿尔及利亚Ciav9089利比亚PI367389葡萄牙A.lusitanica野生种AsCN25414西班牙CN25885摩洛哥CN25899葡萄牙CN26441西班牙A.maroccana野生种ACCiav8330摩洛哥Ciav8331摩洛哥A.murphyi野生种ACCN21989西班牙CN25974摩洛哥
续表1

种名种型基因组样品号a来源地A.sativassp.nuda栽培种ACDCiav9009加拿大:安大略PI401795荷兰PI401812德国A.occidentalis野生种ACDCN4547加那利群岛CN23036加那利群岛CN25942摩洛哥CN25956摩洛哥CN26226加那利群岛A.sativa栽培种ACDPI40650中国:甘肃Ciav1946澳大利亚PI93751中国:西藏PI103669乌拉圭PI103670澳大利亚PI175021澳大利亚PI194896印度PI258641埃塞俄比亚PI258644格鲁吉亚PI258649中国PI258655格鲁吉亚PI258656乌克兰PI258657蒙古PI258658波兰PI258659蒙古:扎布汗PI258663乌克兰PI258665波兰PI258666亚美尼亚PI258677乌克兰:克里米亚PI258685阿尔卡利亚PI258724俄罗斯PI258726土耳其:安卡拉PI258734俄罗斯PI411426土耳其PI411439土耳其PI636013匈牙利PI636073土耳其:内夫谢希尔CN2811土耳其:约兹加特CN21957匈牙利:赫维什CN24942南澳大利亚CN53095新南威尔士CN64226埃塞尔比亚CN82122伊朗A.sterilis野生种ACDPI411503阿尔及利亚PI411656埃塞俄比亚A.strigosa野生种AsPI83722澳大利亚PI158246西班牙:卢戈Ciav9066加拿大:安大略PI436082智利:拉各斯A.vaviloviana野生种ABPI412761厄立特里亚PI412766埃塞俄比亚
续表1
种名种型基因组样品号a来源地A.ventricosa野生种CvCN21405阿尔及利亚A.wiestii野生种AsPI53626埃及:吉萨Ciav9053加拿大:安大略PI299112智利A.insularis野生种ACsn意大利:西西里岛6-B-22意大利:西西里岛INS-4意大利:西西里岛
注:a居群编号以PI开头的材料来自美国国家种质资源库(USDA),其余材料均来自加拿大农业和农业食品部(AAFC)。
表2 供试燕麦材料来源地纬度分类

纬度分类包含地区北纬20°、南纬20°埃塞俄比亚,厄立特里亚,墨西哥,巴西,智利北纬20°~40°、南纬20°~40°摩洛哥,中国,乌拉圭,澳大利亚,以色列,希腊,葡萄牙,利比亚,加那利群岛,伊朗,意大利,埃及,阿尔及利亚,叙利亚,突尼斯,印度北纬40°以上及高寒地区土耳其,德国萨克森,英国,西班牙,加拿大,美国,瑞典,荷兰,法国,格鲁吉亚,乌克兰,俄国,匈牙利,蒙古,波兰,白俄罗斯,亚美尼亚
注:纬度划分参照伍光和等[9]的方法。
1.2 燕麦籽粒品质性状测定
品质指标的测定:蛋白质含量采用凯氏定氮法[10]测定;脂肪含量采用索氏抽提法测定[11];β-葡聚糖含量采用EBC方法[12]测定。
1.3 田间农艺性状统计
田间随机选择3株植株调查单株芒性、芒型、芒色、主穗小穗数;收获时每小区取样3株室内考种,调查单穗粒重、单株粒重、千粒重、粒色、粒形、粒茸毛和粒饱满度。统计分析的性状分为2类:第1类是质量性状,包括芒性、芒型、芒色、粒色、粒形、粒茸毛、粒饱满度7个性状;第2类是数量性状,包括单主穗小穗数、单株粒重、单穗粒重、千粒重4个性状。
1.4 数据分析
表型数据的方差分析及相关性分析使用SPSS 22.0软件,其中方差分析的变异来源分类标准为:1) 染色体倍性,包括二倍体、四倍体、六倍体; 2) 物种(表1),包括24个种; 3) 基因组类型(表1),包括A基因组二倍体、C基因组二倍体、AB基因组四倍体、AC基因组四倍体和ACD基因组六倍体共计5种类型; 4) 来源地纬度(表2 ),包含3种类型; 5) 驯化程度(表1),分为栽培材料和野生材料2种,其中栽培材料为Avena sativa,野生材料则为剩余的23个种。
表5 栽培材料和野生材料4个产量性状的平均数、标准差、变异系数和多样性指数

性状材料类型材料份数最小值最大值平均数标准差变异系数CV(%)多样性指数H’主穗小穗数栽培材料2656.4253.2119.44651.203142.871.8577野生材料7919.2185.277.559543.080455.551.9038主穗粒重(g)栽培材料330.422.471.02390.486748.001.3379野生材料800.011.310.41650.329779.161.7799单株粒重(g)栽培材料346.12529.96614.9826.82745.571.8000野生材料800.23426.5539.11805.745963.021.9374千粒重(g)栽培材料252.64625.30810.60856.845464.531.8124野生材料790.81332.0586.79146.063789.291.6094
2 结果与分析
2.1 供试燕麦物种品质性状方差分析
用SPSS 22.0软件对燕麦所有倍性、物种、基因组、来源地纬度驯化程度进行了方差分析,从表3可以看出,供试燕麦材料的基因组类型、物种和驯化程度对被测3个品质性状影响呈极显著(p<0.01);同时,供试燕麦材料的染色体倍性对蛋白质和脂肪含量也有极显著影响(p<0.01);但供试燕麦材料的纬度分布与被测3个品质性状的影响不显著(p>0.05)。由于驯化程度对供试燕麦物种的3个品质性状的影响最大。因此,本研究主要对不同驯化程度对供试燕麦品质性状的影响进行深入分析。
表3 供试燕麦品质性状方差分析
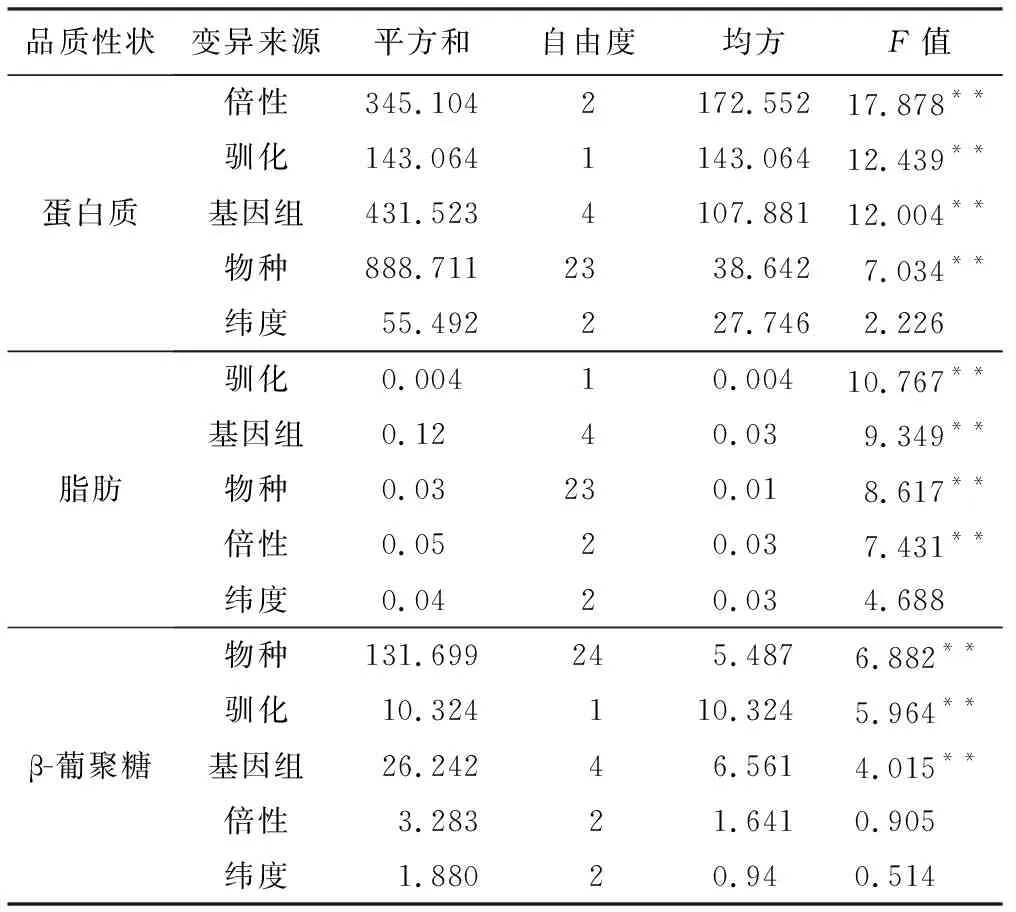
品质性状变异来源平方和自由度均方F值蛋白质倍性345.1042172.55217.878**驯化143.0641143.06412.439**基因组431.5234107.88112.004**物种888.7112338.6427.034**纬度55.492227.7462.226脂肪驯化0.00410.00410.767**基因组0.1240.039.349**物种0.03230.018.617**倍性0.0520.037.431**纬度0.0420.034.688β-葡聚糖物种131.699245.4876.882**驯化10.324110.3245.964**基因组26.24246.5614.015**倍性3.28321.6410.905纬度1.88020.940.514
注:**代表差异极显著,*代表差异显著。
2.2 燕麦物种农艺性状多样性分析
由表4可以看出,粒型、粒色和粒茸毛3个性状,栽培材料的多样性指数都较野生材料低,而在其他4个性状上,栽培材料的多样性指数均高于野生材料。单独分析野生材料发现,籽粒大多表现为较饱满、少茸毛、黑色,粒型以纺锤形为主,有少数为椭圆型籽粒,其余均为长筒形;而栽培材料的籽粒大多表现为较饱满、多茸毛、黄色,粒型以长筒形为主,同样存在少量椭圆形籽粒,其余均为纺锤形。两组材料在芒的性状上差异最大,其中芒型的H’差异最大,达到了91.3%,其次是芒色,H’差异为90.1%。野生材料几乎全都表现为黑色、弯曲的强芒性,而栽培材料主要表现为黑色、弯曲的弱芒性,并且出现了无芒的性状,直芒和黄色芒类型的比例也较野生材料有所上升。
表4 燕麦栽培材料和野生材料7个穗部性状的频率分布和多样性指数

性状材料类型材料份数频率分布123多样性指数H’粒型栽培材料340.3530.5590.0880.9070野生材料800.5130.3750.1130.9566粒色栽培材料340.8240.1760.4660野生材料800.1880.8130.4826粒茸毛栽培材料340.1180.2650.6180.6491野生材料800.4380.3750.1881.0436粒饱满度栽培材料340.1180.4120.4710.9721野生材料800.0630.4630.4750.8843芒性栽培材料340.1470.5880.2650.9460野生材料800.1880.8130.4825芒型栽培材料290.3790.6210.6636野生材料800.0130.9980.0585芒色栽培材料290.7240.2760.5903野生材料800.9980.0130.0585
注:粒型:1-纺锤形,2-长筒形,3-椭圆形;粒色:1-黄色,2-黑色;粒茸毛:1-少,2-中,3-多;粒饱满度:1-干瘪,2-中等,3-饱满;芒性:1-无,2-弱,3-强;芒型:1-直,2-弯曲;芒色:1-黑色,2-黄色。
表6 栽培材料和野生材料主要品质性状的平均数、标准差、变异系数和多样性指数

性状材料类型材料份数最小值最大值平均数标准差变异系数CV(%)多样性指数H’β-葡聚糖含量(%)栽培材料331.66.3541.025625.641.9574野生材料791.158.374.56671.418231.051.9807蛋白质含量(%)栽培材料3012.5223.0517.4322.438413.991.9056野生材料7514.8326.6821.13492.802113.262.0496脂肪含量(%)栽培材料310.030.090.060.0226.001.7978野生材料770.040.120.080.0223.001.9468
表7 基于数量性状平均值和质量性状赋值的相关系数

粒型粒色粒茸毛粒饱满度葡聚糖蛋白质脂肪芒性芒型芒色小穗数主穗粒重单株粒重千粒重粒型10.1050.488**0.172-0.042-0.1450.104-0.120-0.024-0.059-0.1370.0330.0930.211*粒色10.138-0.053-0.064-0.154-0.048-0.1190.1070.177-0.0080.0120.0240.208*粒茸毛10.055-0.071-0.245*0.033-0.250**-0.1660.096-0.1280.234*0.192*0.393**粒饱满度10.027-0.095-0.264**-0.030-0.009-0.1030.1590.1780.1520.043葡聚糖10.166-0.0970.209*0.1550.0480.192-0.094-0.027-0.221*蛋白质10.401**0.392**0.257**-0.011-0.282**-0.504**-0.423**-0.192脂肪10.338**0.350**-0.101-0.521**-0.349**-0.1350.021芒性10.601**0.158-0.079-0.195*-0.079-0.086芒型10.447**-0.217*-0.0680.0290.072芒色10.214*0.1180.0600.024小穗数10.261**0.164-0.379**主穗粒重10.794**0.662**单株粒重10.587**千粒重1
注:**代表相关性极显著,*代表相关性显著。
从表5可看出,栽培材料和野生材料各产量相关的农艺性状的平均数、标准差、变异系数的H’均存在较大变异。以变异系数为例,栽培材料和野生材料4个性状的CV变幅分别为42.87%~64.53%和55.55%~89.29%。2个类型的材料各性状的变幅极广,主穗小穗数19~253粒,主穗粒重0.42~2.47 g,单株粒重0.234~29.966 g,千粒重0.813~32.058 g。野生材料的主穗小穗数、单穗粒重、单株粒重3个性状H’较栽培材料高,增加的程度单穗粒重最大,达到了24.83%;而千粒重栽培材料H’比野生材料高。2个类型变异系数的变化趋势与H’一致。
2.3 燕麦主要品质性状多样性分析
从表6可看出,栽培材料和野生材料的β-葡聚糖含量和脂肪含量的平均数、标准差、变异系数的H’均存在较大变异,而蛋白质含量的各数据差异不大。以变异系数为例,栽培材料和野生材料3个性状的CV变幅分别为13.99%~64.53%和13.26%~89.29%。2个类型的材料各性状的变幅极广,β-葡聚糖含量1.15%~8.37%,蛋白质含量12.52%~26.68%,脂肪含量3%~12%。野生材料的β-葡聚糖含量、蛋白质含量、脂肪含量3个性状H’较栽培材料高。
2.4 燕麦属物种品质性状与农艺性状的简单相关性分析
通过对燕麦主要农艺性状和品质性状的简单相关性分析可以确定燕麦高产和优质的特征。对数量性状,取其平均值进行计算,对质量性状则进行赋值(见表4)。相关分析结果(表7)表明,燕麦粒茸毛与芒性、蛋白质含量均呈极显著负相关(p<0.01),粒饱满度与脂肪含量呈极显著负相关(p<0.01),芒性与主穗粒重、主穗小穗数均呈显著负相关(p<0.05),β-葡聚糖含量与千粒重呈显著负相关(p<0.05),蛋白质含量和脂肪含量均与主穗小穗数、单穗粒重呈极显著负相关(p<0.01),此外蛋白质含量还与单株粒重呈极显著负相关(p<0.01),主穗小穗数与千粒重呈极显著负相关(p<0.01);粒型与粒茸毛呈极显著相关性,粒型、粒色、粒茸毛均与千粒重显著相关,葡聚糖含量与芒性显著正相关,蛋白质含量和脂肪含量与芒性、芒型均为极显著正相关关系,芒性与芒型、芒性与芒色均为极显著相关,芒色与主穗小穗数显著相关。蛋白质含量和脂肪含量之间呈极显著的正相关,主穗小穗数与单穗粒重呈极显著正相关,单株粒重、单穗粒重和千粒重之间均为极显著的正相关关系。
3 讨论与结论
随着科学膳食观念的普及,燕麦作为一种重要的健康和功能食品在市场上的需求逐年上升[13],燕麦相对于小麦、水稻等主粮作物而言产量较低,但是在诸如蛋白质含量、β-葡聚糖含量等营养品质上有很大的优势[14],如何综合提高燕麦的产量和品质成为了当下燕麦生产研究中的重要问题。燕麦种质资源遗传多样性是燕麦育种的基础,对燕麦种质资源进行系统评价,是充分利用目标性状突出的燕麦种质资源的前提,基于此,我们对代表24个物种的116份燕麦材料的11个农艺性状和3个品质性状进行了系统分析。
3.1 燕麦野生资源农艺性状和品质性状的遗传多样性
研究表明,在世界范围内不同的燕麦物种中存在着很高的遗传多样性,但是针对六倍体栽培燕麦品种间的遗传多样性研究表明,栽培燕麦品种间的遗传多样性差异不大[15]。研究结果也表明,在不同物种的燕麦材料中,遗传多样性差异主要来源于驯化程度的不同,即野生材料和栽培材料之间的差异,而在相同的驯化程度分组内差异不显著。随着驯化程度的提高,占主导地位的穗部性状从带弯曲强芒的黑色、少绒毛的纺锤形籽粒变为带弯曲弱芒的黄色、多茸毛的长筒形籽粒,进而出现了无芒的材料;而品质性状方面,脂肪含量、β-葡聚糖含量和蛋白质含量随着驯化程度的提高而下降,其中脂肪含量的下降可能是育种家考虑到在燕麦储藏、运输和加工过程中,过高的脂肪含量都有不利的影响[16]。
3.2 燕麦属物种野生材料和栽培材料的遗传多样性分析
通过对栽培材料和野生材料的遗传多样性对比分析发现,除了芒和千粒重在栽培材料中的遗传多样性高于野生材料外,其余的农艺性状和品质性状均为野生材料中遗传多样性较高。这一结果表明,到目前为止燕麦的育种工作虽然集中于籽粒产量上,但是在“产量三要素”即单位面积穗数、每穗粒数和千粒重中侧重了对千粒重的选择,而对每穗粒数(主穗粒重/主穗小穗数)的性状关注程度较低。同时,在野生材料尤其是二倍体材料中存在着很多具有优良品质性状的种质资源,对比我国各地生产上主栽燕麦品种的品质性状的统计结果,高β-葡聚糖含量的材料共有17个(≥5.87%),其中最突出的是As基因组二倍体A.atlantica,其β-葡聚糖含量为6.32%~8.37%;高脂肪(≥8%)的材料共有47个,其中最突出的是Cp基因组二倍体A.clauda和A.eriantha,其脂肪含量高达11%~12%;高蛋白质含量(≥18%)的材料共有75个,其中最突出的是23.18%~26.68%[17-18]。由此可见,现有的栽培燕麦品种在品质性状上还有很大的提升空间,由于品质性状在栽培燕麦中遗传多样性较低,所以要想提高栽培燕麦的品质性状,需要充分利用丰富的燕麦野生资源。
3.3 燕麦属物种品质性状和农艺性状的关系
为了综合的考虑燕麦育种过程中对于营养品质和籽粒产量的选择,我们对燕麦籽粒的品质性状和农艺性状进行了相关性分析,分析结果显示,蛋白质含量和脂肪含量之间呈极显著的正相关(p<0.01),而前人的研究结果存有争议,Magne等的研究结果表明,蛋白质含量与脂肪含量之间不存在显著性的关联,Forsberg等的结果却认为,蛋白质含量与脂肪含量之间存在极显著的正相关[19]。燕麦籽粒的品质性状即β-葡聚糖含量、脂肪含量、蛋白质含量与产量性状之间均存在显著的负相关,这一结果与Forsbrg等[19]的研究结果一致,但是也有学者的研究结果认为脂肪含量与产量无关[20-23]。与此同时,燕麦籽粒的葡聚糖含量、蛋白质含量和脂肪含量与芒性、芒型均显著或极显著正相关,这与我们的遗传多样性研究的结果是一致的,燕麦属物种随着驯化程度的增加,芒的性状逐渐转变为弱芒、无芒,各品质性状的含量也有所下降,进一步的表明了芒与燕麦属物种的品质性状具有极显著的关联性。在产量性状之间的相关性中可以发现,主穗小穗数与单穗粒重有正相关关系,这与Buerstmayr[24]和Tang等[25]的结果是一致的,但与Dumlupinar等的结果存在矛盾,后者认为主穗小穗数和单穗粒重呈极显著负相关[26];此外,Frey等认为小穗数与千粒重之间存在负相关的关系,当千粒重位于27.5~31 g的范围内,小穗数和千粒重的关系能达到平衡,整体的燕麦籽粒产量达到峰值[27],本研究也表明主穗小穗数与千粒重呈极显著的负相关,这可能与植物体内营养分配有关系,二者在营养成分上具有竞争关系。
4 结 论
本研究结合前人对燕麦种质资源的农艺性状、抗旱性等方面的研究结果[25,28],选择出了部分具有优质综合性状的材料,可用以改良燕麦的品质、产量性状和抗逆性。包括CN 25859(A.atlantica), CN 21703(A.hirtula),PI 367381(A.eriantha),PI 258734和PI 258724(A.sativa)。同时,燕麦的籽粒产量和营养品质总体呈负相关的关系,在育种工作中,过于侧重某一个指标时,反而会导致整体的价值下降。因此,在育种工作中,要综合考虑材料的产量性状和品质性状之间的相关性,以选育出整体价值高的材料或者专用型品种。此外,在燕麦属物种中,芒的性状与品质性状紧密关联,品质性状表现较好的材料通常具有强芒性,在田间较容易识别,可以作为育种选择过程中的一个表型标记使用。
