植物单宁的生物活性研究进展
2018-09-22赵韩栋焦文晓范新光姜微波
舒 畅,赵韩栋,焦文晓,范新光,姜微波
(中国农业大学食品科学与营养工程学院,北京 100083)
单宁(tannins)是一类高分子量水溶性多酚类化合物的总称,广泛分布于植物的叶、根、果实和表皮等不同组织中。其分子量在500~3000 Da之间,一些缩合单宁的高聚体分子量可超过3000 Da[1]。单宁中含有大量羟基,可形成分子间和分子内氢键,能够与大分子化合物如蛋白质、碳水化合物相互作用并沉淀。单宁是水果和蔬菜中涩味的主要物质,分子内羟基数在1~5个范围内,涩味随羟基数量增加而增加,但超过7个羟基,由于空间位阻效应(steric hindrance)开始抵消氢键的强度,涩味变弱[2]。
单宁可分为水解单宁和缩合单宁两大类,缩合单宁又称儿茶素单宁或原花青素。水解单宁由酸及酸的衍生物与葡萄糖或多元醇通过酯键形成,易被酸或酶水解为酚酸、糖和多元醇。根据分解产生的酚酸的种类不同,水解单宁又可以分为没食子单宁(棓单宁)和鞣花酸单宁。没食子单宁的水解产物为没食子酸,鞣花单宁的水解产物为六羟基联苯二酸和鞣花酸。水解单宁生物活性较强,在医药、食品、化工等领域应用广泛[3]。
缩合单宁是植物中含量最丰富的多酚类物质,是一类由黄烷-3-醇单元结构通过4→8或4→6 C-C键(也有通过C-O-C键)缩合而形成的寡聚物或多聚物[4],也被称为儿茶素单宁、原花青素、低聚原花青素(oligomeric proanthocyanidin,OPC)。在热醇-酸溶液中能酸解生成花色素。黄烷-3-醇及黄烷-3,4-二醇是缩合单宁的前体,经缩合最终形成缩合单宁。在热酸处理下,黄烷-3-醇不产生花色素,不属于原花色素,但是原花色素的重要前体。由于缩合单宁的结构单元类型、连接方式、聚合度、空间构型多样性,目前只准确测定了部分缩合单宁的化学结构。
单宁广泛分布于水果、蔬菜、坚果等中,主要以膳食多酚的形式被人体摄入,日常饮食中单宁的含量见表1。尽管日常饮食中含有丰富的单宁(每日摄入约0.1~0.5 g),但由于其聚合性质及其高级结构的复杂性,目前对单宁的化学结构及其生物活性的研究仍然较少[5]。
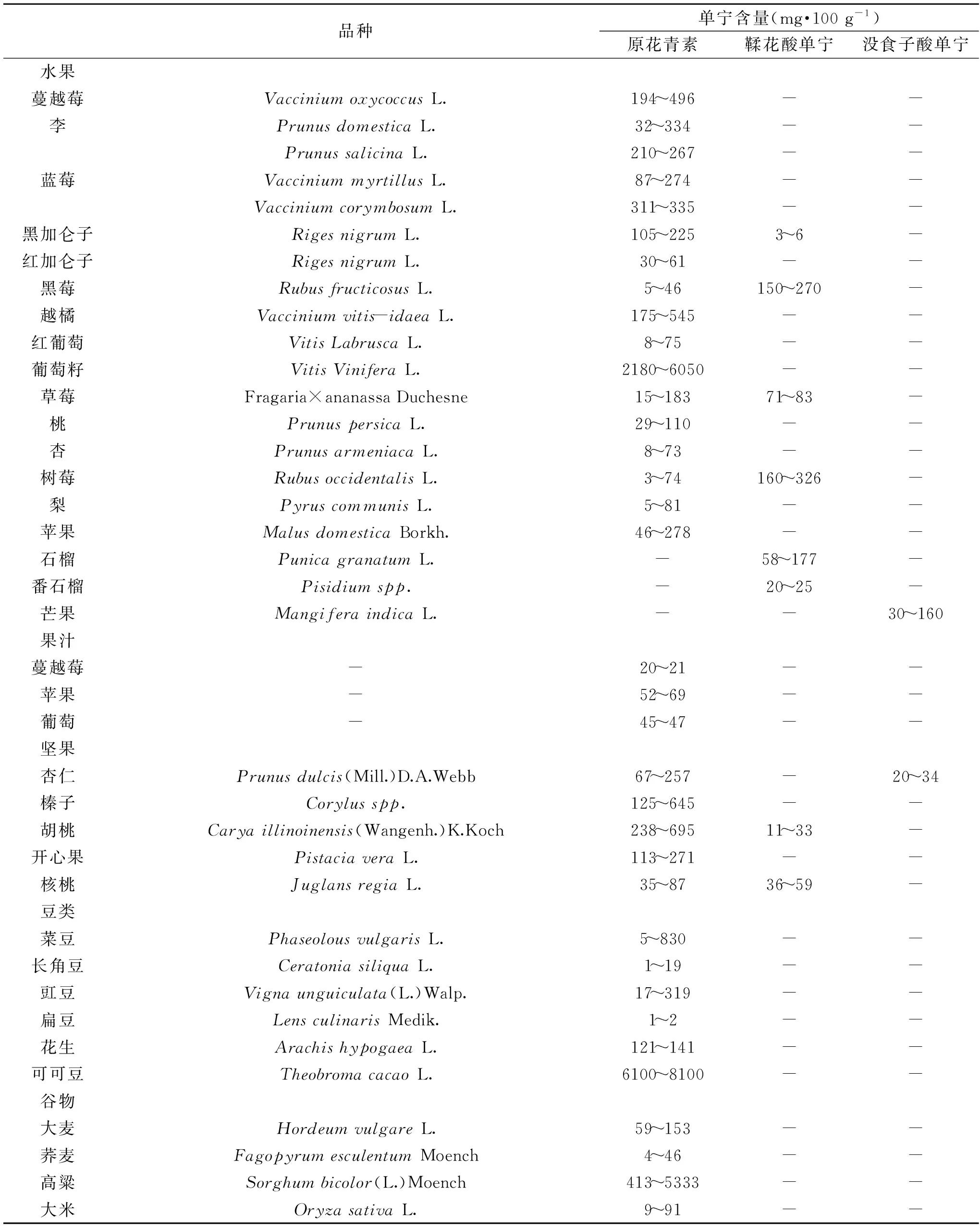
表1 不同食品中单宁的含量Table 1 Tannins content in different foodstuffs
近年来的研究发现原花青素单体具有免疫调节、抗炎、抗癌、抗氧化、保护心血管和抗血栓等多种生物活性[6-8],引起了研究者的广泛关注。本文综述了近几年单宁吸收代谢和生物活性的研究结果,为单宁的生物活性研究、开发利用新型药物资源等提供更多的理论依据。
1 单宁的吸收和代谢
简单酚类物质在人体内的吸收、代谢和生物利用率已有相关研究[9],但由于单宁的化学结构复杂,简单酚类的代谢规律并不完全适用于原花青素,其吸收代谢过程仍存在争论。Wiese等[10]的研究表明,高分子量的原花青素并不能被完全吸收,苹果汁中的原花青素有近90%可在回肠中检出,说明原花青素能够进入结肠而并未被吸收。单宁的聚合度会影响其通过肠屏障的吸收,也会影响肠道微生物对单宁的分解(分解为儿茶素和表儿茶素)。一些体内和体外实验研究了聚原花色素的水解机理及其衍生的低聚分子在小肠中的吸收,在血液中能检测到少量的原花青素单体和二聚体,其主要代谢产物为甲基化和葡糖醛酸化衍生物。在结肠中,原花青素可被肠道微生物分解成一系列简单代谢物,如苯戊酸酯、苯乙酸和苯丙酸[11]。然而,这些实验结果及其机理仍然存在争议。
1.1 体外实验
体外实验结果表明,可可原花青素可在模拟人胃环境下水解成表儿茶素单体或二聚体。但有其他研究表明,可可原花青素在胃中并不能完全解聚。这可能由于实验设计的条件提供的模拟胃酸环境不同,结果也有所不同[12-13]。在消化过程中,小分子的原花青素(如二聚和三聚体)更容易被吸收,高分子量的原花青素可与蛋白质、淀粉和消化酶形成复合物,形成不易消化的复合物,这与单宁的聚合程度有关。低聚原花青素的吸收代谢已有相关综述报道,原花青素A型的二聚体、三聚体和四聚体可穿过Caco-2细胞单层膜,转运率分别为0.6%、0.4%和0.2%,这表明饮食摄入的低聚原花青素在体外可被消化细胞吸收。此外,(+)-儿茶素和原花青素二聚体和三聚体的渗透系数相似,而高聚合度的原花青素的转运率比其低约10倍[11]。Engemann等[14]使用猪盲肠模型模拟肠道微生物群对原花青素A2和肉桂鞣质B1的降解作用,发现约80%的原花青素A2和约40%的肉桂鞣质B1被降解为羟基化的多酚类化合物。水解单宁经几种细菌酶代谢为没食子酸、连苯三酚、间苯三酚,最后可代谢为乙酸酯和丁酸酯[15]。
综上,体外实验的结果表明膳食摄入的原花色素大多可以经过消化系统,并可能到达结肠部位,通过微生物的分解吸收,产生苯乙酸、苯丙酸和苯丁酸等代谢产物[15]。Déprez[16]利用14C标记检测代谢产物,发现代谢产物浓度较理论值低。这可能是由于原花色素与肠细胞中各种其他分子以及与结肠腔中的纤维强烈结合所导致。目前关于原花色素代谢菌菌种的研究较少。
1.2 动物实验
低聚原花青素可在大鼠小肠中被水解成表儿茶素[12]。用荔枝多酚提取物灌胃大鼠,在大鼠血液中可检测到原花青素原型,尿液中可以检测到微量原花青素二聚体和三聚体[17]。这些结果表明,只有原花青素的三聚体和二聚体可以在体吸收,这可能是由于其渗透系数与甘露糖醇相似,也可能由于肠内腔对聚合物的生物利用度有限[18]。用柳树原花青素灌胃大鼠,在尿液中检测到16种代谢物,主要包括苯乙酸、苯丙酸和苯基丁酸等。其含量随聚合度的上升而显著减少:儿茶素单体>二聚体>三聚物>多聚体[19]。
Goodrich等[20]研究了大鼠鼻饲法给予葡萄籽原花青素后的代谢产物,检测到了14种原花青素单体化合物和24种微生物代谢物。组织学检测结果显示,原花青素单体和二聚体在盲肠和结肠中聚集,原花青素在盲肠及其附近区域先出现,大约3 h后达到最大浓度,代谢产物达到最大浓度3~18 h后,出现在其他的末梢区域。Bittner等[21]研究了猪口服给予原花青素B4后的吸收和代谢,结果表明原花青素B4可作为完整分子被吸收,部分由尿液排泄,在体内可被降解为单分子的儿茶素和表儿茶素,然后进一步代谢为甲基化和葡萄糖醛酸化物。
综上,肠道微生物只能分解原花青素单体和二聚体,随原花青素的分子量增大,其分解作用降低。因此聚合度较低的原花青素能够被机体所吸收并发挥其抗氧化活性,聚合度较高的原花青素由于分子量太大而无法被吸收,使其体内抗氧化活性减弱。
2 单宁的生物活性
单宁的生物活性已得到广泛的关注,国内外已有相关文献[22-25]报道。单宁可防止多种慢性疾病的发生,目前认为单宁的生物活性主要来源于黄烷-3-醇及其衍生物[24,26-27]。单宁的生物活性降低主要取决于两方面,一方面是高分子量化合物的生物利用率低,降低了其生物活性;另一方面是单宁与其他物质相结合影响其活性。其结合性质可在胃肠道中产生局部作用(如与蛋白质结合),而其代谢产物(二聚体和三聚体)可被人体吸收,产生全身效应[8-9,18,28-29]。单宁具有抗氧化、抗菌、抗病毒、抗癌、抗炎、抗过敏和血管扩张等作用[6,11,24,26-27,32]。单宁的主要生物活性及其作用机理见表2。

表2 单宁生物活性研究进展Table 2 Advances in research on biological activity of tannins
2.1 抗氧化、清除自由基
单宁是良好的自由基清除剂和脂质过氧化抑制剂,其作用机制主要是其结构中的酚羟基在体内释放H+,竞争性地与自由基结合,从而保护脂质不被氧化,阻断自由基链式反应。体外实验还证明,原花青素可以抑制脂质过氧化酶和脂氧合酶活性[51],从而防止脂质的氧化。水解单宁和原花青素的抗氧化活性取决于其化学结构,随聚合度的增加,抗氧化活性随之增加[8]。蔓越莓原花青素的抗氧化活性是抗坏血酸的10倍,是α-生育酚的37倍[52],一些体外实验和动物实验也证明了相似的结论[6,8,30-31]。
Fushimi[53]比较了青熟和完熟柿(DiospyroskakiThunb.)果肉提取物对大鼠的抗氧化作用,结果表明青熟柿组大鼠血浆中磷脂氢过氧化物(膜脂质过氧化产物)水平显著低于对照组,而完熟柿组与对照组比较无差异。这主要是由于青柿中较多的可溶性单宁提高了果实的抗氧化活性。另一项研究表明,柿果肉和花生皮中的几种原花色素型二聚体在小鼠体内均具有较高的抗氧化活性,B型二聚体在水相中有较高的抗氧化能力,而A型和B型二聚体在组织或脂溶性体系中抗氧化能力相近[54]。
Zhou[55]测定了红景天(RhodiolaroseaL.)中的低聚原花青素对小鼠血清、心脏、肝脏和脑组织中自由基的清除活性。实验表明这些低聚原花青素可以降低细胞的丙二醛含量,增强SOD和谷胱甘肽过氧化物酶(GPx)活性。Li等[56]通过证明原花青素可提高总抗氧化能力,提高SOD、GSH、GPx和CAT活性,降低MDA水平,从而减少氧化损伤。
2.2 抗肿瘤活性
原花青素的抗肿瘤活性主要是通过调节抗氧化、抗炎、调节信号分子的表达,以及促进肿瘤细胞的凋亡、阻滞细胞周期等作用。Gollucke[33]的研究表明葡萄籽原花青素通过调控Bax/Bcl-2基因的表达并激活半胱氨酸蛋白酶-3,诱导小鼠皮肤表皮细胞JB6 C141进行p53依赖性凋亡。此外,葡萄籽原花青素能够抑制人表皮角化细胞中MAPK蛋白(mitogen-activated protein kinase,MAPK)的磷酸化并调控NF-κB的表达。葡萄籽原花青素可以抑制前列腺癌激素不敏感细胞DU145和前列腺癌激素敏感细胞LNCaP的生长并抑制其中基质金属蛋白酶MMP-2和MMP-9的表达,葡萄籽原花青素能够通过增强抗组蛋白乙酰转移酶的活性来调节激素受体介导的转录[57],基质金属蛋白酶(matrix metalloproteinases,MMPs)参与肿瘤组织的生长和代谢。Chung等[58]也发现葡萄籽原花青素可以抑制胰腺癌细胞中基质金属蛋白酶的活性。葡萄籽原花青素可以抑制小鼠乳腺癌4T1细胞、结肠直肠癌HT29细胞、LoVo细胞系的细胞活性、增殖,促进其凋亡[8]。Prasad[23]发现葡萄籽原花青素通过干扰细胞周期进程的正常调节,诱导Cip1/p21和Kip1/p27蛋白水平上升,以及细胞周期蛋白D1、D2 E和细胞周期蛋白依赖性激酶CDK2、CDK4、CDK6,导致细胞生长周期停滞于G0/G1期。
除葡萄籽原花青素外,葡萄和松树皮等富含原花青素的组织同样可以有效抑制HT29人结肠直肠癌细胞增殖,诱导细胞凋亡和G2期细胞周期阻滞[33]。体外实验中,浆果原花青素提取物对人口腔表皮样癌细胞(KB)、人舌鳞癌细胞(CAL-27)、人乳腺癌细胞系(MCF-7)、人结肠癌HT-29、116细胞和前列腺淋巴结癌细胞(LNCaP)有抗增殖和促进凋亡的作用[6]。
2.3 抗菌性
单宁可抑制胞外酶活性、与微生物生长所需的营养物质相结合(如络合金属离子)、使原生质中的蛋白质沉淀变性、破坏细菌外膜通透性或直接作用于微生物代谢影响其生长。此外,单宁还可以干扰微生物细胞壁多肽与巯基反应,使膜蛋白功能丧失[15]。
浆果中的原花青素可以抑制细菌的粘附性,蔓越莓原花青素可以抑制粘附于尿路上皮的大肠杆菌的生长。原花青素的分子结构会影响其抗粘附性,蔓越莓原花青素是由一系列儿茶素寡聚体和A-型连接形成的聚合物,在结构上与葡萄、苹果等其他来源的原花青素不同,是唯一能够抑制细菌在人体粘附的原花青素。动物实验结果也证明蔓越莓原花青素及其代谢物能够抑制细菌在尿路上皮的粘附[59]。
云莓(RubuschamaemorusL.)、树莓、草莓多酚提取物含有丰富的鞣花酸单宁,对空肠弯曲杆菌(Campylobacterjejuni)和白色念珠菌(Candidaalbicans)具有很好的抑制效果。覆盆子和蓝莓多酚提取物对革兰氏阳性菌(单胞李斯特菌、金黄色葡萄球菌、枯草芽孢杆菌和粪肠球菌)和革兰氏阴性菌(费氏柠檬酸杆菌、大肠杆菌、铜绿假单胞菌和鼠伤寒沙门氏菌)都有抑制活性[6],幽门螺杆菌和蜡状芽孢杆菌对浆果原花青素最敏感,而发酵酵母对其具有抗性[60]。林生假榆桔(PteleopsishylodendronMildbr.)多酚提取物富含鞣花酸单宁,能够抑制肺炎克雷伯杆菌、蜡样芽孢杆菌、大肠杆菌和伤寒沙门氏菌活性[15]。石榴皮多酚提取物能抑制金黄色葡萄球菌、伤寒沙门氏菌、单胞李斯特菌和大肠杆菌活性。鞣花酸单宁能够抑制金黄色葡萄球菌(包括甲氧西林金黄色葡萄球菌)、大肠杆菌、白色念珠菌荚膜的形成[61]。安石榴甙、安石榴林具有抗真菌活性,可以抑制白念珠菌、新型隐球菌、烟曲霉活性[15]。
单宁的抗病毒活性与其化学结构有关,化学性质越活跃,其细胞毒性越强。单宁可以通过与病毒包膜组分相结合从而影响其活性。单宁可干扰单纯疱疹病毒(HSV)、艾滋病毒(HIV)对营养物质的吸收及其产生的细胞病变效应[60]。鞣花酸单宁和几种原花青素还是逆转录酶抑制剂[62],可以抑制乙型肝炎病毒感染细胞中E抗原(HBeAg)的分泌[15]。
3 结语
单宁为多酚类化合物,在植物中分布广泛,具有多种生物活性,能够清除自由基、抗菌、抑制肿瘤、保护心血管、预防糖尿病。目前对单宁的研究正日趋成熟,其应用也愈加广泛,但有些方面的研究仍有待提升。首先,生物利用率是单宁在体内产生生物活性的先决条件,单宁在体内的吸收机制、其影响因素尚未完全明确,这对开发和利用单宁类化合物具有重要的意义。此外,单宁的化学结构复杂,分子量大小、结构单元类型、连接方式、聚合度、空间构型等对其生物活性的影响尚不明确,需要大量的实验数据和总结考证。单宁及其代谢产物产生多种生物活性的机制尚有待进一步研究,建议进一步确定其体内代谢中潜在的活性成分。最后,单宁的细胞毒性及机理尚未明确。因此,对于单宁的研究仍有大量工作需要做。
