P2X受体在骨癌痛发生和维持中的作用及其机制探讨*
2018-08-20施任怡蔡杨乾杜俊英房军帆
施任怡 蔡杨乾 杜俊英 房军帆 梁 宜
(浙江中医药大学第三临床医学院针灸神经生物学实验室,杭州 310053)
近年来,恶性肿瘤的发生率呈逐年上升的趋势,严重危及人类生命。然而,随着癌症诊疗技术的进步,癌症病人的生存时间明显延长, 癌痛逐渐成为影响癌症病人生活质量的主要原因之一。据统计约有40%早中期癌症病人和90%晚期癌症病人都在经历中度或重度疼痛;且有高达70%的癌症痛病人疼痛并未得到充分的缓解[1]。骨癌痛是晚期癌症病人最常见的疼痛,也是肿瘤发生骨转移时最常见的症状之一。目前的治疗方法还不能完全有效控制骨癌痛,因此针对骨癌痛的预防和控制是癌症治疗的一大难题,仍需进一步深入研究骨癌痛的发生发展机制。近年研究提示P2X受体参与了肿瘤发生和发展[2]。P2X受体介导的嘌呤信号在癌症发生、肿瘤生长及其诱发痛觉敏化中起着至关重要的作用[3]。P2X受体是一种三磷酸腺苷(adenosin triphosphate,ATP)的门控离子通道,主要包括七个亚型。已有研究证明,其中P2X3、P2X4、P2X6、P2X7受体参与骨癌痛的产生和维持过程,且存在不同的调制机制[4~6]。故本文将围绕P2X受体在骨癌痛的产生和维持中的作用及其可能机制进行阐述,为今后治疗骨癌痛提供新的研究思路和方法。
1.P2X受体的结构和分布特点
(1) P2X受体结构特点:到目前为止,已有七个P2X (P2Xl-7)被克隆,并被认定为P2X受体家族成员。P2X受体各亚型具有相同的一般结构:由胞内的N末端和C末端,两个跨膜区和一个细胞外环组成[7]。其中细胞外环包含有10个保守的半胱氨酸残基,以二硫键互相连接,细胞外环上含有ATP结合位点和相应的激动剂、拮抗剂和调节剂的作用位点。P2X受体七个亚型的氨基酸序列之间有35%~48%的同源性,其氨基酸残基数从379到595个不等[8]。据既往研究表明P2X受体氨基端较羧基端短,羧基末端长度不相同,最短的是P2X6,有25个残基,P2X7则拥有240个残基[9]。P2X受体可形成同聚体或异聚体的ATP门控非选择性阳离子通道,被ATP激活后可以引起Na+、Ca2+内流和K+外流,但对Ca2+的通透性最大[10]。目前认为P2X受体属寡聚蛋白类,其受体亚基数量从3个到6个不等[11,12]。
(2)P2X受体分布特点:P2X受体不同亚型在中枢神经系统和外周神经系统均有分布,并且无论在中枢或是外周神经系统中,P2X受体主要表达在神经元细胞和神经胶质细胞。由于P2X受体对伤害性、炎症性和病理性疼痛的传导都是必不可少的,因此在参与疼痛转导的感觉神经节、背根神经节(dorsal root ganglion, DRG)和脊髓后角神经元均能检测到P2X受体(见表1)。
2.P2X受体介导骨癌痛
有研究报道,ATP及其作用的P2X受体广泛分布于与伤害性信息传递有关的外周或中枢神经细胞中,它们与伤害性刺激的感受及传递密切相关。越来越多的研究证实,ATP介导的嘌呤信号在癌症疼痛的发生中起着至关重要的作用[22]。由于在肿瘤微环境中,ATP的释放不仅能够促进癌细胞的转移和浸润,而且能促进神经细胞和免疫细胞刺激物的产生[23]。有学者发现,在MRMT-1乳腺癌细胞致骨癌痛模型中,ATP在神经元和神经胶质细胞(主要是脊髓星形胶质细胞)集中释放,从而激活P2X受体,介导大鼠疼痛相关行为的产生[24]。综上所述,进一步说明了P2X受体在骨肿瘤生长、癌细胞转移和浸润、疼痛因子的释放,最终介导骨癌痛产生这一过程中起着重要的作用。现有的研究报道显示,参与骨癌痛发生和发展的P2X受体主要有P2X3及其异聚体P2X2/3、P2X4、P2X6、P2X7受体。
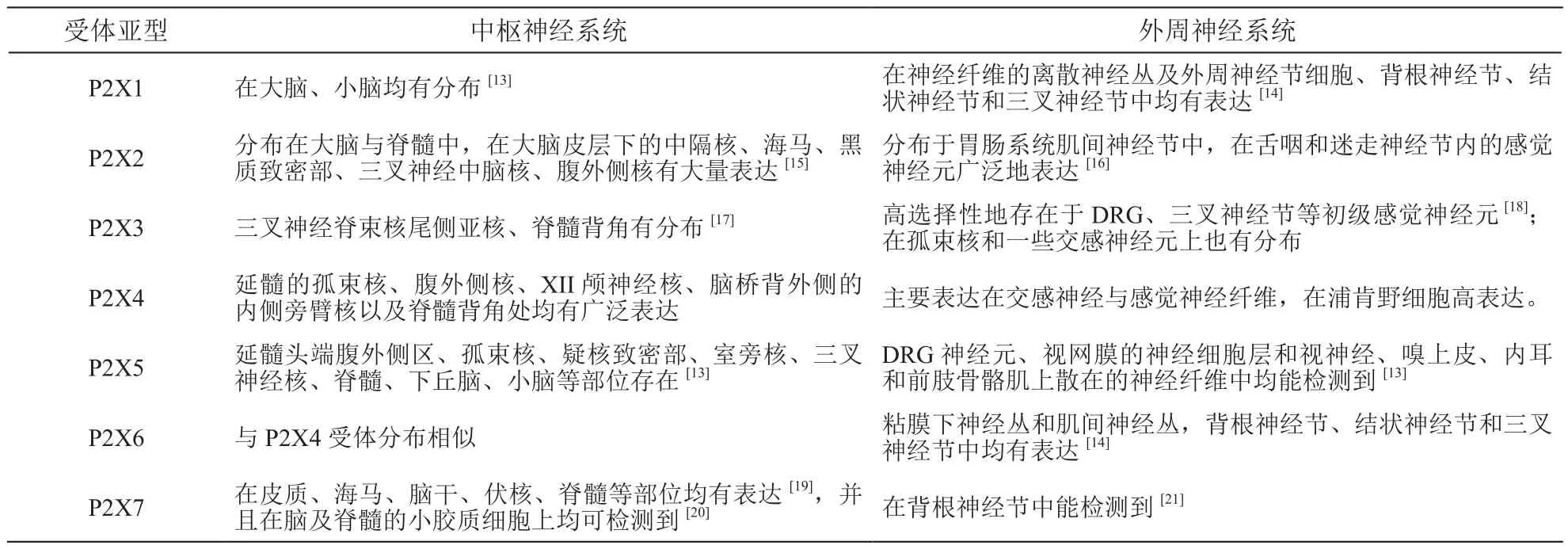
表1 P2X受体各亚型在中枢和外周神经系统的分布
(1)P2X3和P2X2/3受体:在P2X受体中,与疼痛关系最为密切的是P2X3受体,ATP主要通过同聚P2X3受体和异聚P2X2/3受体参与痛觉及伤害性信息传递,并参与骨癌痛的感知。研究发现,骨癌痛模型中,MRMT-1癌细胞释放出ATP引起骨传入神经高表达P2X3和P2X2/3受体,进一步促进脊髓水平的中枢敏化引起骨癌痛的形成[25]。也有学者发现骨癌痛后DRG神经元中P2X3受体mRNA表达升高,在接种癌细胞后的第14、21天显著增多[4]。Gilchrist等[26]报道,骨癌痛小鼠趾部皮肤降钙素基因相关肽(calcitonin generelated peptide,CGRP)免疫阳性的C纤维上P2X3受体表达上调,增加C纤维对ATP诱发疼痛的反应性,促进骨癌痛的形成。而使用P2X3和P2X2/3受体选择性拮抗剂A-317491可以减轻大鼠骨癌痛模型的炎症性和神经性疼痛[27]。也有文献报道,鞘内持续注射P2X3受体反义寡核苷酸2 d后,可明显减轻大鼠足底注射α,β-meATP引起的机械痛觉过敏,同时背根神经节的P2X3受体蛋白水平显著降低。口服或者鞘内注射高选择性P2X3和P2X2/3受体拮抗剂AF-353可减轻大鼠骨肿瘤引起的疼痛,但对骨肿瘤引起的骨质破坏无作用[28]。这些研究提示P2X3受体和异聚P2X2/3受体在骨癌痛的伤害性信息传递中起重要作用。
(2)P2X4受体:有研究表明,P2X4受体参与了神经病理性疼痛和炎性痛的产生和维持,同时脊髓P2X4受体特异性存在于小胶质细胞表面,参与了大鼠骨癌痛的产生[29]。任春光[30]等报道,骨癌痛模型中,脊髓背角小胶质细胞中P2X4受体阳性细胞数表达在术后第6~18 d呈进行性上升趋势,鞘内注射TNP-ATP(P2X1-4受体抑制剂)后,骨癌痛大鼠的机械性触诱发痛降低。P2X4受体基因敲减,同样可有效对抗骨癌痛导致的痛觉过敏。这些研究结果提示,脊髓小胶质细胞P2X4受体参与了大鼠胫骨癌痛的产生和维持。
(3)P2X7受体:以往研究证明,P2X7受体激活不仅能促进疼痛因子释放,而且能通过肿瘤的生长、浸润和血管化最终导致骨癌痛的产生。Huang等[31]发现,胫骨注射Walker 256癌细胞后的第3~14 d,骨癌痛大鼠延髓头端腹内侧(rostroventral medulla,RVM)中小胶质细胞和星形胶质细胞上的P2X7 mRNA和蛋白水平上调;RVM中注射P2X7受体拮抗剂BBG或P2X7受体基因si RNA可以抑制RVM中P2X7受体表达,起到镇痛作用。有文献报道,腹腔注射P2X7受体特异性拮抗剂A-839977可缓解骨癌痛大鼠疼痛行为[32]。
但是,骨癌痛作为一种机制复杂而独特的慢性痛,P2X7受体在其中的作用研究也有相反的结论。Hansen等[33]人研究发现,P2X7受体缺陷的BALB/cJ小鼠对骨癌痛易感,而且皮下注射选择性的P2X7受体拮抗剂A438079 (300 μmol/kg)后,均不能减轻骨癌痛小鼠的痛行为。有学者认为,在小鼠骨癌痛模型中,P2X7受体不参与骨癌诱发痛觉敏化的发生和发展。考虑可能与大、小鼠骨癌痛条件下胶质细胞不同的激活情况有关。
值得一提的是,经Park等[34]人研究发现,在骨癌痛小鼠脊髓背角中P2X6受体表达显著上调,提示P2X6受体可能介入小鼠骨癌痛的脊髓机制。综上所述,P2X3、P2X4、P2X6受体与骨癌痛的产生和维持最为密切,而P2X1和P2X5受体与骨癌痛的研究目前未见相关报道,但不排除两者存在关联的可能性,有待更系统性的研究。
3.P2X受体参与骨癌痛的机制研究
目前对于P2X受体介导骨癌痛的机制研究主要涉及到P2X3、P2X4、P2X7受体,其他类型受体由于缺乏相关研究证据,暂时还不清楚其参与骨癌痛的内在机制。
(1)P2X3受体参与骨癌痛的可能机制:目前P2X3受体参与骨癌痛的作用机制主要被认为与钙敏感蛋白(visinin-like protein 1, VILIP -1)、核转录因 子 (nuclear transcription factor-κB, NF-κB)、 溶 血磷脂酸受体1 (LPA receptor subtype 1, LPAR1)、阿片受体(µ-阿片受体)有关。Liu等[35]研究发现在大鼠骨癌痛模型DRG神经元上VILIP-1被激活后促进其氨基末端与P2X3受体的C-末端直接作用,增加P2X3受体在细胞膜上的表达与功能;过表达VILIP-1增加正常大鼠功能性P2X3受体的表达。亦有文献报道,在骨癌痛大鼠的脊髓中NF-κB可与P2X3受体相互作用实现对P2X3受体表达的调制[36]。鞘内注射 NF-κB p65 shRNA 慢病毒或者 PDTC(NF-κB抑制剂)可以逆转P2X3受体高表达,从而降低骨癌痛大鼠机械痛。近期有相关研究表明,LPAR1通过激活其下游Rho/ROCK信号,引起细胞膜上相关Ca2+通道活性增强或敏感性增高,进而促使Ca2+内流增大,参与P2X3受体介导的骨癌痛;VPC32183(LPAR1抑制剂)可以阻断α,β-meATP诱导的自发痛行为;并且鞘内注射BoTXC3(Rho抑制剂)、Y27632(ROCK抑制剂)可以瞬时翻转P2X3受体介导骨癌痛大鼠机械痛下降和自发痛行为[37]。Chizhmakov等[38]人报道,P2X3受体可能通过降低阿片受体(µ-阿片受体)的敏感性促进癌性疼痛的产生。而相关的研究表明骨癌痛模型中,µ-阿片受体表达减少,功能减弱是通过磷脂酶C (phospholipase C, PLC)的活化实现的。由上述讨论可见,P2X3受体介导骨癌痛的具体机制目前仅局限于相关受体、蛋白分子和核转录因子与P2X3受体的相互作用,而其他物质是否参与P2X3受体介导的骨癌痛有待进一步证实。
(2)P2X4受体参与骨癌痛的可能机制:目前研究结果显示,Toll样受体4 (Toll-Like Receptor-4,TLR4)的活化、脑源性神经营养因子(brain-derived neurotrophic factor, BDNF)大量释放参与脊髓背角的小胶质细胞中P2X4受体介导骨癌痛的产生和维持。最新的研究报道,脊髓背角的小胶质细胞中P2X4受体的激活受TLR4的调制,但TLR4并不直接作用于P2X4受体,而是通过作用于p38 MAPK信号转导通路,从而激活P2X4受体;使用TLR4 siRNA能降低p-p38 MAPK蛋白表达,阻断P2X4受体以及痛觉过敏的产生。同样的,鞘内注射SB203580(p38 MAPK抑制剂)可以有效缓解P2X4受体活化诱导的机械痛觉过敏[5]。另外,研究者还发现,脊髓背角小胶质细胞上P2X4受体被激活后,促使BDNF大量释放,引起感觉超敏导致骨癌痛;P2X4R siRNA可以抑制BDNF,缓解骨癌痛大鼠机械痛[39]。后期的进一步研究表明BDNF的释放,能促使BDNF与其特异性受体结合,进一步调控神经元N-甲基-D-天冬氨酸受体(N-methyl-D-aspartic acid receptor,NMDA)功能,从而实现骨癌痛的产生[40]。此外,结合前期关于P2X4受体参与神经性病理性疼痛的机制报道,BDNF的大量释放,可以抑制脊髓背角神经元胞内氯化物的产生[41]。因此笔者认为BDNF作为重要中介物质,在P2X4受体介导的骨癌痛中发挥重要作用。除此之外,也有文献报道,纤维连接素和核苷酸结合寡聚蛋白2 (nucleotide-binding oligomerization domain containing2,NOD2)可以引起P2X4受体mRNA表达增加,表明纤维连接素和NOD2可能也参与了P2X4受体的上调[42,43]。综上所述,脊髓背角小胶质细胞中P2X4受体经证实参与骨癌痛的产生和维持,但是笔者认为在中枢和外周神经系统中的其他部位的P2X4受体可能也参与了骨癌痛的产生及维持且具有不同的作用机制,还需要进一步研究和确证。
(3)P2X7受体参与骨癌痛的可能机制:从文献资料的阐述中了解到,P2X7受体参与骨癌痛主要是通过骨质破坏、免疫细胞释放的各种细胞因子最终引起初级感觉神经元兴奋性的改变,从而介导骨癌痛的产生。骨质破坏主要是由肿瘤细胞的浸润促进了破骨细胞的生成,抑制成骨细胞分化,打破了骨溶解与骨再生的平衡所致。而目前研究表明,P2X7受体在成骨细胞和破骨细胞上均有分布,且其参与骨癌痛的作用机制也有所不同:成骨细胞上P2X7受体通过激活PI3K/Akt、NFATc1、ROCK和VEGF等信号通路促进早期肿瘤的发展和成骨细胞的损伤,分别阻断其信号通路能有效抑制成骨细胞的损伤,促进骨再生,减轻骨质破坏[44~46];破骨细胞上的P2X7受体能上调RANKL(转录因子NFκB配体)的表达,增强骨吸收和溶骨性病变[47];除此之外,P2X7受体在免疫细胞上通过在高浓度的ATP激活作用下,释放大量的促炎症因子,如IL-1β、IL-6和IL-18等,这些细胞因子均可兴奋初级感觉神经元,介导痛觉过敏或痛觉超敏[48]。P2X7受体在炎性痛和神经病理性疼痛中的作用已有大量深入的研究,而在骨癌痛中的作用机制研究相对较少,以期更深入的研究。
4.结语
目前,针对骨癌疼痛机制的研究是肿瘤相关研究中最迫切需要解决的问题之一。大量文献研究显示P2X受体可能是介导肿瘤生长和癌痛的共同作用靶点,拮抗该受体既可以起到抑制肿瘤生长浸润的作用,又可以有效地缓解骨癌痛。但是,癌性疼痛不仅仅是神经病理性疼痛、炎性疼痛两者的结合,因此对P2X受体激活在骨癌痛调制机制中的作用还需做更加全面的探究;另外,P2X7受体在骨癌痛中的作用目前尚处于初始阶段,相关的研究报道相对较少,对于P2X7受体在其中的作用也较有争议,需要更多相关的研究和探索来进一步阐明。
