淮河青虾线粒体细胞色素氧化酶I序列的研究
2018-07-13肖明松鲍方印
肖明松, 鲍方印, 康 健
(安徽科技学院 生命与健康科学学院,安徽 凤阳 233100)
日本沼虾(Macmbrachiumnipponensis) 俗称青虾,又名河虾、沼虾,为节肢动物门甲壳纲十足目游泳虾亚目长臂虾科沼虾属,广泛分布于我国各地的江河、湖泊、水库、池塘及沟渠中,是我国淡水天然虾类中的一个重要种群,也是我国淡水养殖的重要虾类之一。其市场销售量经久不衰,深受消费者青睐[1-3]。青虾具有适应力强、繁殖力高等特点,与其他甲壳类水产品相比[4-5],如河蟹,青虾具有营养丰富,肉质细嫩鲜美[6],易消化,食用方便且蛋白质含量高等优点。另外青虾体内含有丰富的镁离子,可以有效预防心脑血管疾病的发生,并且青虾具有增强记忆力的功效[7]。近年来,由于农村地区工业化的发展,很多地区建立起了许多化工厂,污水排放,生活垃圾等严重影响了青虾的生长环境[8]。在日本沼虾养殖过程中由于长期捕大留小、忽视良种选育以及过度捕捞野生资源,使得日本沼虾野生资源逐渐衰竭,养殖日本沼虾出现了个体小型化、性早熟以及生长速率慢等种质退化现象[4,9]。目前,其研究大多数集中在其分类[1-3]、生活习性[6]、繁殖和养殖等方面的研究[10-11],而关于其种群遗传和分子进化方面的研究相对较少[4,12-13],尤其关于淮河流域青虾的分子生物学研究更少。因此,研究淮河流域青虾细胞色素氧化酶I基因序列为后期青虾的种质资源保护以及青虾养虾业的可持续发展提供理论参考。
1 材料与方法
1.1 材料
青虾取自淮河蚌埠段,虾龄10个月左右,全长为8~12 cm,体重15~20 g,将受试材料用95%的乙醇固定后运回实验室,置于-20 ℃冰箱冷冻保存以备用。
1.2 方法
1.2.1模板DNA 的制备采用苯酚-氯仿法提取淮河青虾背部肌肉组织的基因组DNA,并用紫外分光光度计测量其浓度和纯度,同时用琼脂糖凝胶电泳检测基因组DNA 的完整性并估测分子量。基因组DNA样品置于-20 ℃冰箱保存备用[14]。
1.2.2PCR反应用于青虾线粒体细胞色素氧化酶I序列扩增的两条引物为: LCO1490 (5- GGT CAA CAA ATC ATA AAG ATA TTG G -3)和HCO2198 (5- TAA ACT TCA GGG TGA CCA AAA AAT CA-3)[15]由生工生物工程(上海)股份有限公司合成。PCR 反应体系为30 μL,其中含10×buffer反应缓冲液3.0 μL,3 μL dNTP (2.5 mmol/L),引物LCO1490和HCO2198各1.2 μL,0. 3 μL Taq 酶,模板DNA 1.0 μL,加灭菌蒸馏水至30.0 μL。PCR反应条件为:94 ℃预变性4 min;94 ℃变性30 s,52 ℃退火45 s,72 ℃延伸1 min,共38个循环;最后72 ℃延伸10 min,4 ℃保存。
1.2.3PCR产物检测、测定和分析 用1.0%的琼脂糖凝胶电泳PCR 产物,电泳结果在化学发光凝胶成像系统(ChemiDoc XRS)中拍照,保存。同时,将PCR产物送往生工生物工程(上海)股份有限公司进行测序,将测序结果分别在NCBI上进行BLAST搜索验证。用Clustal X 2.11软件[16]进行碱基比对,辅以人工校对。将获得的序列与GenBank 中已有的青虾线粒体DNA全序列(HQ 830201)进行同源性比较。最后,利用MEGA 4.1 软件包分析序列特征、碱基组成等[17],并计算碱基偏倚性。
2 结果与分析
2.1 青虾基因组DNA电泳检测
青虾总基因组DNA凝胶电泳检测(图1)。由图1可知,青虾总基因组DNA条带清晰、整齐、较完整、有拖尾,说明DNA片断有降解,含有一定杂质。目的片断大于2000 bp,说明提取的总基因DNA分子量较大。

图1 青虾总DNA琼脂糖凝胶电泳
2.2 青虾线粒体DNA Cyt b基因全序列PCR扩增
PCR扩增产物的琼脂糖凝胶电泳检测(图2)。由图2可知,引物LCO1490和 HCO2198在青虾不同肌肉组织中均能扩增到线粒体细胞色素氧化酶I序列序列条带,细胞色素氧化酶I序列条带电泳图谱条带清晰、明亮,线粒体细胞色素氧化酶I的片段大于1000 bp。这可说明苯酚-氯仿法提取的青虾肌肉组织DNA 质量较高,能够用于后期的群体遗传学研究。

图2 青虾细胞色素氧化酶I的PCR产物电泳检测
2.3 青虾线粒体细胞色素氧化酶I全序列

图3 青虾线粒体细胞色素氧化酶I全序列
青虾线粒体细胞色素氧化酶I全序列长度为1 535 bp,编码蛋白含511个氨基酸。其中A、T、C和G 4种碱基分别为459个、471个、345个和260个,它的起始密码子为ACG,终止密码子为TA。青虾线粒体细胞色素氧化酶I基因序列(图3)。
2.4 青虾线粒体细胞色素氧化酶I的密码子碱基使用
由表1可知,青虾细胞色素氧化酶I序列平均碱基组成 A、T、C及G分别为 30.0%、30.6%、22.5%及16.9%;4种碱基在密码子的第1位上使用较为均衡;碱基T在密码子第2位上的使用率比较高(40.6%),而碱基G的使用比率则降低(16.0%);碱基A在密码子第3位上的使用比率最高(45.8%),碱基G的使用比率最低,仅为5.7%。A+T的碱基百分比总体上大于C+G,平均值分别是60.6%和39.4%。
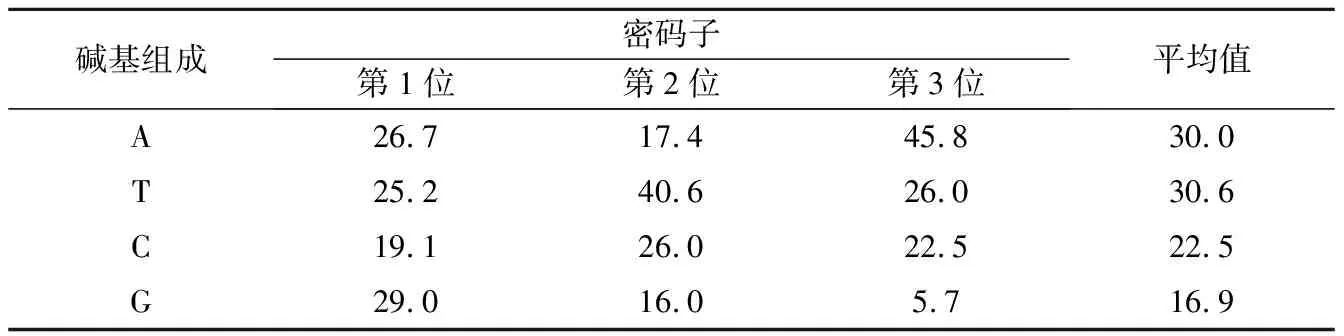
表1 青虾细胞色素氧化酶I的密码子碱基使用情况
3 结论与讨论
青虾线粒体细胞色素氧化酶I(COI)基因序列与其它沼虾的细胞色素氧化酶I基因序列相比,具有较高的同源性。青虾线粒体细胞色素氧化酶I基因全序列为1 535 bp,含起始密码子ACG和终止密码TA。其中,细胞色素氧化酶I基因的终止密码子为不完全的终止密码子TA,但通过转录加工过程中的多聚腺苷酸化,从而形成完整的TAA终止密码子[13]。邵爱华等研究硬骨鱼亚纲1315 种鱼类COI 由1545~1593 个碱基组成,编码514~530 个氨基酸。COI 基因序列中没有发现任何碱基的缺失或插入。15 种鱼类COI 基因的起始密码均为GTG,与其他基因的起始密码子不同[18]。根据检测结果显示,A、T、C、G四种碱基分别含有数目459、471、345、260,占总序列碱基的比例为30.0%、30.6%、22.5%、16.9%,A、T含量明显较多,且A+T的含量(60.6%)明显高于C+G的含量(39.4%)。这一特征和已报道3种沼虾类[19]和中华绒螯蟹线粒体细胞色素氧化酶I基因序列的碱基分布相似[20]。经比较发现两种甲壳纲动物有一个共同遗传分子学的特征:它们细胞色素氧化酶I基因序列的A+T的含量都明显高于C+G的含量,且G含量都表现最低。由于物种的不同,它们的COI序列基因长度存在明显的不同,各碱基出现的比例也不相同。青虾线粒体细胞色素氧化酶I基因的密码子碱基使用存在偏倚性,如碱基A在密码子第3位含量为45.8% ,而碱基G的含量仅为5.7%,密码子第3位反G碱基偏倚分布在其他物种中也被发现,而在一些进化高级的物种细胞色素氧化酶I基因则发现密码子第3位具有对A和C极强偏倚特征[21],这可能是由于蛋白质编码基因的核苷酸突变在密码子第三位点上受到的自然选择压力较小所致,证明密码子第三位点能够更清晰地表明线粒体基因组核苷酸组成的不均一性[22]。
