Ckip-1对小鼠耳廓软骨发育的影响
2018-07-03刘富伟胡奥张浚睿郝英许珊珊侯燕靳丹赵芝鹤薄斌孔亮
刘富伟 胡奥 张浚睿 郝英 许珊珊 侯燕 靳丹 赵芝鹤 薄斌 孔亮
小耳畸形是仅次于唇腭裂之后最常见的口腔颌面部畸形,发病率为5.18/万,同时也是最易导致患者面容不对称的先天性畸形之一[1-2]。耳廓的支架构成以软骨为主,组织发育来源与颅面骨相似,其中软骨细胞的增殖与分化对软骨组织最终形态及功能发挥起重要作用[3]。酪蛋白激酶2相互作用蛋白1(Casein kinase 2 interacting protein-1,Ckip-1)作为重要的骨负调控因子,研究证实应用其siRNA可以促进大鼠下颌骨牵张区骨形成以及种植体骨结合效果[4-5],为其在颌面部应用打下实验基础。课题组前期研究显示Ckip-1对于颌骨和长骨呈现不同的影响效果[6]。然而Ckip-1对于软骨,尤其是颌面部耳廓软骨发育的作用仍罕见报道。在前期实验中发现 Ckip-1基因敲除(Ckip-1-/-)小鼠呈现小耳畸形,本研究将系统的从动物表型、组织学、细胞学及相关细胞因子检测等方面评估Ckip-1在小鼠耳廓软骨发育中的作用,以期为小耳畸形的病因学提供实验证据。
1 材料与方法
1.1 材料
1.1.1 实验动物 选取清洁级Ckip-1杂合子C57小鼠(军事科学院构建)交配生育后取鼠尾组织,PCR鉴定基因型,选取同窝同性别的基因敲除型(Ckip-1 knockout,Ckip-1-/-) 小鼠及野生型(Wild Type,WT)小鼠做配对实验。取6月龄Ckip-1-/-小鼠作为实验组,WT小鼠为对照组(n=6、同窝同性别、雄性)。
1.1.2 实验试剂与仪器 DMEM/F12培养基(Hy-Clone,美国);100 ml/L新生牛血清(Gibco,美国);10 g/L青霉素/链霉素、引物(GeneRay,上海吉凯基因科技有限公司);噻唑蓝(MTT,Sigma,美国);DMSO(上海生工生物工程股份有限公司);总RNA提取试剂、实时荧光定量PCR试剂盒(RNAiso Plus,宝生物工程大连有限公司);Trizol试剂盒(上海普飞生物技术有限公司);扫描电镜(JEDL,日本);分光光度计(BioTek,美国)。
1.2 方法
1.2.1 动物表型观察 取6月龄Ckip-1-/-和WT小鼠(n=6、同窝同性别、雄性),经1%戊巴比妥钠(50 mg/kg)适度麻醉下,以耳廓基部线为基准,观察并测量小鼠耳廓整体形态及边缘位置。
1.2.2 甲苯胺蓝染色 过量麻醉处死动物,解剖取耳廓组织,4%多聚甲醛固定,常规切片。用0.5%甲苯胺蓝水溶液(甲苯胺蓝0.5 g,蒸馏水100 ml,常温保存)置50~60℃温箱染色30 min,双蒸水洗去多余的染液,无水乙醇脱水,中性树胶封片。倒置显微镜下观察染色情况并拍照记录。
1.2.3 ATDC5细胞系培养 ATDC5细胞置于培养基(DMEM/F12)中,加入10%胎牛血清、50 U/ml青霉素、50 U/ml链霉素。1次/3 d换液,保持细胞的对数生长,培养环境为37℃、5%CO2。
1.2.4 细胞模型准备——慢病毒转染及分组 Ckip-1干涉慢病毒载体信息:hU6-MCS-Ubiquitin-EGFPIRES-puromycin;靶点序列:5'-CCTGAGTGACTATGAGAAG-3'(上海吉凯基因)。将ATDC5细胞制成3×104~5×104个/ml细胞悬液,铺12孔板,每孔1 ml;待细胞达到20%左右密度时在感染增强液Eni.S和polybrene辅助下进行感染,感染后8 h换液,感染后72 h进行荧光拍照。收集细胞抽提RNA及蛋白进行QPCR、Western blot检测。分组为:敲低组(Ckip-1干涉慢病毒感染组)、阴性对照组(阴性慢病毒感染组)、空白对照组(未经慢病毒感染组)。
1.2.5 QPCR检测mRNA表达情况 总RNA提取使用Trizol试剂盒,cDNA合成使用M-MLV试剂盒。采用FTC-3000型实时定量PCR仪检测mRNA表达。应用相应引物进行扩增,以 β-actin为对照,应用2-ΔΔCt法计算基因表达量差异。
1.2.6 扫描电镜检测 标本以2.5%戊二醛固定、脱水、干燥、喷金,SEM下观察细胞形态和细胞连接情况。
1.2.7 MTT 实验 在预设的时间点(1、2、3、4、5、6 d),吸出培养基,加入MTT溶液后孵育4 h。吸去培养基,PBS冲洗,每孔加入150μl二甲基亚砜(DMSO)。每孔取出100μl溶液转移至96孔酶标板,测定570 nm处溶液的吸光值。
1.2.8 细胞克隆形成实验 于直径60 mm培养皿中各接种1 000个细胞。培养3 d后,弃去培养基,PBS清洗,甲醇固定15 min。1%亚甲基蓝染液染色30 min后,去离子水清洗后晾干,计数并统计克隆形成率。
1.3 统计学分析
用SPSS22.0软件对数据珋x±s进行统计分析,组间比较用单因素方差分析,检验水准α=0.05,以P<0.05为差异有统计学意义。
2 结果
2.1 Ckip-1-/-小鼠耳廓表型及组织学分析
Ckip-1-/-小鼠表现为小耳畸形,具体为:①耳廓长径、宽径的同时缩小,整体呈蜷缩状不伸展(图1A、1C)。②甲苯胺蓝染色显示,WT型小鼠耳廓软骨板横断面呈现双层软骨细胞结构,成熟软骨细胞较多,周边散在幼稚软骨细胞(图1D);而Ckip-1-/-小鼠耳廓软骨板横断面呈现多层结构,结构紊乱边缘不齐,可见大量幼稚软骨细胞,软骨成熟度较低(图1B)。
2.2 Ckip-1对小鼠ATDC5软骨细胞系功能的影响
siRNA干扰技术对ATDC5细胞的转染效率大于90%,细胞内Ckip-1转录及翻译水平均被下调(图2),成功构建了Ckip-1敲低的体外细胞模型。
2.2.1 Ckip-1下调引起软骨细胞形态改变 光镜下观察可见敲低组软骨细胞伸展范围增大,细胞突触增多、增粗,细胞由三角形向多角形转变。同时胞内可见更多颗粒物,细胞核更为明显(图3)。电镜观察可见,在长期培养后,阴性对照组细胞呈现明显的极性,细胞沿某一方向伸展(图3),而敲低组中,细胞伸展范围较大,细胞极性不明显,呈现较为“杂乱”的排布方式。
2.2.2 Ckip-1下调引起软骨细胞增殖能力改变Ckip-1下调引起软骨细胞增殖能力改变,具体:①与阴性对照组相比,敲低组细胞自第3日起开始呈现显著的增殖抑制现象(图4);②克隆形成实验也得到相似结果,即敲低组细胞克隆形成率显著降低,提示细胞贴壁数量或单个细胞增殖能力下降(图4)。
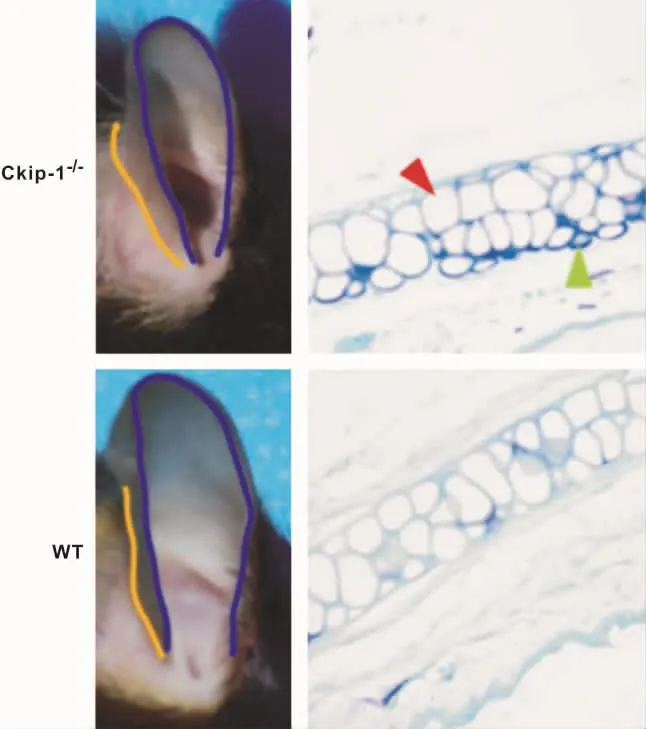
图1 Ckip-1-/-小鼠耳廓大体及组织学表型Fig 1 The macroscopic and histologic phenotype of the auricle of Ckip-1 -/- and WT mice
2.3 Ckip-1对软骨分化关键分子的影响
利用QPCR技术,探究Ckip-1下调对于软骨细胞分子层面的影响,具体:① Ckip-1下调在软骨细胞中可激活Bmp信号通路,与骨组织相似;②敲低组软骨早期分化关键分子Sox9水平上升(图5);但软骨细胞主要功能分子Col II的水平下降(图5);③敲低组软骨肥大化标志物Runx2与Col X呈现相反的变化趋势(图5),其具体原因有待进一步证明。
3 讨论
本实验通过动物现象观察、组织切片分析、细胞功能评估及关键分子检测等不同层面,系统研究Ckip-1对小鼠耳廓软骨发育畸形的可能原因,对解释病因及临床诊治提供实验依据。Ckip-1分子在细胞内广泛分布,其分子结构包括在N端的血小板-白细胞C激酶底物相关结构域(Pleckstrin Homology,PH),C端的亮氨酸拉链结构(Leucine Zipper,LZ)等多个分子结合位点,这些位点为其提供了介导多种蛋白及基因生物学行为的可能[7]。而其对软骨组织及细胞的影响可能基于以下几方面:
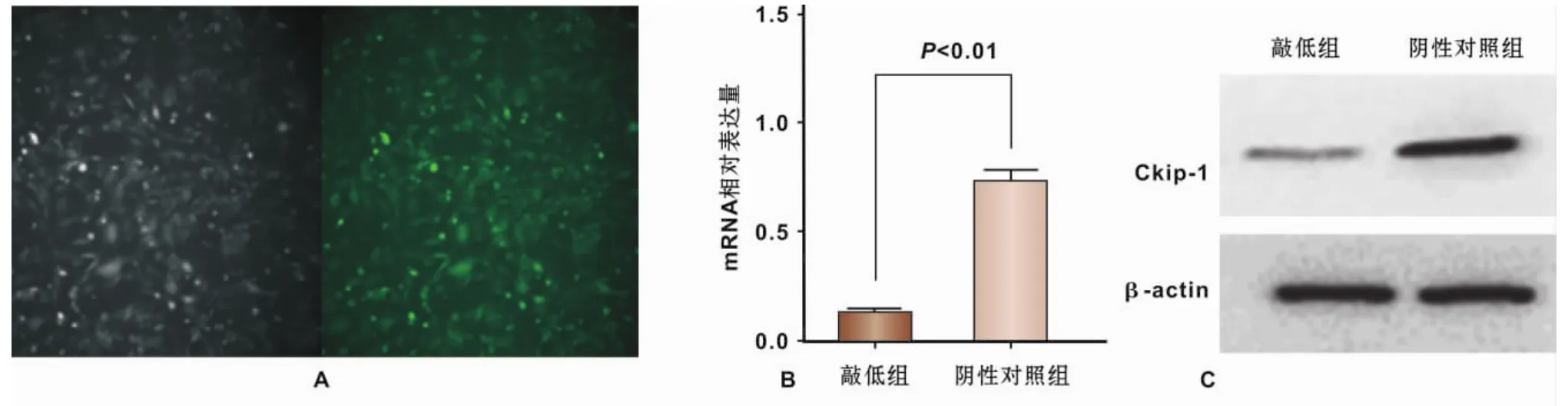
A:转染效率;B:Ckip-1转录水平(QPCR);C:Ckip-1蛋白翻译水平(Western Blot)图2 siRNA干扰技术建立Ckip-1下调的ATDC5软骨细胞系A:Transfection efficiency;B:Ckip-1 mRNA expression;C:Ckip-1 protein expressionFig 2 Down-regulation of Ckip-1 in ATDC5 chondroncytes by siRNA
3.1 Ckip-1调控软骨细胞生存
细胞生存状态直接决定了细胞功能的行使甚至组织的构成。本实验中,敲低组ATDC5中,细胞出现增殖及生存能力减弱的现象,这可能是由于Ckip-1对细胞的生存具有调控作用。在细胞质内,Ckip-1蛋白可以被Caspase-3在D310到D345区域裂解后入核,并通过直接结合或者抑制c-Jun磷酸化等方式发挥对AP-1的抑制作用,进而促进细胞凋亡[8]。同时,Ckip-1也能通过上调p53 N-末端丝氨酸-15的磷酸化水平来抑制p53的降解,进而抑制细胞的增殖能力[9]。但在肺癌的研究中,却得到相反的结论,下调Ckip-1基因水平可以有效抑制肺癌H1299细胞系的增殖和克隆形成能力,导致S期细胞周期阻滞和G2期的推广,以及显著增强H1299细胞系凋亡[10],同时这表明Ckip-1对于细胞生存的调节存在细胞特异性。本实验结果表明Ckip-1在软骨细胞生存中的作用与大多数组织,尤其是骨组织的显著差异,并最终导致动物小耳畸形的发生,提示Ckip-1的稳态对颅颌面组织发育的重要意义。

图3 Ckip-1对ATDC5软骨细胞系细胞形态影响Fig 3 The effect of Ckip-1 on the cell morphology of ATDC5 chondrocytes
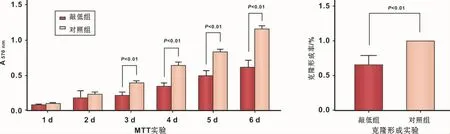
图4 Ckip-1对ATDC5软骨细胞系细胞生存能力影响Fig 4 The effect of Ckip-1 on the proliferation of ATDC5 chondrocytes

图5 Ckip-1对ATDC5软骨细胞系分化关键因子表达的影响Fig 5 The effect of Ckip-1 on the mRNA level of chodrocyte differentiation related factors in ATDC5 chondrocytes(QPCR)
3.2 Ckip-1调控软骨组织微观结构
软骨组织由软骨细胞及细胞外基质等构成,其中,软骨细胞形态及软骨细胞间连接状态等在软骨组织微观结构的基础。本实验中,下调Ckip-1将导致软骨细胞伸展范围的增加,而电镜结果也显示更多板状伪足的产生。这可能是由于Ckip-1对于细胞骨架具有调节作用。研究报道,Ckip-1可以通过抑制CP(一种广泛表达的异源二聚体,其可绑定到肌动蛋白丝上并发挥阻止添加或移除活性亚基的作用),进而促进肌动蛋白丝的延伸而抑制其分叉[11-13],这解释了细胞伪足的多方向伸展状态。同时,肿瘤学的研究也证实,Ckip-1可以通过Ckip-1/CK2/PAK1通路对基于细胞骨架功能的质膜变形和细胞迁移产生作用[14]。而这种细胞伪足功能的改变,将对细胞的连接及迁移产生影响,引起细胞分散和极性降低,并可能最终导致软骨组织微结构的不良,而这与组织学切片结果中软骨板形态不甚规则相一致。
3.3 Ckip-1调控软骨分化
软骨分化是软骨组织行使功能的基础。本实验中,下调Ckip-1导致:①Sox9水平升高,而Sox9作为软骨早期分化的重要标志物,可通过结合于PTHrP基因的启动子区并增加该启动子活性等方式,进而促进软骨细胞增殖和分化;② Runx2水平下降,而Runx2是软骨形成过程中矿物沉积阶段的特征性产物,其表达降低将导致软骨细胞成熟进程减缓[15-17];③ 软骨胶原分泌改变,本实验中,II型胶原的降低与X型胶原升高相伴,提示软骨基质分泌不良,但是否同时指明软骨朝末期分化加速仍需进一步实验证明[18-20];④Bmp信号通路的激活,与骨中较为类似,但不同的细胞生理效应提示其可能同时通过其他通路影响软骨发育[21]。这些结果共同解释了本实验中组织学分析中,软骨细胞成熟程度降低这一现象。
4 总结
在耳廓软骨发育过程中,Ckip-1可能通过改善细胞增殖及生存状态,调节分化进程等途径发挥重要作用,其缺乏将导致小鼠小耳畸形。
[1] 林琳,潘博,蒋海越,等.小耳畸形家系的基因定位及候选基因筛查初步研究[J].中国优生与遗传杂志,2013,21(6):17-19.
[2] Luquetti DV,Leoncini E,Mastroiacovo P.Microtia-anotia:A global review of prevalence rates[J].Birth Defects Res A Clin Mol Teratol,2011,91(9):813 -822.
[3] Zhang YB,Hu J,Zhang J,et al.Genome-wide association study identifies multiple susceptibility loci for craniofacial microsomia[J].Nat Commun,2016,7:10605.
[4] Zhang L,Wu K,Song W,et al.Chitosan/siCkip-1 biofunctionalized titanium implant for improved osseointegration in the osteoporotic condition[J].Sci Rep,2015,5:10860.
[5] Zhou ZC,Che L,Kong L,et al.CKIP-1 silencing promotes new bone formation in rat mandibular distraction osteogenesis[J].Oral Surg Oral Med Oral Pathol Oral Radiol,2017,123(1):e1-e9.
[6] 秦东泽,胡奥,郭威孝,等.CKIP-1对不同时间和部位松质骨影响的Micro-CT分析[J].口腔医学,2017,37(3):198-202.
[7] Nie J,Liu L,He F,et al.CKIP-1:A scaffold protein and potential therapeutic target integrating multiple signaling pathways and physiological functions[J].Ageing Res Rev,2013,12(1):276-281.
[8] Zhang L,Xing G,Tie Y,et al.Role for the pleckstrin homology domain-containing protein CKIP-1 in AP-1 regulation and apoptosis[J].EMBOJ,2005,24(4):766 -778.
[9] Zhang L,Tie Y,Tian C,et al.CKIP-1 recruits nuclear ATM partially to the plasma membrane through interaction with ATM[J].Cell Signal,2006,18(9):1386 -1395.
[10] Chen GM,Ding RF,Tan YD,et al.Role of the CKIP1 gene in proliferation and apoptosis of the human lung cancer cell line H1299[J].Genet Mol Res,2015,14(2):4005-4014.
[11] Bruck S,Huber TB,Ingham RJ,et al.Identification of a novel inhibitory actin-capping protein binding motif in CD2-associated protein[J].J Biol Chem,2006,281(28):19196-19203.
[12] Canton DA,Olsten ME,Niederstrasser H,et al.The role of CKIP-1 in cell morphology depends on its interaction with actin-capping protein[J].J Biol Chem,2006,281(47):36347-36359.
[13] Canton DA,Olsten ME,Kim K,et al.The pleckstrin homology domain-containing protein CKIP-1 is involved in regulation of cell morphology and the actin cytoskeleton and interaction with actin capping protein[J].Mol Cell Biol,2005,25(9):3519-3534.
[14] Kim YB,Shin YJ,Roy A,et al.The role of the pleckstrin homology domain-containing protein CKIP-1 in activation of p21-activated kinase 1(PAK1)[J].J Biol Chem,2015,290(34):21076-21085.
[15] Komori T.Runx2,a multifunctional transcription factor in skeletal development[J].JCell Biochem,2002,87(1):1-8.
[16] Zheng Q,Zhou G,Morello R,et al.Type X collagen gene regulation by Runx2 contributes directly to its hypertrophic chondrocyte-specific expression in vivo[J].J Cell Biol,2003,162(5):833-842.
[17] Zhang Y,Yang TL,Li X,et al.Functional analyses reveal the essential role of SOX6 and RUNX2 in the communication of chondrocyte and osteoblast[J].Osteoporos Int,2015,26(2):553-561.
[18] Narcisi R,Quarto R,Ulivi V,et al.TGF β-1 administration during ex vivo expansion of human articular chondrocytes in a serum-free medium redirects the cell phenotype toward hypertrophy[J].J Cell Physiol,2012,227(9):3282-3290.
[19] Ahmad PJ,Trcka D,Xue S,et al.Discoidin domain receptor-1 deficiency attenuates atherosclerotic calcification and smooth muscle cell-mediated mineralization[J].Am J Pathol,2009,175(6):2686 -2696.
[20] Aigner T,Neureiter D,Campean V,et al.Expression of cartilage-specific markers in calcified and non-calcified atherosclerotic lesions[J].Atherosclerosis,2008,196(1):37-41.
[21] 田晓光,沈舒宁,段银钟,等.CKIP-1对骨髓间充质干细胞体外增殖和分化能力的影响[J].实用口腔医学杂志,2016,32(1):10-14.
