罗汉果苷ⅢE的酶法合成
2018-07-02陈依军王淑珍吴旭日
陈 玲,陈依军,王淑珍,吴旭日
(中国药科大学生命科学与技术学院化学生物学研究室,南京 210009)
罗汉果(Siraitiagrosvenorii)是一种葫芦科多年生草本植物,其成熟果实是治疗咽炎、咽痛和咳嗽等的传统中药,也是开发天然低热量甜味剂的重要来源之一[1-2]。自1983年Takemoto等[3]从S.grosvenorii果实中分离出甜味化合物罗汉果苷(mogroside,MG)Ⅳ、Ⅴ和Ⅵ后,已有超过30种同系列化合物被陆续分离鉴定,它们均为罗汉果苷元[10α-cucurbit-5-ene-3β,11α,24(R),25-tetraol]连接2~6个葡萄糖单元形成的四环三萜皂苷。通过对罗汉果苷结构及其甜味的构效关系研究发现,结构中含3个葡萄糖单元的罗汉果苷均具有较好的甜味,但口味存在一定差别[1]。例如,罗汉果苷Ⅳ、Ⅴ和赛门苷Ⅰ的甜度分别是5%蔗糖的350倍、425倍和500倍以上,且口感相对纯正[4]。在全球倡导低热量健康饮食的大背景下,罗汉果苷无疑成为各大食品和饮料生产企业开发低(无)热量非糖类甜味剂的热点。
罗汉果苷作为罗汉果生长成熟过程中合成的次级代谢产物,获得性受到季节的限制,且绝大多数罗汉果苷的天然含量较低,难以通过提取分离实现规模化生产,严重阻碍了其被开发为优质天然甜味剂的可能性[5]。据报道,罗汉果苷Ⅴ(MG V)是罗汉果苷元3位和24位羟基分别β-连接2个和3个葡萄糖单元的皂苷,在罗汉果成熟果实中含量最高,也是目前唯一可产业化提取制备的罗汉果苷类化合物[6]。研究人员利用比较转录组学和体外活性验证手段初步阐释了罗汉果苷Ⅴ的完整生物合成途径[7-8]。从该途径中可以看出,罗汉果苷ⅢE(MG IIIE)是罗汉果苷Ⅴ生物合成的关键中间体,同样也是多种高甜度罗汉果苷Ⅳ、异罗汉果苷Ⅴ、赛门苷Ⅰ的生物合成前体,但其在罗汉果中的含量甚微,无法提取制备。如图1所示,选择性水解罗汉果苷Ⅴ结构中3位和24位以β-1,6-糖苷键连接的葡萄糖单元即可获得罗汉果苷ⅢE。但是,目前化学方法基本无法实现键专一性地水解罗汉果苷Ⅴ的2个β-1,6-糖苷键,因此,采用专一性强的糖苷水解酶建立绿色化酶法合成工艺成为最佳选择[9-11]。本研究以价低易得的罗汉果苷Ⅴ作为底物,筛选酶库获得了特异性合成罗汉果苷ⅢE的糖苷水解酶CPU-GH117,通过诱导表达条件和催化反应条件的系统优化,建立了罗汉果苷Ⅴ选择性水解制备罗汉果苷ⅢE的酶法合成新工艺,为其他高甜度罗汉果苷的制备和甜味剂开发提供了物质基础。

Figure1 Enzymatic synthesis of mogroside IIIE (MG IIIE) from mogroside V (MG V)
1 材 料
1.1 药品与试剂
罗汉果苷Ⅴ和ⅢE(成都德思特生物技术有限公司);卡那霉素、异丙基硫肽半乳糖苷(IPTG)(上海生工生物工程有限公司);三羟甲基胺基甲烷、丙烯酰胺、N′,N′-甲叉双丙烯酰胺、十二烷基磺酸钠、四甲基乙二胺(南京生兴生物技术有限公司);蛋白质相对分子质量标准(美国Fermentas公司);乙腈和甲酸(色谱纯,美国Tedia公司);蛋白质浓度测定试剂盒(江苏凯基生物技术有限公司);蛋白胨和酵母粉(英国Oxoid公司);其余试剂均为市售分析纯。糖苷水解酶库(共有57种酶)由中国药科大学化学生物学研究室前期创建。
1.2 仪 器
Agilent 1260高效液相色谱仪、Agilent 6224液质联用色谱仪(美国Agilent科技有限公司);YMC-Pack ODS-A C18色谱柱和C18硅胶填料(日本YMC股份有限公司);pH计(瑞士梅特勒公司);高压细胞破碎仪(英国Constant Systems公司);蛋白质电泳仪(美国Bio-Rad公司);冻干机(上海Christ仪器有限公司)。
2 方 法
2.1 罗汉果苷Ⅴ和ⅢE的HPLC分析
色谱条件:色谱柱为Agilent C18反相柱(250 mm×4.6 mm,5 μm),流动相A为超纯水(含0.1%甲酸),流动相B为乙腈(含0.1% 甲酸),洗脱梯度为10%~90% B,梯度洗脱时间为25 min,进样量为10 μL,柱温为30 ℃,检测波长为210 nm;流速为1 mL/min。在该色谱条件下,罗汉果Ⅴ和ⅢE的保留时间分别为10.0 min和11.3 min。
2.2 糖苷水解酶库的诱导表达和筛选
糖苷水解酶库中所有酶采用相同条件在24孔板中进行诱导表达:0.2 mmol/L IPTG、诱导温度15 ℃和诱导时间12 h。诱导表达完成后向菌液中加入罗汉果苷Ⅴ至0.5 mg/mL,于室温反应12 h,然后用等体积的甲醇停止反应,12 000 r/min离心10 min,上清液过滤后进行HPLC分析,以罗汉果苷ⅢE的生成情况筛选键专一性水解罗汉果苷Ⅴ的糖苷水解酶。
2.3 糖苷水解酶活性测定
以罗汉果苷Ⅴ的峰面积(A)对其质量浓度 (c,mg/mL) 线性回归制作标准曲线:A=1 212c+44.977,r2=0.999 2,线性范围为0.1~5 mg/mL。本研究以测定反应体系中罗汉果苷Ⅴ的减少量计算糖苷水解酶的活力。含糖苷水解酶的粗酶液为重组表达菌体经高压破碎和冷冻离心所得。酶活力测定体系为1 mL,含罗汉果苷Ⅴ 1 mg和0.1 mol/L NaOAc-HAc缓冲液(pH 7.0)。1个酶活单位(U)指在35 ℃条件下每分钟消耗1 μmol/L罗汉果苷Ⅴ所需的酶量。比活力为每毫克总蛋白所含有的活力单位数(U/mg)。
2.4 糖苷水解酶诱导表达条件考察
在0.2 mmol/L IPTG和诱导12 h条件下,分别考察诱导温度为15、20、25、30、35 ℃时糖苷水解酶的可溶性表达量。在15 ℃和诱导12 h条件下,分别考察IPTG用量为0.2,0.4,0.6,0.8,1.0 mmol/L时糖苷水解酶的可溶性表达量。利用最优诱导温度和IPTG用量考察4~24 h诱导时间条件下的糖苷水解酶可溶性表达量。
2.5 罗汉果苷ⅢE的生物合成反应条件考察
利用单因素分析方法考察了糖苷水解酶选择性水解罗汉果苷Ⅴ制备ⅢE的pH、温度、酶用量和底物浓度。pH考察范围为3.0~8.0,缓冲液种类为0.1 mol/L Tris-HCl(7.5~8.0)、0.05 mol/L NaOAc-HAc(4.5~6.0)以及0.05 mol/L Na2HPO4-柠檬酸(3.0~4.0,6.5~7.0);反应温度考察范围为25~65 ℃;酶量考察范围为0.5~5 U/mL;底物浓度范围为1~10 mg/mL。上述所有反应的体积均为1 mL,pH和温度考察的反应时间为8 h,酶用量和底物浓度考察的反应时间为24 h。
2.6 反应体系的放大验证
利用最优反应条件将水解罗汉果苷Ⅴ制备ⅢE的反应体系扩大至500 mL,并于不同反应时间点取样监测反应进程。反应结束后,反应液煮沸5~10 min,离心去除变性蛋白质,上清液冷冻干燥制备罗汉果苷ⅢE粗品。利用C18反相柱(内径2 cm,长度50 cm)分离纯化罗汉果苷ⅢE,洗脱液为含45%乙腈的水溶液。HPLC监测纯化过程,收集含罗汉果苷ⅢE的洗脱液,干燥后获得罗汉果苷ⅢE,最后利用HPLC检测样品纯度。
3 结 果
3.1 糖苷水解酶的筛选
本课题组前期建立了由57种糖苷水解酶组成的酶库,酶的来源涉及植物、真菌和细菌。本研究采用统一的诱导表达条件对所有酶进行了大肠埃希菌的异源表达,然后构建全细胞生物催化体系用于筛选键专一性水解罗汉果苷Ⅴ合成罗汉果苷ⅢE的糖苷水解酶。结果显示,表达Saccharomycescerevisiae糖苷水解酶CPU-GH17的大肠埃希菌细胞在实验条件下能够催化罗汉果苷Ⅴ水解生成单一产物MG-X,而其他全细胞催化剂水解罗汉果苷Ⅴ所得产物多为混合物(图2-A)。如图2-B所示,MG-X与罗汉果苷ⅢE对照品的保留时间完全相同,且其实测相对分子质量[M-H]-m/z=961.538 0与罗汉果苷ⅢE的相对分子质量的计算值961.545 0一致。上述结果表明,MG-X即为罗汉果苷Ⅴ选择性水解2个β-1,6糖苷键产生的ⅢE。因此,本研究选择糖苷水解酶CPU-GH17作为生物催化剂,以建立绿色高效的罗汉果苷ⅢE酶法合成新工艺。
3.2 CPU-GH17的可溶性表达
为提高糖苷水解酶CPU-GH17的可溶性表达量,以便于后续的反应条件考察,本研究对影响大肠埃希菌中重组蛋白可溶性表达的主要因素如IPTG用量、诱导温度和诱导时间进行了系统考察[12]。如图3-A所示,CPU-GH17在SDS-PAGE中测定的相对分子质量约为50 kD,与计算的相对分子质量51.6 kD基本一致。IPTG作为一种高效的乳糖启动子诱导剂,其浓度是影响重组蛋白表达的重要因素。如图3-B所示,CPU-GH17的可溶性表达量在IPTG摩尔浓度为0.4 mmol/L时达到最高值,约占可溶性总蛋白的16.5%,并随着IPTG用量增加逐渐降低。诱导温度过高,重组蛋白容易错误折叠形成包涵体。温度过低则会导致菌体生长缓慢,重组蛋白的总量减少[13]。从图3-C中可以看出,低温更利于CPU-GH17的可溶性表达,15 ℃的表达量可达到22.1%。在0.4 mmol/L IPTG和15℃条件下,CPU-GH17的表达量随着诱导时间延长逐渐增加,并在12 h时达到最高值22.8%(图3-D)。因此,CPU-GH17可溶性表达的最佳条件最终确定为:0.4 mmol/L IPTG、15℃和12 h。在该诱导条件下,CPU-GH27的比活力为0.049 U/mg。

Figure2 Selected CPU-GH17 for the biosynthesis of MG IIIE
A:HPLC analysis of (1) MG IIIE standard,(2) hydrolysis of MG V with CPU-GH17,(3) hydrolysis of MG V with CPU-GH24 and (4) MG V standard;B:LC/MS analysis of MG-X

Figure3 Optimization of expression conditions for CPU-GH17
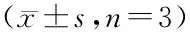
3.3 CPU-GH17生物合成罗汉果苷ⅢE的最佳条件
3.3.1 最佳反应温度 酶催化的反应速率与温度密切有关,反应温度越高越有利于反应的启动和进行,但温度过高会导致酶蛋白的变性失活[14],因此温度的选择对CPU-GH17水解罗汉果苷Ⅴ合成罗汉果苷ⅢE至关重要。本研究考察了25~65 ℃条件下罗汉果苷ⅢE的生成效率,结果显示,随着反应温度上升罗汉果苷ⅢE的生成率逐渐增加,并在45 ℃时达到最大值(图4-A)。此外,在45 ℃条件下孵育12 h,CPU-GH17酶活力降低不超过20%。因此,最终选择45 ℃作为最佳反应温度进行后续反应条件的优化。
3.3.2 最佳反应pH 反应pH的不匹配可能会导致酶变性失活以及延缓催化残基的质子化,从而降低反应效率[14]。如图4-B所示,糖苷水解酶CPU-GH17对反应pH变化较为敏感,其水解罗汉果苷Ⅴ的最佳pH为6.0,过高或过低都会导致反应效率的明显降低。
3.3.3 最佳酶用量和底物浓度 由于底物抑制和产物抑制的存在,生物催化反应中的酶的用量和底物浓度一般相互关联,互相影响[15]。为获得CPU-GH17催化合成罗汉果苷ⅢE的最佳底物浓度,本研究在反应温度45 ℃和pH 6.0的条件下综合考察了底物浓度和酶用量的相互关系。如图4-C所示,随着酶用量的增加,底物浓度可逐渐升高,且罗汉果苷Ⅴ可完全选择性转化为ⅢE。但是当质量浓度超过7.5 mg/mL水平时,底物浓度的增加会造成罗汉果苷ⅢE的产率逐渐下降,这可能是由于酶失活、底物抑制以及产物抑制等影响因素中的一种或多种综合作用所致。因此,最终确定制备罗汉果苷ⅢE的最佳条件确定为:45 ℃、pH 6.0、底物质量浓度为5 mg/mL、酶浓度为3 U/mL。

Figure4 Optimization of reaction conditions for CPU-GH17

3.4 罗汉果苷ⅢE的制备
在确定最佳反应条件后,本研究在500 mL规模(2.5 g)对其稳定性和放大可行性进行了考察,并用HPLC监测反应过程以确定最佳反应时间。如图5-A所示,反应进行20 h后罗汉果苷Ⅴ即可完全水解转化为罗汉果苷ⅢE,且随着反应时间的推移并未出现罗汉果苷ⅢE的降解。在反应完成后,反应液经过加热变性、离心去蛋白和冻干,获得罗汉果苷ⅢE的粗品2.52 g。粗品经过C18反向柱一步纯化和洗脱液浓缩冻干,最终获得了白色粉末状罗汉果苷ⅢE纯品1.96 g,计算回收率约为77.8%,HPLC检测纯度为97.8%(图5-B)。由此可见,本研究利用糖苷水解酶CPU-GH17建立了一种有望规模化制备罗汉果苷ⅢE的酶法合成新工艺,可为罗汉果苷甜味剂开发提供了重要合成前体。

Figure5 Bio-preparation of MG IIIE at 500 mL-scale

4 讨 论
罗汉果苷ⅢE作为多种高甜度罗汉果苷化合物合成的关键前体,其规模化制备将对罗汉果苷类甜味剂的开发产生重大影响[16]。在前期创建的糖苷水解酶酶库基础上,本研究利用可产业化生产的罗汉果苷Ⅴ作为底物,筛选得到能够键专一性水解罗汉果苷Ⅴ制备ⅢE的糖苷水解酶CPU-GH17。以蛋白质可溶性表达量为评价指标,确定了CPU-GH17在大肠埃希菌中的最佳诱导表达条件:0.4 mmol/L IPTG、15 ℃和诱导12 h。然后,在确定CPU-GH17催化合成罗汉果苷ⅢE的最佳反应条件(45 ℃、pH 6.0、5 mg/mL底物和3 U/mL CPU-GH17)后,反应体系被放大至500 mL,验证了工艺的稳定性和可行性。因此,罗汉果苷ⅢE的酶法合成新工艺已基本建立,为规模化制备其他高甜度罗汉果苷提供了可能。虽然目前该工艺的底物质量浓度仅能达到5 mg/mL,但是本研究正在利用定向进化手段改造CPU-GH17,以期提高其底物和产物耐受性以及催化效率,从而大幅度提升生物合成罗汉果苷ⅢE的底物浓度并降低成本。
参 考 文 献
[1] Wang L,Yang Z,Lu F,etal.Cucurbitane glycosides derived from mogroside IIE:structure-taste relationships,antioxidant activity,and acute toxicity[J].Molecules,2014,19(8):12676-12689.
[2] Akihisa T,Hayakawa Y,Tokuda H,etal.Cucurbitane glycosides from the fruits ofSiraitiagrosvenoriiand their inhibitory effects on Epstein-Barr virus activation[J].JNatProd,2007,70(5):783-788.
[3] Takemoto T,Arihara S,Nakajima T,etal.Studies on the constituents of fructus Momordicae.I.on the sweet principle[J].YakugakuZasshi,1983,103(11):1151-1154.
[4] Jia Z,Yang X.A minor,sweet cucurbitane glycoside fromSiraitiagrosvenorii[J].NatProdCommun,2009,4(6):769-772.
[5] Li D,Ikeda T,Huang Y,etal.Seasonal variation of mogrosides inLoHanKuo(Siraitiagrosvenorii) fruits[J].JNatMed,2007,61(3):307-312.
[6] Zhang M,Yang H,Zhang H,etal.Development of a process for separation of mogroside V fromSiraitiagrosvenoriiby macroporous resins[J].Molecules,2011,16(9):7288-7301.
[7] Itkin M,Davidovich-Rikanati R,Cohen S,etal.The biosynthetic pathway of the nonsugar,high-intensity sweetener mogroside V fromSiraitiagrosvenorii[J].PNatlAcadSci2016,113(47):E7619-E7628.
[8] Qi T,Ma X,Mo C,etal.An efficient approach to findingSiraitiagrosvenoriitriterpene biosynthetic genes by RNA-seq and digital gene expression analysis[J].BmcGenomics,2011,12(1):343-355.
[9] Van LJ,Faijes M,Nieto J,etal.Hydrolase and glycosynthase activity of endo-1,3-beta-glucanase from the thermophilePyrococcusfuriosus[J].Archaea,2004,1(4):285-292.
[10] Michlmayr H,Varga E,Malachova A,etal.A versatile family 3 glycoside hydrolase from bifidobacterium adolescentis hydrolyzesβ-glucosides of theFusariummycotoxins deoxynivalenol,nivalenol,and HT-2 toxin in cereal matrices[J].ApplEnvironMicrob,2015,81(15):4885-4893.
[11] Wang H,Yan Y,Lin L,etal.EngineeringSaccharomycescerevisiaewith the deletion of endogenous glucosidases for the production of flavonoid glucosides[J].MicroCellFact,2016,15(1):134.
[12] Francis DM,Page R.Strategies to optimize protein expression inE.coli[J].ProtocProteinSci,2010,5(24):1-29.
[13] Kaur J,Kumar A,Kaur J.Strategies for optimization of heterologous protein expression inE.coli:roadblocks and reinforcements[J].IntJBiolMacromol,2018,106:803-822.
[14] Vera C,Guerrero C,Wilson L,etal.Optimization of reaction conditions and the donor substrate in the synthesis of hexyl-β-d-galactoside[J].ProcessBiochem,2017,58:128-136.
[15] Prajapati VS,Trivedi UB,Patel KC.Optimization of glucoamylase production byColletotrichumsp.KCP1 using statistical methodology[J].FoodSciBiotechnol,2013,22(1):31-38.
[16] Mooradian AD,Smith M,Tokuda M.The role of artificial and natural sweeteners in reducing the consumption of table sugar:a narrative review[J].ClinNutrEspen,2017,18:1-8.
