陕西省韩城市利什曼原虫SSU rRNA基因系统发育研究
2018-06-21,,2,,,,
,,2,,,,
利什曼原虫所致的利什曼病,可分为内脏利什曼病(又称为黑热病)、黏膜皮肤利什曼病和皮肤利什曼病。引起不同临床类型的利什曼原虫在形态结构上没有明显差别,传统方法对虫株的鉴别主要依靠流行病学及临床资料[1]。基于分子研究的方法已被广泛运用于微生物和寄生虫的分类,相别于传统方法,具有快速、准确,且可与国内外数据比对构建进化关系的特点。SSU rRNA是真核生物一个较保守的基因,但在原虫的不同种间的变异比原核生物还要大,可用于原虫种属之间亲缘关系的研究[2-3]。本文研究分析了陕西省韩城市利什曼原虫标本的SSU rRNA基因片段,与国内外发表的该基因数据库进行比对,从分子水平确立陕西省利什曼原虫种属分类地位。
1 材料与方法
1.1 陕西省利什曼原虫阳性标本
1.1.1虫株标本 采集病犬及病例骨髓标本,以NNN培养基25 ℃进行培养,第10 d收集培养液镜下观察,发现有前鞭毛体存在者为培养阳性。取培养液用QIAamp DNA Mini Kit(Qiagen公司生产)提取基因组DNA。
1.1.2病例标本 3例有黑热病疫区接触史,症状典型的病例,取骨髓穿刺液,瑞氏染色,镜下观察发现典型利杜体者为阳性,取骨髓液用QIAamp DNA Mini Kit提取基因组DNA。
1.1.3媒介标本 采集病区白蛉标本,单只装管,冷冻研磨砂,以天隆动物组织基因组核酸提取试剂盒(西安天隆公司生产)提取基因组DNA,以kDNA引物RV1、RV2及K13A、K13B[4]扩增阳性者作为媒介感染的阳性标本。
1.2 SSU rRNA基因序列扩增及测序
1.2.1SSU rRNA引物及扩增条件 参照文献[5],研究序列SSU rRNA的上游引物为R222:TATTGGAGATTATGGAGCTG;下游引物R333:AAAGCGGGCGCGCGGTGCTG。由上海生工公司合成。PCR试剂盒由Takara公司生产,反应体系50 μL:10×PCR缓冲液5 μL,上下游引物各1 μL,脱氧核糖核苷三磷酸(dNTP,各2.5 mmol/L)1 μL,加水补齐至47.6 μL,加入模板DNA 2 μL,100 ℃,解链10 min,再加入Taq酶(2.5 U)0.4 μL,再按照94 ℃ 75 s,55 ℃ 1 min,72 ℃ 60 s循环32次,最后72 ℃延伸10 min。产物大小约395 bp。
1.2.2PCR产物测序 上述PCR产物由QIAxcel Advanced毛细管电泳仪(Qiagen公司生产)进行电泳检测,条件为AM420,条带清晰且产物大小符合目的序列的PCR产物送上海生工公司进行纯化,双向测序拼接,将拼接序列提交GenBank数据库。
1.3SSU rRNA基因分析及系统发育树 将获得序列通过Blast查询搜索,并参照文献[5-6]下载国内外利什曼原虫的SSU rRNA基因序列,与本研究所获得的序列共同构成分析的文件数据库。利用DNAStar软件中的Megallign模块进行序列比对,分析突变区域碱基的变化情况,与文献[7-8]报道进行比较。再利用Mega 6.06绘制系统进化树,进行同源性分析。
2 结 果
2.1SSU rRNA基因PCR扩增结果 获得1份利什曼原虫培养虫株,1份犬骨髓、3份病例骨髓、2份白蛉共7份利什曼原虫kDNA阳性标本。对7份标本进行SSU rRNA扩增,毛细管电泳有5份标本出现大小约400 bp的目的条带,见图1。来自白蛉的核酸标本未扩增出目的条带,有3份标本条带较淡,有一份出现杂带,未影响测序。将出现条带的样本进行双向测序,有3份标本测序成功,分别为QB41-1、QB41-2(单向测序)、Niufei。

C2:QB41-1,C4:Niufei,C5:Niufei-2,C9:JXF,C10:QB41-2,A12:Marker图1 SSU rRNA序列扩增结果Fig.1 SSU rRNA sequence amplification electrophoresis
对测序成功的3条序列提交GenBank,获得登录号分别为MG020105、MG020106、MG020107,下载文献[3,7-8]中的序列以及通过Blast获得的代表性利什曼原虫的SSU rRNA序列,组成本研究所用的序列数据信息表如表1。
2.2 利什曼原虫SSU rRNA序列分析
2.2.1序列同源性分析 利用DNAStar MegaAllign7.07进行比对分析,发现本研究的3条序列中有2条完全相同,另一条测序不完整。与GenBank登录号为HQ895849和HQ895854的序列在第49位均为C、第92位均为A,与其余序列有3-4个核甘酸的差异,序列对比图未列出,序列同源分析如图2,可见陕西MG020106_QB41-1、MG020107_Niufei标本与甘肃HQ895849和新疆HQ895854虫株100%同源,与其余国内利什曼原虫同源性在98%以上。
表1 本研究所用的利什曼原虫SSU rRNA序列信息
Tab.1 SSU rRNA sequence of Leishmania in this study

登录号虫株长度来源MG020105∗QB41-2365陕西韩城MG020106∗QB41-1398陕西韩城MG020107∗Niufei396陕西韩城HQ895845MHOM/CN/54/#3390北京HQ895846MHOM/CN/80/XJ801383新疆HQ895847MHOM/CN/?/GS4355甘肃HQ895849MHOM/CN/93/GS7393甘肃HQ895852MCAN/CN/86/SC7393四川HQ895853MHOM/CN/84/SD1393山东HQ895854IPHL/CN/77/XJ771394新疆HQ895859MHOM/CN/84/JS1392江苏M80291L.adleri882肯尼亚M80292L.brazilensis883-M80293L.mexicana883-M84225L.tarentolae2195-M81430L.chagasi882-M81428L.aethiopia882-X53912L.amazonensis2138巴西X53915L.major2137苏丹X53916L.tarentolae2137-X07773L.donovani2205苏丹AF13836L.colombiensis546-CQ277631L.mexicana563-XR_001203206L.infantum387-
*为本研究测序序列,其余下载自GenBank
2.2.2利什曼原虫SSU rRNA多变区段序列比较 将GenBank下载的序列与本研究的3条序列以Clustal W方法对齐,分析两个独特序列区UQ-I和UQ-II的序列点突变的情况,见表2。可见陕西3条序列点突变完全一致,不同于其余所有虫株。仅与甘肃HQ895849、新疆HQ895854、 印度L.d.DD8、M81430、苏丹X07773、XR_001203206虫株在374位有一个点位突变,而与国内其它虫株的突变差异较大。另外,陕西省虫株MG020105、MG020107在45、46位还有两个碱基缺失。在与国外的虫株比较中,在72位、312位、376附近还有碱基位点的变化,表中未列出。
表2 陕西省利什曼原虫与国内分离株SSU rRNA序列点突变的比较
Tab.2 Comparison of point mutations in Leishmania SSU rRNA sequences

虫株UQ-IUQ-II475292353374MG020105CTATCMG020106CTATCMG020107CTATCHQ895854CTATTHQ895849CTATTHQ895845TTGACHQ895846TTGACHQ895847TTGACHQ895852TTGACHQ895853TTGACHQ895859TTGTCM81430CTATTX07773CTATTXR_001203206CTATTL.d.SD2TTGTTL.d.DD8CTATTL.d.SC6TTGACL.d.SC10TTGACL.d.GS7TGGAC
2.3基于SSU rRNA基因序列的系统发育树 将本研究序列与GenBank下载的的SSU rRNA序列导入MEGA6.06,以Clustal W对齐后,构建系统进化树如图3,可见国内利什曼原虫聚为3支,陕西省的3条序列与杜氏利什曼原虫HQ895854、HQ895849、X07773、夏科氏利什曼原虫M81430以及婴儿利什曼原虫XR_001203206聚为一类。而其余来自北京、新疆、甘肃、四川等地的杜氏利什曼原虫聚在另一支。1株来自江苏的虫株与墨西哥利什曼原虫、硕大利什曼原虫、亚马逊利什曼原虫聚为一支。

图2 利什曼原虫SSU rRNA序列同源比较Fig.2 Sequence distence of Leishmania SSU rRNA gene
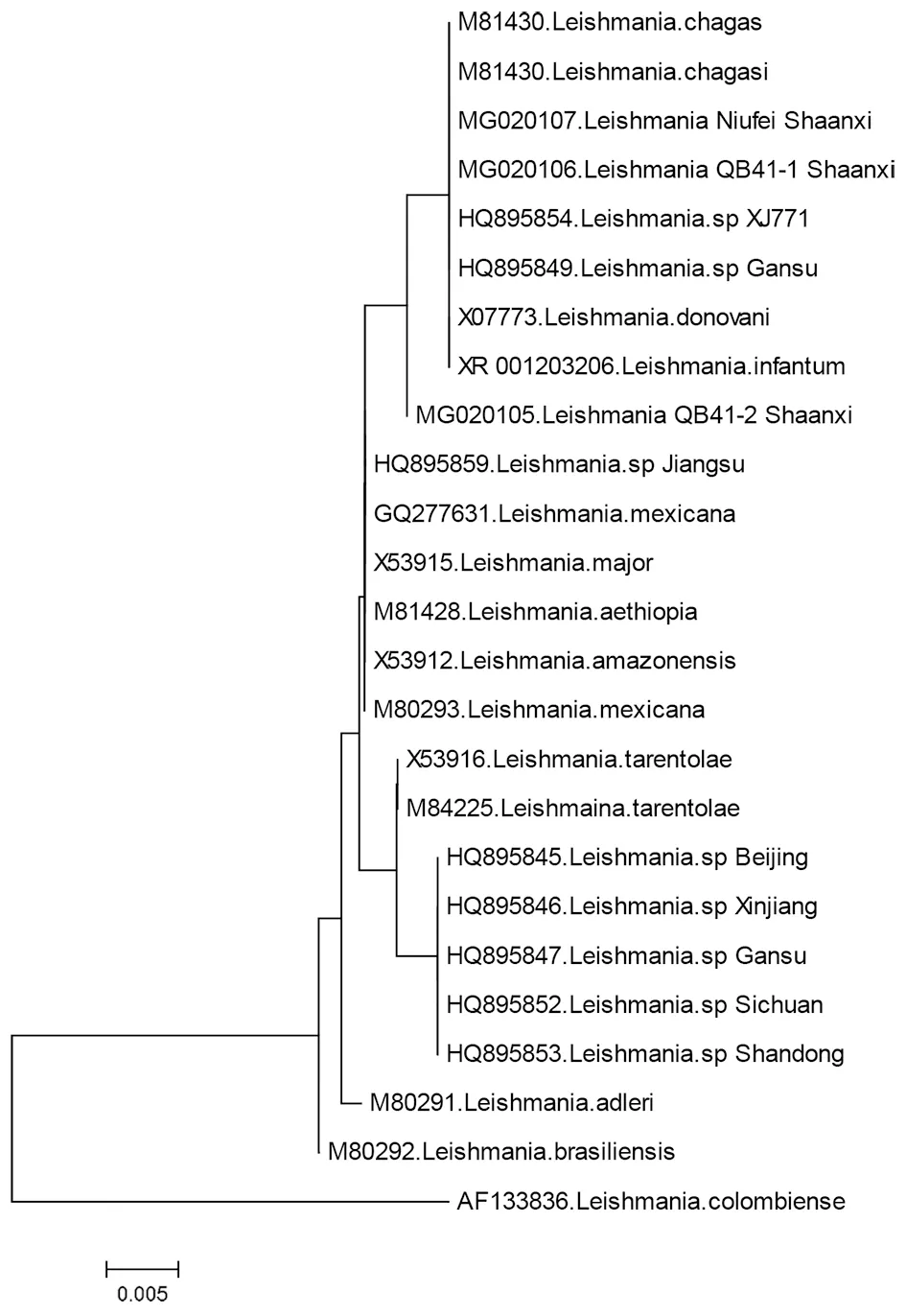
图3 基于利什曼原虫SSU rRNA基因序列的系统进化树Fig.3 Phylogenetic tree based on the SSU rRNA gene sequence of Leishmania
3 讨 论
陕西省是我国黑热病的传统疫区,解放前后对人民群众生命健康造成极大的危害。韩城市1967-2011年之间无病例发生。近年来在该地区屡见黑热病病例报告,甚至出现小的暴发点[9]。但陕西省缺乏利什曼原虫病原体的研究资料,2017年从韩城市病犬分离到1株利什曼原虫,并获得了数份利什曼原虫PCR阳性的标本。本文采用了SSU rRNA基因作为研究序列,以韩城市犬只、白蛉、病例的标本,对SSU rRNA基因片段进行PCR扩增,对扩增产物进行双向测序拼接,所获得的3条序列分别来自犬只与病例,将序列与国内外发表的SSU rRNA进行对比分析,构建了基于SSU rRNA的系统发育树。结果表明陕西省韩城市的标本与杜氏利什曼原虫、婴儿利什曼原虫以及夏科氏利什曼原虫聚为一类。
SSU rRNA(核糖体小亚基RNA)具有一个rRNA分子,在真核生物为18S,其基因序列及二级结构是至今发现的最为保守的一类核糖体基因,以此构建的系统发育树,在分子水平上更适合反映原虫的进化关系。本研究利用SSU rRNA构建的系统发育树表明,陕西省的利什曼原虫与新疆、甘肃、苏丹的杜氏利什曼原虫以及夏科氏利什曼原虫、婴儿利什曼原虫聚类为一支。而国内其余地区的利什曼原虫则聚在另一支,一株来自江苏的虫株与墨西哥利什曼原虫、硕大利什曼原虫、亚马逊利什曼原虫聚为一支。与曹得萍[3]、Guan W[6]等人的研究结果一致,从SSU rRNA的序列发育树来看,陕西省的原虫与新疆、甘肃、苏丹的杜氏利什曼原虫以及婴儿、夏科氏利什曼原虫的遗传关系近缘。此3类均是引起内脏利什曼病的原虫,世界卫生组织建议将其归属于杜氏利什曼原虫的种群[10]。Van Eys[4]等研究了9株利什曼原虫的SSU rRNA序列,认为其中存在2个独特的的突变序列区,分别是1-148位的UQ-1,包括7个突变点,229-392位的UQ-II包括2个突变点。卜毅玲[7-8]等人研究了5株利什曼原虫的SSU rRNA基因片段,发现在UQ-1上有3个碱基突变,UQ-II上有2个碱基突变。并认为在UQ-II区中,只有山丘疫区存在2个点突变,与平原疫区的L.d.SD2也存在碱基序列上的不同,认为我国山丘疫区与平原疫区存在种间的差异。本研究所分析的陕西省3条SSU rRNA序列不同于已发表的任何一条序列,特别是与国内众多利什曼原虫序列突变位点差异较大,仅与HQ895854、HQ895849在374位有一个位点突变(T变C)。同时还与来自印度的L.d.DD8、苏丹的X07773以及另一株夏科氏利什曼原虫在同样位点有突变(T变C),不同的是,苏丹的X07773和夏科氏虫株在376位有C碱基的插入。这些突变与郑学礼[11]等报道的新疆皮肤型利什曼原虫也有明显的差异。而芦殿梅[12]等用RAPD证实,印度平原型标准株L.d.DD8与国内荒漠型的HQ895854遗传距离较近,HQ895854虫株1977年分离自新疆塔里木硕大白蛉[3,7],属婴儿利什曼原虫。历史资料表明,陕西省黄土高原病区属犬源性疫区,病原体为婴儿利什曼原虫[13]。韩城市位于黄土高原与关中平原的交界地带,流行学资料显示犬只感染率高而人的感染率低,且以儿童发病较多,符合犬源性疫区特征,该疫区的原虫应为婴儿利什曼原虫[13-14],与SSU rRNA序列分析研究结果一致。
陕西省关中平原地区的人源性利什曼原虫疫区已消灭多年,陕北黄土高原及陕南山区偶有内脏利什曼病散发[15-16]。韩城市位于黄土高原与关中平原交界地带,自1967年以后再无病例报告,据郭晓莉[9]等报道,韩城市2012年暴发黑热病之前,有人从外地引进藏獒饲养,间隔45年再度出现利什曼病的流行,据报道[14,17]在犬源性疫区,野生动物也参与了传播过程,成为保存宿主之一。韩城的利什曼病是本地还是输入入值得探讨。虽然基于SSU rRNA的系统发育树中陕西虫株与新疆聚为一支,但突变位点来看与新疆1977年分离株有1个位点的差异,一方面SSU rRNA虽然可以代表一定的遗传变异关系,但毕竟序列较少,仍有局限性存在,另一方面二者相隔分离时间相差40年,是距离年代久远的变异还是陕西的虫株,由于缺乏陕西省利什曼原虫本底资料,还需要进一步的研究。
[1] 詹希美. 人体寄生虫学[M]. 北京:人民卫生出版社, 2010:80-87.
[2] Tenter AM. Current research on Sarcocyst is species of domestic animals[J]. J Parasitol. 1995, 25(11): 1311-1330.
[3] 曹得萍, 廖琳, 陈达丽,等. 利用SSU rRNA分子探讨中国利什曼原虫分子系统发育学[J]. 青海医学院学报, 2013, 34(1):25-30.
[4] 汪俊云, 陈生邦, 高春花,等. 甘肃文县婴儿利什曼原虫无症状感染犬的检测[J]. 中国人兽共患病学报, 2006, 22(8):734-737.
[5] van Eys GJ, Schoone GJ, Kroon NC, et al. Sequence analysis of small subunit ribosomal RNA genes and its use for detection and identification ofLeishmaniaparasites[J]. Mol Biochemic Parasitol, 1992, 51(1): 133-142.
[6] Guan W, Cao DP, Sun K, et al. Phylogenic analysis of ChineseLeishmaniaisolates based on small subunit ribosomal RNA (SSU rRNA) and 7 spliced leader RNA (7SL RNA)[J]. Acta Parasitologica, 2012, 57(2): 101-113. DOI: 10.2478/s11686-012-0022-9
[7] 卜玲毅, 胡孝素, 敬保迁,等. 利什曼原虫中国分离株SSUrDNA多变区序列分析[J]. 寄生虫病与感染性疾病, 2001, 9(1):1-3.
[8] 卜玲毅, 胡孝素, 敬保迁,等. 我国内脏利什曼病山丘疫区与平原疫区利什曼原虫SSUrDNA多变区序列分析[J]. 中国寄生虫学与寄生虫病杂志, 2000, 18(6):321-324.
[9] 郭晓莉, 薛小萍. 1例黑热病临床及其流行病学[J]. 医药前沿, 2016, 6(12):52-53.
[10] Schnur LF. Control of the leishmaniases. Report of a WHO Expert Committee. Technical Report Series 793[R]. WHO, 1990:25.
[11] 郑学礼, 敬保迁. 新疆皮肤利什曼病病原体SSU rRNA基因克隆与序列分析[J]. 中国寄生虫学与寄生虫病杂志, 2000, 18(5):260-262.
[12] 芦殿梅, 胡孝素, 乔中东,等. 用RAPD技术对利什曼原虫kDNA、nDNA的分析[J]. 寄生虫与医学昆虫学报, 2002, 9(1):1-6.
[13] 管立人, 瞿靖琦, 柴君杰. 中国利什曼病的现状和对开展防治工作的若干建议[J]. 疾病预防控制通报, 2000(3):49-53.
[14] 金长发, 洪玉梅, 熊光华,等. 中国犬源性和野生动物源性内脏利什曼病的研究进展[J]. 国际医学寄生虫病杂志, 2007, 34(5):227-230.
[15] 景彩霞, 曹志敏. 延安市幼儿内脏利什曼病1例[J]. 中国寄生虫学与寄生虫病杂志, 2014, 32(4):252-252.
[16] 薛亚娟, 刘志刚. 延安市2例内脏利什曼病病例报道[J]. 中国寄生虫学与寄生虫病杂志, 2016, 34(5):476-476.
[17] 陈凯, 伍卫平, 官亚宜. 利什曼原虫动物宿主研究进展[J]. 中国人兽共患病学报, 2015, 31(5):462-466. DOI: 10.3969/ j.issn.1002-2694.2015.05.015
