马铃薯花芽分化与内源激素动态变化的关系
2018-04-08艾星梅何睿宇胡燕芳
艾星梅,何睿宇,胡燕芳
(1 西南林业大学 园林学院,昆明 650224;2 迪庆州林业科学研究所,云南迪庆 674499)
植物的花芽分化一般而言均按照一定的程序进行[1],是营养生长向生殖生长转变的关键时期,经历形态学和生理学的复杂变化过程。大多数植物开花后加速植株进入衰老状态[2-3],而马铃薯由于遗传上的差异性,在整个生长发育过程中有些品种开花,有些品种开花较少或只结薯不开花,除了受外界温光的调控外[4-5],影响花芽分化的内因也是极其复杂的,其中包括生长期间各营养器官花序、茎叶、块茎间激烈的养分竞争[6],以及体内各种酶活性和碳水化合物存在一定的差异性[7]等。此外,内源激素也是调控植物花芽分化的重要物质,激素含量及比例会导致花芽分化生理状态的改变。因此,马铃薯花芽分化与多种指标的变化相关联。目前,有关马铃薯内源激素方面的研究已有不少报道,研究表明马铃薯生育周期长短、块茎产量、不同温光条件以及蕾花果脱落等均与内源激素含量有关。其中,生育期较短的品种GA3、IAA的含量均比生育期长的品种高,而ABA含量则较生育期长的低[8];在整个生长发育过程中,ABA含量的不断升高易引起蕾花果脱落,但与块茎产量呈极显著正相关,GA3/ABA、IAA/ABA、CTK/ABA 3个比值变化与块茎产量呈负相关[4,9];也有研究表明,ABA和JA含量升高与植株衰老的关系比与块茎形成的关系更为密切[10]。迄今,马铃薯内源激素含量变化与成花的关系研究鲜见报道。因此,本试验在研究马铃薯形态分化的基础上,进一步对开花与未开花马铃薯叶片的4种内源激素(GA3、IAA、ZR、ABA) 动态变化进行定量分析,以期了解马铃薯花芽分化的生理基础,揭示花芽不同分化阶段内源激素变化规律,为马铃薯花期调控、栽培和杂交育种提供理论依据。
1 材料和方法
1.1 试验材料
本试验以云南主栽的3个马铃薯品种为材料,包括2个开花品种‘云薯205’(YS205,开花较早,中晚熟)和‘合作88’(HZ88,开花较晚,晚熟),以及未开花品种‘云薯304’(YS304,无花,早熟)。供试材料于4月13日种植在西南林业大学后山基地,每个品种种植40个单株,株行距为30 cm×60 cm,均采用整薯播种,期间统一进行精细的田间管理,并详细观察记录各时期的花芽形态变化。
1.2 试验方法
1.2.1样品的采集样品的采集包括花芽分化前期、花芽分化初期、现蕾期、初花期和盛花期共5个时期,从4月26日(花芽分化前期)开始进行第一次采样,于上午9:00~10:00,对开花品种YS205的顶端叶片和花芽进行取样,未开花品种YS304的叶片采集时间与YS205同时进行,由于HZ88开花时间较YS205晚,为了使2个开花品种在生理性状上趋于一致,HZ88叶片和花芽的采集时期均比YS205延后5~7 d,采集后的样品迅速用冰盒带回实验室。HZ88和YS205样品采集标准为:第1个花芽分化开始为花芽分化初期;50%植株完成花芽分化为现蕾期;花序花朵开放量≥5%时为初花期;花序花朵开放量≥25%时为盛花期。
1.2.2花芽分化解剖观察用镊子及时剥除花芽和顶芽的苞片后放入FAA固定液(福尔马林∶冰醋酸∶70%酒精=5∶5∶90 )中固定,4 ℃保存。将固定好的材料采用常规石蜡切片方法[11],制成厚度8~10 μm切片,番红-固绿染色,中性树胶封片,置于显微镜(凤凰牌PH100-3B41L-IPL)下观察,软件Phmias 2008 Cs ver.3.0拍照。
1.2.3内源激素含量测定在解剖学的基础上,取0.2 g花芽分化相应时期的顶端叶片,经液氮速冻后,于超低温冰箱-80 ℃保存作为内源激素待测样。IAA、GA3、ZR和ABA测定采用酶联免疫吸附法(ELISA)提取、分离和检测[12],每个品种随机选取植株10株进行标记,每个时期采集3个样品,每个样品重复测定3次,取平均值。
试验数据采用Excel软件进行整理和统计分析。
2 结果与分析
2.1 马铃薯花芽发育期外部形态变化
马铃薯开花品种HZ88和YS205的花序为聚伞型花序,且花序较多。2个品种花芽发育的外部形态变化表现为: ①花芽分化初期:浅绿色的花萼包被花瓣,外表面密生绒毛,内表面毛状物很少;②现蕾期:随着花蕾膨大,花萼不断退缩,顶端5裂联合,雌雄蕊由花冠包被,柱头稍长,花粉和柱头趋于成熟,此期纵径的生长停止较横径早;③初花期:花萼表皮毛依然存在,花冠下部联合成管状,上部逐渐展开为五角星形, HZ88和YS205两个品种此时均已完成授粉受精;④盛花期:随着花朵开放时间的延长,成熟花粉粒不断从花药中纵裂扩散,柱头逐渐衰败枯萎;⑤果实发育期:当花败落后,已完成受精的植株花萼并不脱落,而是继续生长,顶端裂片逐渐合拢包裹果实,颜色加深成为深绿色,从本试验结果来看,HZ88具有一定的自交结实率,YS205无发育果实;⑥果实成熟期:HZ88的花萼横径增长停止较纵径早,当花萼不再膨大后,果皮由绿色变成黄白色或白色,至此浆果发育成熟。
2.2 马铃薯花芽分化的形态学观察
随着细胞分裂、生长与分化的进行,马铃薯花芽内部结构也发生着不可逆的变化。花芽分化前期为营养生长阶段,此时的芽体较小,生长点呈扁平状,外表平滑不突出,细胞体积小且排列紧密(图1,A)。进入生殖生长阶段后,根据花芽的发育进程及其所表现的结构特征,将马铃薯HZ88和YS205的花芽分化过程划分为2大发育时期,即花芽分化期和花器官发育期。
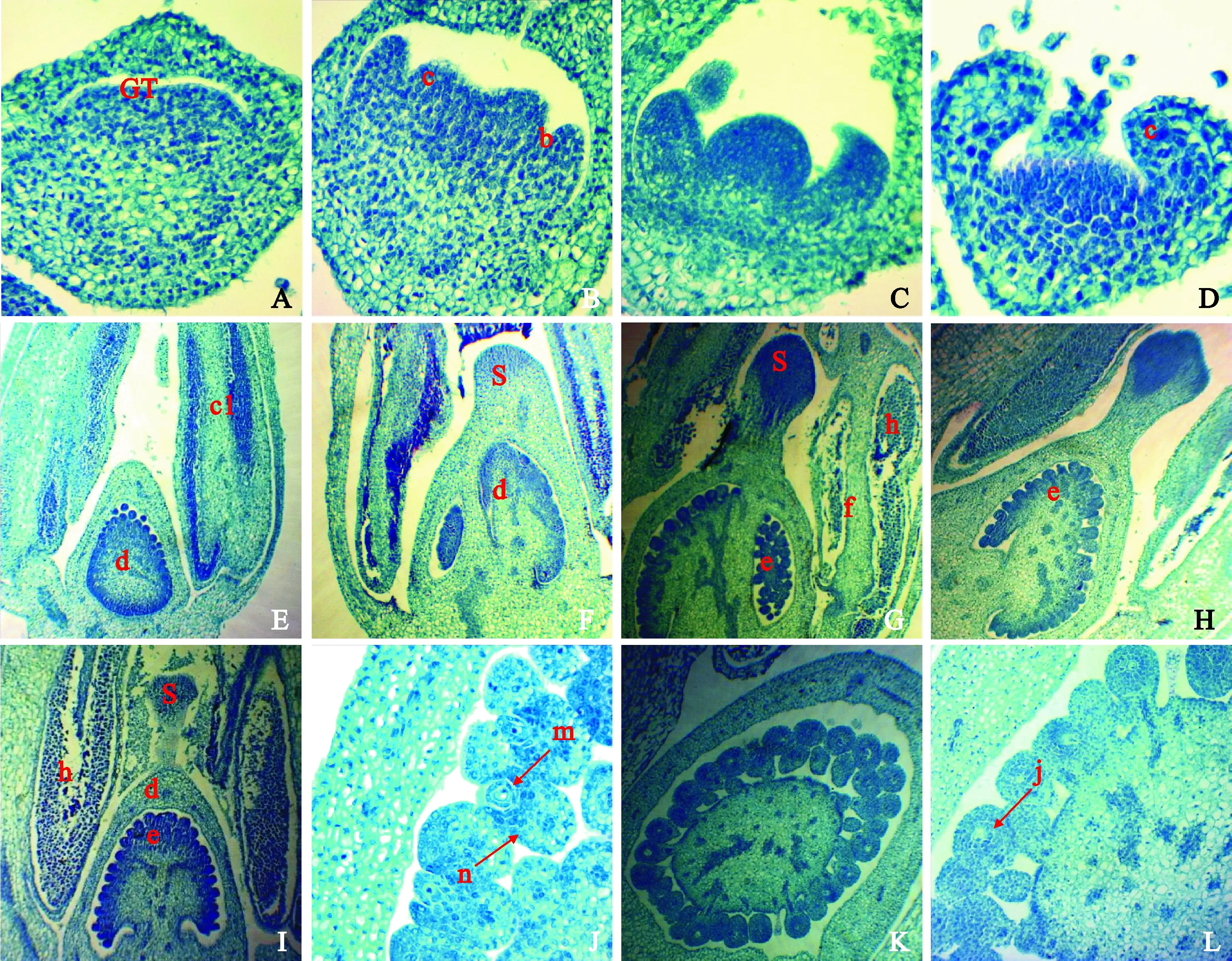
GT.生长锥;b. 花瓣原基;c. 雄蕊原基;c1.雄蕊;d.子房;e.胚珠;f.药隔;g.花粉囊;h.花粉粒;j.受精卵;m.精子;n.卵细胞;S.柱头A.花芽分化前期,×100;B~D.花芽分化期:花瓣原基、雌雄蕊原基形成期,×100;E.花器官发育初期,子房未分室,×40;F. 子房分室,花粉囊形成,×40;G~I.柱头花粉成熟期,×40;J.受精过程,×100;K~L. 完成受精,×40,×100图1 马铃薯花芽发育进程的纵切面解剖结构GT. Growing tip; b. Petal primordium; c.Stamen primordium; c1. Stamen; d. Ovary; e. Ovules; f. Connectivum; g. Anther; h. Pollen; j. Zygote; m. Sperm; n. Egg cells; S. StigmaA. Before flower bud differentiation, ×100; B-D: Flower bud differentiation stage: petal primordium, pistil and stamen primordium formation, ×100; E. Early stage of floral organ development, ovary was not divided, ×40;F. Ovary divided into chambers and anthers formed, ×40; G-I. Stigma and pollen maturity, ×40; J. Fertilization process,×100; K-L.Complete fertilization, ×40,×100Fig.1 Longitudinal section anatomy of potato flower bud during development stages
2.2.1花芽分化期在花芽分化初期前期,生长点细胞纵向分裂大于横向分裂,顶端分生组织逐渐扩展变宽,原体排列不规则的细胞明显增多,生长锥逐渐向上凸起趋于半球形(图1,B、C);到花芽分化初期后期,生长锥已拓展变宽,高度较小,两侧分生组织生长较快,内侧生长点分化突起发育为花原基,标志着植株由营养生长转变成生殖生长。半球状花原基逐渐变成扁圆形,周围细胞分裂较快,中央部位呈凹陷状,逐渐形成5个萼片原基(见横切面图2,A~C;纵切面仅见2个突起,图1,D)。萼片原基逐渐伸长并向内弯曲,呈镊合状排列,花开放后萼片大小一致、向外伸展;萼片原基内侧基部形成5个与萼片相间的小突起,即花瓣原基(横切面解剖结构,图2,D),最终发育成5瓣连接的轮状花冠。萼片和花瓣继续向上伸长,生长锥持续加宽,在基部形成一轮雄蕊原始体突起,即5个雄蕊原基,雄蕊原基逐渐分化,顶端发育为花药。随着雄蕊原基的分裂,内侧形成宽大的突起,即为雌蕊原基。雌蕊原基形成后快速分化生长,形成子房,花芽分化过程完成,进入花器官发育阶段。
2.2.2花器官发育期马铃薯花器官发育初期,雌雄蕊快速生长,在雌蕊心皮发育过程中,子房中轴产生一团组织突起,进而形成胚珠,对花蕾纵切面解剖结构显示(图1,E),子房逐渐分室,呈卵圆形,花瓣由花萼包被;此时花药是一团同形的分裂活跃的细胞,外面有1层表皮,每个花粉囊没有明显的药隔分开。随着花器官的发育,每个花药中的花粉囊逐渐形成,且两室子房清晰可见(图1,F;图2,E),内有多个胚珠。现蕾期,形成5个完整的花药,每个花药中的4个花粉囊由药隔完全隔开,呈对称的“U”字形(图2,F),花药壁由1层表皮、1层花药内壁、2层中层和1层绒毡层组成。花粉母细胞进行减数分裂过程中,花粉囊壁的绒毡层开始解体(图2,G、H),并释放出大量的营养物质,而花粉粒从解体的绒毡层细胞中不断吸取营养;从横切面解剖结构图可以看出(图2,I~L),细胞增大,最后分离形成游离的单核花粉粒,此时的花粉粒细胞壁薄,单核花粉进一步发育分裂形成2个子核,1个生殖核,1个营养核;最后形成成熟花粉粒,同时花粉囊壁解体,进行散粉(图1,G~I;图2,M)。此时子房发育成熟,胚珠饱满(图2,N),柱头呈头状,分泌大量黏液,是进行自花授粉的最佳时机。成熟花粉粒沿着花粉管通道进入子房到达胚珠,与卵细胞结合形成受精卵,完成受精(图1,J~L;图2,N、O)。马铃薯子房通常具有两室,在本试验中发现,开花较早的YS205和开花较晚的HZ88在现蕾前期花器官发育均呈一致的变化趋势,随着花器官的不断发育,YS205的子房逐渐形成完整的三室(图2,J、P)。
2.3 马铃薯花芽分化过程中叶片内源激素的变化
2.3.1IAA含量在整个花芽分化过程中,3个马铃薯品种叶片的IAA含量均呈先升后降的单峰曲线变化(图3,A)。其中,在花芽分化前期,开花品种YS205和HZ88叶片内IAA含量均显著低于未成花YS304(P<0.05);3个品种叶片IAA含量随着花芽发育逐渐上升,当开花品种处于现蕾期时,各品种的IAA含量均达到最大值,在此过程中未成花品种YS304的IAA含量和上升幅度均显著高于2个开花品种;随后,3个品种的IAA含量迅速下降,尤其以YS304下降幅度最大,至盛花期时,3个品种的IAA含量均降到最低,此时YS205、HZ88和YS304的IAA含量分别比最高值时下降了44.27%、52.34%和55.71%。可以看出,马铃薯有花芽分化植株IAA含量在整个分化过程中均显著低于同期无花芽分化植株且变化幅度较小,说明低水平的IAA含量有助于马铃薯花芽分化、促进花的形成。
2.3.2GA3含量3个马铃薯品种叶片中GA3含量动态变化与IAA相似,也呈单峰曲线;在整个花芽分化过程中,YS205和HZ88的GA3含量变化幅度趋于一致,两者在花芽分化初期和现蕾期低于YS304,而在初花期和盛花期高于YS304(图3,B)。其中,在花芽分化前期,3个品种叶片的GA3含量均较低;随着花芽发育的进行,3个品种的GA3含量先升后降,当YS205和HZ88处于现蕾期时,各品种叶片的GA3含量均达到最高值,并以YS304最高(1211.56 ng·g-1);当2个品种处于盛花期时,YS304的GA3含量迅速降到最低,仅为361.38 ng·g-1,并与同期开花品种相比差异显著。可以看出,未开花品种YS304叶片的GA3上升和下降速度在整个花芽分化过程中均明显高于开花植株YS205和HZ88,且变化幅度更大。
2.3.3ZR含量由图3,C可知,在马铃薯花芽分化过程中,3个品种叶片的ZR含量在花芽分化前期均较低,但开花品种YS205和HZ88均高于未成花品种YS304,之后三者的ZR含量均逐渐上升;到现蕾期时,3个品种的ZR含量均达到各自的最高值;随着花朵的开放,YS205和HZ88的ZR含量略有下降且趋于平缓的趋势,当YS205和HZ88处于盛花期时,未开花品种YS304的ZR值降到最低,略高于花芽分化前的水平。在整个过程中,成花品种YS205和HZ88的ZR含量始终高于未成花植株,而且差异显著。
2.3.4ABA含量3个马铃薯品种叶片内ABA含量在花芽分化过程中呈逐渐上升的趋势,且未成花品种始终低于成花品种(图3, D)。其中,在花芽分化前期,成花品种YS205和HZ88和未成花品种YS304的ABA 含量均较低;随着生育进程的推进,3个品种的ABA含量持续升高,且开花品种YS205和HZ88的上升速度较快;到盛花期时,三者均达到最大值,但YS205、HZ88和YS304的ABA含量之间均存在显著差异性。在整个花芽分化过程中,马铃薯ABA的含量表现为YS205>HZ88>YS304。
可见,在整个分化过程中,较低水平的IAA和GA3含量有利于启动马铃薯花芽分化,成花品种的ZR和ABA含量始终高于未成花植株,说明高水平的ZR和ABA易促进花原基的形成。
2.4 马铃薯花芽分化过程中叶片内源激素平衡与花芽分化的关系
ABA/IAA、ABA/ GA3、ZR/IAA、ZR/ GA3体现了植物内源激素间的动态变化,在整个花芽分化过程中,马铃薯叶片中的内源激素比值变化趋势如图4所示。首先,3个品种ABA/IAA和ABA/GA3比值在花芽分化前期均处于较低水平,后随着发育的进程两者比值均不断升高,且成花品种YS205和HZ88的比值始终高于未成花品种YS304(图4,A、B)。这说明较高水平的ABA含量,以及较低水平的IAA和GA3含量有利于马铃薯花芽分化,且ABA/IAA和ABA/GA3比值越高花芽分化越早。这也与各品种实际开花时间一致,品种YS205的开花时间比HZ88要早。
其次,在整个花芽分化过程中,马铃薯成花品种YS205和HZ88 叶片中ZR/IAA比值均高于未成花品种YS304,且YS304的ZR/IAA比值趋于平缓且处于较低水平。而叶片ZR/GA3的比值在花芽分化的不同阶段作用也不同。在花芽分化前期,成花品种YS205和HZ88的ZR/GA3比值高于未成花植株YS304,即ZR含量越高或GA3含量越低,越有利于营养物质的积累;随后,3个品种的ZR/ GA3随着发育进程均有下降趋势,至现蕾期时均降到最低;之后,各品种的ZR/ GA3比值逐渐回升(图4,C、D)。可以看出,马铃薯成花品种的ZR/IAA、ZR/ GA3比值始终高于未成花品种。

图4 3个马铃薯品种花芽分化过程中的各种内源激素比值变化Fig.4 The variation of ABA/IAA, ABA/GA3, ZR/IAA and ZR/GA3 during flower bud differentiation of three potato varieties
3 讨 论
花芽分化的形态建成是一个复杂的过程,马铃薯花期长短不一,花序的繁茂性也存在很大的差异性,每个植株上的一个花序持续时间为15~40 d不等,晚熟品种的开花期可长达2个月以上,但每朵花持续开放时间仅为5 d左右。根据马铃薯花芽分化各时期的纵切面和横切面解剖结构特征来看,从花芽分化初期,到雌雄蕊原基形成,完成整个花芽分化仅用了1周左右的时间。此后花器官不断发育,逐渐形成5个完整的花药和2室子房,其中YS205的子房在发育过程中发生了变异,逐渐形成完整的3室,这可能与子房发育的基因调控有关,目前尚不明确,还有待于进一步研究。随着细胞体积的增大,花粉粒和柱头逐渐成熟,胚珠饱满,是进行自花授粉的最佳时期,YS205和HZ88都先后经历了授粉受精过程,只有HZ88具有一定的自交结实率,YS205花蕾易脱落,无坐果率,这可能与内外多种因素的共同影响有关。此外,马铃薯花芽内部发育的同时,与外部形态之间有相对稳定的时序性对应关系,因此可以通过花芽的外部形态和大小对内部的发育进程进行初步判断。
内源激素对植物的生长发育具有一定的调控作用[13],其含量的高低能够促进或抑制花芽形态分化。前人研究表明,成花植株与未成花植株的内源激素含量变化明显不同。鲁亚婷等[14]研究表明,开花雷竹的ABA水平,以及ABA/IAA和iPA/IAA高于未开花植株,而IAA和GA水平低于未开花植株。何见等[15]报道,开花光皮树保持相对较低和稳定的IAA、GA3含量,ABA含量高且变化幅度较大,未开花光皮树IAA、GA3含量先升高后降低,ZR含量则相对较低,ABA含量先升高后下降。那光宇等[16]研究结果显示,什锦丁香花芽分化过程中成花枝条叶片中较低水平的IAA、GA3含量和较高水平的ZR、ABA含量有利于成花植株的花芽分化。本试验结果表明,马铃薯3个品种的IAA、GA3、ZR和ABA含量在花芽分化前期均较低,且无明显差异性,此时内源激素含量的高低并不能决定花芽是否分化;随着发育的进程,3个品种的IAA和GA3含量均呈先升后降的单峰曲线变化,其中开花品种YS205和HZ88的IAA和GA3含量在现蕾期以前均低于未成花品种YS304,开花品种达到现蕾期以后,YS304的IAA和GA3含量迅速下降;此外,YS205和HZ88的ZR和ABA含量则始终高于YS304,说明开花与否与内源激素含量的积累程度有关。值得一提的是,盛花期开花品种HZ88的ABA含量明显低于YS205,这可能与HZ88子房发育形成浆果有关,浆果内含有大量幼小种子,而幼小种子可以合成大量的IAA,对ABA具有一定拮抗的作用[4],因此,今后还需对果实的激素含量进行进一步研究。
不同激素并不是孤立地对成花过程发生作用,它们之间存在相互促进和相互拮抗的作用,Luckwill[17]首次提出了“激素平衡假说”,正是这种平衡状态,控制着核酸、蛋白质、可溶性糖和淀粉等物质的代谢,从而对植物的花芽分化和生长进行调控[18]。大量研究结果表明,ABA/IAA、ABA/ GA3、ZR/IAA、ZR/GA3体现了植物内源激素间的动态变化,比值的升高有利于促进花芽分化[19-20]。本研究中,开花马铃薯品种YS205和HZ88在花芽分化过程中的ABA含量大幅积累增加,ABA/IAA和ABA/GA3比值明显高于未开花品种YS304,说明ABA含量和比值升高有利于马铃薯花芽分化;同时,成花品种ZR/IAA、ZR/GA3的比值始终高于未成花品种。表明上述内源激素水平及比值对马铃薯花芽形态分化有相应的促进与抑制作用。
对于不同生育期的马铃薯而言,早熟品种比晚熟品种更早进入衰老期[21],用常规育种方法很难解释这种相关关系[22]。马铃薯开花与否除了与品种的遗传特性和环境有关外,还与不同发育时期的内源激素含量和比例变化相关联,各种激素处于一种动态的平衡状态,共同调节马铃薯的花芽形态分化过程。至于马铃薯成花品种与未成花品种营养物质的转移特征还需进一步结合块茎和其它矿质元素的变化来研究,从而为马铃薯花期调控、栽培和杂交育种提供理论依据。
参考文献:
[1]曲波,张微,陈旭辉,等.植物花芽分化研究进展[J].中国农学通报,2010,26(24):109-114.
QU B,ZHANG W,CHEN X H,etal. Research progress of flower bud differentiation mechanism of plant[J].ChineseAgriculturalScienceBulletin,2010,26(24):109-114.
[2]王双燕,李志宏,易同培,等.三月竹开花前后几种重要代谢物差异研究[J].广西植物,2016,36(2), 246-252.
WANG S Y, LI Z H, YI T P,etal. Difference of some important metabolites of flowering and non-floweringChimonobambusaopienensis[J].Guihaia, 2016,36(2), 246-252.
[3]蒋冬月,沈鑫,李永红,等.黄兰(MicheliachampacaL.)花开放和衰败过程中生理生化指标的变化[J].中国农学通报, 2013,29(16):125-128.
JIANG D Y, SHEN X, LI Y H,etal. Physiological and biochemical characteristics ofMicheliachampacaL. during florescence and flower senescence[J].ChineseAgriculturalScienceBulletin, 2013,29(16):125-128.
[4]门福义,王俊平,宋伯符,等. 马铃薯蕾花果脱落与内源激素和光照的关系[J].中国马铃薯,2000,14(4):198-201.
MEN Y F,WANG J P,SONG B F,etal. Relation between potato floral bud, flower and berry abscission and endogenous hormone and light[J].ChinesePotato,2000,14(4):198-201.
[5]TURNER A D AND EWING E E. Effects of photoperiod, night temperature, and irradiance on flower production in the potato[J].PotatoResearch, 1988,31(2): 257-268.
[6]王俊平,门福义,宋伯符,等.马铃薯营养物质分配对蕾花果脱落的影响[J].中国马铃薯,2001,15(1):1-4.
WANG J P, MEN F Y, SONG B F,etal. Effects of nutrient distribution on the abscission of bud, flower and berry in potato[J].ChinesePotato, 2001,15(1):1-4.
[7]艾星梅,何睿宇,徐永艳,等.马铃薯开花与未开花品种的生理生化差异性研究[J].江西农业大学学报,2017,39(2):230-236.
AI X M, HE R Y, XU Y Y,etal. A study of physiological and biochemical changes in flowering and non-flowering potatoes[J].ActaAgriculturaeUniversitatisJiangxiensis, 2017,39(2):230-236.
[8]刘梦芸,蒙美莲,门福义,等.马铃薯生育期间内源激素的变化[J].马铃薯杂志,1996,10(4):197-202.
LIU M Y, MENG M L, MEN F Y,etal. The change of intrinsic hormones in the growing period of potato[J].ChinesePotatoJournal,1996,10(4):197-202.
[9]刘梦芸,毛雪飞,门福义,等.马铃薯内源激素变化与块茎增大生长的相关规律[J].华北农学报,1997,12(2):86-92.
LIU M Y, MAO F X, MENG F Y,etal. Correlation law between the change of intrinsic hormones of potato tuber and growth of the tuber[J].ActaAgriculturaeBoreali-Sinica,1997,12(2): 86-92.
[10]肖关丽,郭华春.马铃薯温光反应及其与内源激素关系的研究[J].中国农业科学,2010,43(7):1 500-1 507.
XIAO G L, GUO H C. Sensitivity of Potato Cultivars(Solanum tuberosum L.) to Temperature and Photoperiod and the Relationship with Some Endogenous Hormone[J].ScientiaAgriculturaSinica,2010,43(7):1 500-1 507.
[11]段国禄,施江.植物制片、标本制作和植物鉴定[M].北京:气象出版社,2008:18.
[12]李宗霆,周燮.植物激素及其免疫检测技术[M].南京:江苏科学技术出版社,1996:80-298.
[13]符继红,孙晓红,王吉德,等.植物激素定量分析方法研究进展[J]. 科学通报,2010,55(33):3 163-3 176.
FU J H, SUN X H, WANG J D,etal. Progress in quantitative analysis of plant hormones[J].ChineseSciBull, 2010,55(33):3 163-3 176.
[14]鲁亚婷,袁晓亮,林新春,等.雷竹花芽形态分化过程中内源激素的变化规律[J].浙江农林大学学报,2012,29(2):161-165.
LU Y T, YUAN X L,LI X Z,etal. Endogenous hormone changes during floral bud morphological differentiation ofPhyllostachysviolascens[J].JournalofZhejiangForestryCollege,2012,29(2):161-165.
[15]何见,蒋丽娟,李昌珠,等. 光皮树花芽分化过程中内源激素含量变化的研究[J].中国野生植物资源,2009,28(2):41-45.
HE J, JIANG L J,LI C Z,etal. Changes of Endogenous Hormones, during the flower bud differentiation ofComuswilsonian[J].ChineseWildPlantResources,2009,28(2):41-45.
[16]那光宇,张姝媛,郭娜,等.什锦丁香花芽分化过程中植物内源激素的变化[J].内蒙古农业大学学报(自然科学版),2012,3(5-6):58-61.
NA G Y, ZHANG S Y, GUO N,etal. A study on the changes of plant hormones during flower bud differentiation stage inSyringachinensis[J].JournalofInnerMongoliaAgriculturalUniversity(Natural Science Edition),2012,3(5-6):58-61.
[17]LUCKWILL L C. The Control of Growth and Fruitfulness of Apple tree, Physiology of Tree Crops [M]. London:Academic Press,1970:237-254.
[18]段娜,贾玉奎,徐军,等.植物内源激素研究进展[J].中国农学通报,2015,31(2):159-165.
DUAN N,JIA Y K, XU J,etal. Research progress on plant endogenous hoemones[J].ChineseAgriculturalScienceBulletin, 2015,31(2):159-165.
[19]冯枫,杨际双.切花秋菊‘神马’花芽分化与内源激素的关系[J]. 中国农业科学,2011,44(3):552-561.
FENG F, YANG J S. Relationship between floral bud differentiation and endogenous hormones in autumn-cuttingChrysanthemummorifolium‘Jinba’[J].ScientiaAgriculturaSinica,2011,44(3):552-561.
[20]莫长明,涂冬萍,黄杰,等.罗汉果花芽分化过程中形态及其激素水平变化特征[J]. 西北植物学报,2015,35(1):98-106.
MO C M, TU D P, HUANG J,etal. Morphological and endogenous hormones characteristics of flower bud ofSiraitiagrosvenoriiduring its differentiation [J].ActaBotanicaBoreali-OccidentaliaSinica,2015,35(1):98-106.
[21]王冬雪,张丽莉,石瑛. 不同熟性马铃薯各生育时期功能叶生理指标变化的研究[J].中国农学通报,2014,30(3):124-128.
WANG D X, ZHANG L L, SHI Y. Research of the physiological indicators of different maturity potato (SolanumtuberosumL.) functional leaf ar each growth stages[J].ChineseAgriculturalScienceBulletin, 2014,30(3): 124-128.
[22]喻树迅,宋美珍,范术丽,等.短季棉早熟不早衰生化辅助育种技术研究[J].中国农业科学,2005,38(4):664-670.
YU S X, SONG M Z, FAN S L,etal. Studies on biochemical assistant breeding technology of earliness without premature senescence of the short-season upland cotton[J].ScientiaAgriculturaSinica,2005,38(4):664-670.
