红富士苹果叶片净光合速率与生理生态因子的关系
2018-01-18王传印包东娥李新艳刘遵春卞公明
王传印,包东娥,李新艳,刘遵春,卞公明
(1.菏泽市林业局,山东菏泽 274099;2.河南科技学院)
果树90%以上的干物质来源于叶片的光合产物[1]。净光合速率是指光合作用速率减去呼吸作用速率,体现了植物有机物的积累。研究光合作用,对于选育优良品种、改进栽培措施、提高单株产量起着重要的指导作用。国内外对苹果树叶片光合作用已做了大量的研究,笔者对红富士苹果的光合特性及其与主要环境因子之间的关系进行研究,为红富士优质高效栽培提供理论依据。
1 材料和方法
试验于2014年4月下旬在河南省科技学院果树教学基地进行,试验树红富士(MaluspumilaMill),5年生,株行距3m×4m,树高2.2m左右,正常肥水管理,树体健壮,长势一致,开花结果正常。当地年平均气温14.9℃,日照1928.5小时,年降水量656.3mm,无霜期220天。
用LI-6400型便携式二氧化碳(CO2)气体分析仪测定叶片净光合速率(Pn),同时记录大气温度(Ta)、光合有效辐射(PAR)、大气CO2浓度(Ca)、空气相对湿度(RH)等生态因子指标,叶片的蒸腾速率(Tr)、气孔导度(Gs)、细胞间隙CO2浓度(Ci)等生理因子参数。
随机选取5株长势一致的健壮植株,每株选择树冠东南方向外围发育枝条中部成熟叶片3片,挂牌标记。于晴朗天的8∶00~18∶00时段测定净光合速率(Pn)的日变化,每隔1小时测定1次,重复3次。所测数据用Microsoft Excel 2003处理,以平均值进行分析,对净光合速率与生态生理因子进行图表分析。
2 结果与分析
2.1 红富士叶片净光合速率与生态因子的日变化
图1A~D为红富士苹果叶片净光合速率日变化与4个生态因子的日变化曲线图。净光合速率日变化曲线呈双峰状,8时始,净光合速率快速上升,至10时左右达高峰值13.57μmolCO2·m-2·s-1。10时以后下降,至13左右至低谷值8.74μmolCO2·m-2·s-1(为上午高峰值的64.4%),此后1小时又缓慢上升,于14时左右出现第2个高峰值11.93μmol CO2·m-2·s-1(为上午峰值的87.9%)。在保持了较短时间的峰值后迅速降至最低。具有光合“午休”现象。一天中净光合速率的极差为9.3μmolCO2·m-2·s-1。
由图1A可看出,大气温度一直在27℃以下,叶片净光合速率于10时达高峰值、13时低谷值时,大气温度分别约20℃、24℃,所以此时气温不是影响净光合速率的主要因素。
由图1B可看出,光合有效辐射呈先升后降变化,12时达高峰值约1700μmol·m-2·s-1。10时,光合有效辐射1420μmol·m-2·s-1,净光合速率达高峰值;当光合有效辐射﹥1420μmol·m-2·s-1时,净光合速率下降。可以推断光合“午休”现象可能是由“光抑制”引起的。
在图1C、1D中,大气二氧化碳浓度变化比较平稳,净光合速率变化与之无规律性相关。与空气相对湿度的日变化曲线也无规律性相关。
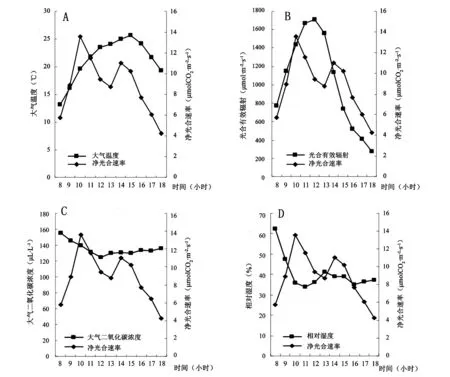
图1 叶片净光合速率与气温、光合有效辐射、大气二氧化碳浓度、相对湿度日变化图
2.2 红富士苹果叶片净光合速率与生理因子的日变化
在图2A、图2B中,红富士苹果叶片净光合速率的日变化与蒸腾速率和气孔导度的日变化均呈正相关,且均为双峰型。
图2B中,10~13时,气孔导度与净光合速率均呈下降态,而图2C中,此时段细胞间隙CO2浓度却略有上升,表明细胞间隙CO2浓度Ci不是导致净光合速率下降的直接原因。图2C中,10时前,净光合速率随细胞间隙CO2浓度Ci的降低而增强,认为净光合速率的变化主要是由气孔因素控制的。经过10~13时的叶片光合“午休”之后,净光合速率随着叶片蒸腾速率和气孔导度的上升再次小幅度冲高,且保持了较短时间。15时后,净光合速率随着各项有利因素的下降而下降,直至日落。
3 小结与讨论
植物光合作用的日变化有单峰型、双峰型、不规则型3种类型,因植物种类、品种、叶位、天气、栽培条件等而异[2]。单峰曲线多出现在阴天[3,4]和没有酷暑的天气条件下[5,6]。果树的净光合速率日变化多呈双峰型[7],本研究的红富士苹果呈双峰型,但还有部分地区的红富士为单峰型[8]。目前尚不能得到关于红富士苹果叶片净光合速率日变化较为统一的结论。
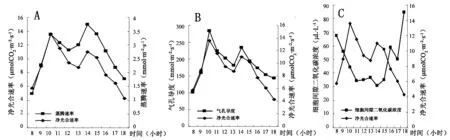
图2 叶片净光合速率与蒸腾速率、气孔导度、细胞间隙二氧化碳浓度日变化图
本研究发现,春季红富士苹果叶片充分展开成熟后,测定其净光合速率日变化呈双峰曲线,10时左右升至高峰值,13时左右降至低谷值,14时左右出现第2个小高峰值,持续一段时间快速降至最低。影响叶片净光合速率的主要生态因子是光合有效辐射、大气CO2浓度,主要生理因子是气孔导度和蒸腾速率。所以红富士生产管理上应保持通风透光良好,合理整形修剪,夏季气温过高、光照太强时,适当遮阴、喷雾或浇水等,减少由于气温过高和光照过强对叶片光合的影响。
本试验研究的结论与孙霞[9]、马慧丽[10]和高照全[11]等的相似,但各地环境不同而研究结果可能略有不同。
对植物光合作用造成影响的各种内外因子多,不同果树对相同因子的敏感度可能不太一致,甚至可能相悖,且各因子也会相互作用,增加了研究的复杂程度。另外,光合作用外界因素多变,有些研究只对一个或几个因素进行简单试验和分析,而对不同果树不同品种的试验,结果可能不同,所以不同地区的多因素试验模拟和试验结果的综合分析还有待进一步的加强和完善。在栽培管理过程中,要综合考虑各因子对净光合速率所产生的直接或间接影响,以及各因子之间的相互关系[12]。
[1] 赵同生,张新生,陈冬梅,付友.苹果光合作用研究进展[J].河北农业科学,2010,14(7):7-10.
[2] 彭永宏,章文才.猕猴桃的光合作用[J].园艺学报,1994,21(2):151-157.
[3] 廖镜思,刘殊,陈清西,等.龙眼光合特性及影响因子的研究[J].园艺学报,1996,23(1):1-7.
[4] 彭方仁,黄宝龙.密植板栗树光合作用研究[J].浙江林学院学报,1997,24(2):151-154.
[5] 温商霖,刘英军.葡萄田间光合作用的研究[J].园艺学报,1989,16(3):168-171.
[6] 王红霞,张志华,玄立春.果树光合作用研究进展[J].河北农业大学学报2003,26(5):49-52.
[7] 邓明净,孙振委,张洪燕,张义奇,王秋涛,刘金川.红富士苹果盆景叶片净光合速率的研究[J].现代农业科技,2007,6(13):15-17.
[8] 郭献平,王燕凌,廖康,郑海燕,杨文君.水分胁迫对新疆野苹果净光合速率和水分利用率日变化的影响[J].新疆农业大学学报,2009.03.
[9] 孙霞,柴仲平,蒋平安.水、氮对南疆'红富士'苹果光合特性日变化的影响[J].北方果树,2010,(1): 3-6.
[10] 马慧丽,吕德国,秦嗣军,刘国成.寒富苹果叶片发育过程中光合特性的研究[J].安徽农业科学,2007, 35(14):4197-4206.
[11] 高照全,李天红,冯社章,张显川.苹果叶片的净光合速率和光能利用效率的动态模拟[J].植物生理学通讯,2010,5:487-492.
[12] 王继和,张明,王春荣.金冠,毛里斯.新红星苹果光合特性的研究[J].西北植物学报,2000,(05): 802-811.
