长鳍裸颊鲷精巢发育的组织学观察
2017-12-18曹伏君章俊杰
曹伏君,叶 宁,章俊杰
( 广东海洋大学,湛江市海洋生态与养殖环境重点实验室, 广东 湛江 524025 )
长鳍裸颊鲷精巢发育的组织学观察
曹伏君,叶 宁,章俊杰
( 广东海洋大学,湛江市海洋生态与养殖环境重点实验室, 广东 湛江 524025 )
以2月龄至2龄的长鳍裸颊鲷精巢为材料,采用常规石蜡切片方法,光镜下观察长鳍裸颊鲷精巢发生、发育至成熟的过程。结果表明,长鳍裸颊鲷的精巢为叶型,位于腹腔后上方,尾部没有合并。精巢发育分为6个时期。精细胞的发育过程也分为6个时期。精细胞的发育由同步到不同步,最后又同步的变化规律。为长鳍裸颊鲷分批排精提供了保障。
长鳍裸颊鲷;精巢;组织学
长鳍裸颊鲷(Lethrinuserythropterus)又名红鳍龙占,隶属于鲈形目、鲈亚目、裸颊鲷科、裸颊鲷属[1],分布于印度—西太平洋区,我国南海亦有分布。其体长可达到50 cm以上,主要以软体动物、甲壳类及小鱼为食,食性较广。长鳍裸颊鲷肉质细嫩,味道鲜美,且抗病力强,是一种优良的养殖种类。目前在海南、粤西等地已经成功地解决了人工繁殖问题,已进入大面积的网箱养殖阶段[2]。关于长鳍裸颊鲷的生物学研究目前尚未见相关报道。本研究通过观察长鳍裸颊鲷精巢的性腺发育,了解其发育规律,为长鳍裸颊鲷的生物学和人工繁殖提供基础资料。
1 材料与方法
1.1 材料
试验期为2012年4月—2014年4月。样品鱼逐月采自海南省海港良种场和广东省湛江东海岛德海养殖场。共采集199 尾,其中成鱼46 尾,幼鱼153 尾,体质量6.7~39.6 kg,鱼龄为2个月至2年。
1.2 方法
活体取出整个精巢,将精巢样本切成小块(幼鱼取整个精巢)后,波恩氏液固定,系列酒精脱水,二甲苯透明,石蜡包埋。连续切片,苏木精—伊红染色。Olympus显微镜镜检,并进行显微摄影[3]。
2 结 果
2.1 精巢形态的发育及显微观察
2.1.1 Ⅰ期精巢
Ⅰ期精巢为初产鱼特有,外观犹如细线状,边缘光滑,半透明,成对存在但在后端并未合并(图1a)。精原细胞数量很少,且排列较为分散,呈圆形或者椭圆形,核占整个细胞的比例较大,染色后核与胞质均呈弱嗜碱性。
2.1.2 Ⅱ期精巢
精巢稍微增大,边缘波浪形,灰白色。精巢约占腹腔1/10的体积,位于腹腔后部,性腺指数(0.06±0.01)%(图1b)。此期精巢有两种情况,一种是繁殖萎缩之后,再待发育的个体;另一种是幼鱼初次发育开始进入生殖周期的个体。可较为明显地观察到由结缔组织间隔形成的生精小管,但无囊腔。组织切片观察可见,精原细胞数目明显增多,在生精小管中,多个精原细胞被结缔组织包裹在一起,排列成束。
2.1.3 Ⅲ期精巢
精巢呈扁片状,同样为灰白色,但透明度降低,可明显看到血丝(图1c)。精巢占腹腔约1/3的体积,位于腹腔后部靠上,性腺指数(0.10±0.04)%。此时的生殖细胞由精原细胞分裂为精母细胞的现象最为活跃,因而数量进一步增加。精细小管是由精原细胞和由其分化而形成的初级精母细胞共同构成,并且在小管中央开始出现空腔。此时可以观察到精巢细胞的发育开始出现非同步性。
2.1.4 Ⅳ期精巢
精巢呈乳白色,精巢占腹腔1/2以上的体积,性腺指数(0.55±0.03)%(图1d)。此时精细小管腔内壁的细胞非同步性表现得更为明显,由精原细胞、初级精母细胞、次级精母细胞及精子细胞组成。在一个精巢断面内,这些发育不同阶段的细胞形成一个圆型的细胞群,一圈圈由外至内,从精原细胞向精子细胞逐层变态发育。其中初级精母细胞最大,次级精母细胞的体型较小,但染色较深,精子细胞最小,染色最深。而此期的生精小管逐渐退化消失。
2.1.5 Ⅴ期精巢
Ⅴ期的精巢更加膨大,较软,乳白色,可以发育到占据腹腔2/3以上的体积,性腺指数(3.58±0.02)%(图1e)。此时细胞数量达到最大值,精小囊消失或融合相通,在精细小管腔和壶腹腔中聚集着大量成熟的精子,经过苏木精—伊红染色后着色最深,可以清楚看见被染成浅红色的流线形尾巴,而精子的圆形头部被染成蓝色。
2.1.6 Ⅵ期精巢
经过排出精子后的精巢体积萎缩,因萎缩形成的空腔由脂肪填充,性腺指数(0.03±0.02)%(图1f)。而精巢内部因精子排出而留下许多空腔,组成精囊壁的细胞主要是精原细胞和初级精母细胞,囊腔中仍残留有少数衰老退化的精子以及精子细胞变态过程中脱落的细胞器,此时精巢的组织结构开始慢慢恢复成为第Ⅱ期精巢的状态。
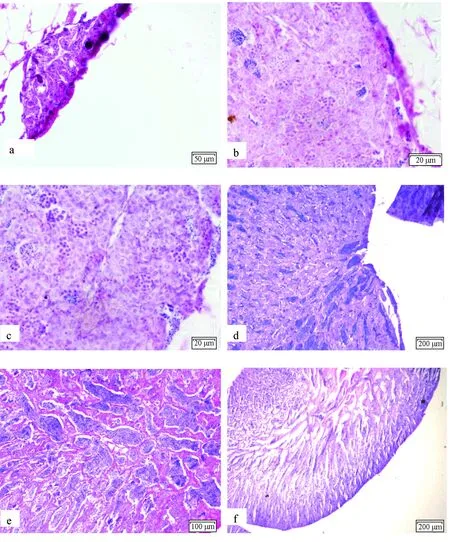
图1 精巢形态的发育的显微观察a.Ⅰ期精巢,×40; b. Ⅱ期精巢,×40; c. Ⅲ期精巢,×40; d. Ⅳ期精巢,×4; e.Ⅴ期精巢,×4;f. Ⅵ期精巢,×4.
2.2 精子发生过程
2.2.1 精原细胞增殖期
精原细胞体积最大,核呈椭圆形,核膜较为清晰,一般具有一个大而圆的核仁,细胞质内有丰富的膜层结构等。根据功能及形态上的差异,将精原细胞分为两类:
第一类精原细胞是原始的胚胎干细胞(图2a)。它只能一直进行有丝分裂,而不发生减数分裂,主要是为了维持精巢内的精原细胞数量,较为明显的特征是核呈卵圆形,中位,染色质均匀分散;
第二类精原细胞则是能够复制分裂之后成为初级精母细胞(图2b)。细胞核呈椭圆型,靠近细胞的一端,染色质相对浓缩,不均匀。这类精原细胞多次进行分裂(图2c,d)。
2.2.2 精母细胞生长期
精原细胞分裂形成初级精母细胞(图2e,f)。早期的初级精母细胞呈椭圆形,体积为精原细胞的一半,细胞核形状不规则,核膜较清晰。生长期的初级精母细胞核质致密,细胞器增加。接近第一次成熟分裂的初级精母细胞核膜不如早期清晰,染色体粗丝状,并进行配对和交换。
2.2.3 精母细胞成熟期
初级精母细胞经历第一次减数分裂后产生2个次级精母细胞(图2g,h) ,此时细胞形状仍保持原有的椭圆形,可观察到细胞器大多仍集中在细胞的一端。之后,次级精母细胞则在很短的时间内进行第二次减数分裂,从而产生2个精子细胞。
2.2.4 精子细胞变态期
精子细胞较小,椭圆形,核质远比次级精母细胞致密,核膜极不清晰(图2i,j)。精子细胞经过一系列形态变化,发育为成熟的精子。初期的精子细胞外形非常不规则,核膜同样不清晰,核内染色质则逐渐聚合成短条状。最后,呈椭圆形,鞭毛继续保持着向后延长,形成精子。
2.2.5 精子成熟期
精子呈圆锥状,分头、中片、尾三部分(图2k,l)。头部大部分被细胞核占据。
3 讨 论
3.1 精巢发育分期
要研究鱼类性腺发育和成熟的一般规律,并且了解其与其他外部因素的关系,则必须依靠划分性腺成熟阶段,从而鉴别鱼的成熟度。我国学者一般采用 Meueh[4]原则,即将硬骨鱼性腺发育分为6个阶段。而根据长鳍裸颊鲷精巢的组织学、细胞学特征,即通过观察其外观变化、精巢内精细胞的发育程度及数量,精细胞的发育过程和精巢发育过程不同,其精巢发育也为6个阶段,与其他大多数鱼类相似。
3.2 精巢结构类型
根据精巢结构以及生精细胞在精巢内分布的特点,通常将硬骨鱼的精巢结构类型分为2类:小管型和小叶型[5-6]。而叶型精巢的主要特点是:精原细胞存在于小叶的边缘处,在发育分化过程中,精小囊逐渐向中央的管腔部位移动,当生殖细胞发育为成熟精子后,精小囊破裂精子释放,进入小叶腔内再流入输出管中。因此,成熟精子主要在小叶的中央部位和输出管内。
长鳍裸颊鲷的精巢与大多数硬骨鱼类的精巢较为相似,呈现为叶型,即其精巢里的精原细胞主要存在于精小叶的边缘,精小叶呈辐射状排列,生精细胞在发育过程中原位发育,并未向中央部位迁移,每个精小叶都是直接与输出管相连。而当发育及变态成为成熟精子之后,精小囊将会自行破裂,使精子释放至小叶腔内,在繁殖季节中,由于充满精子的精小叶间相邻的壁破裂,许多小叶腔从而相互连通成精子通道,并且在最靠近输出管的小叶腔壁开口,使成熟精子能顺利由此进入输出管。这与一般认为的鲈形目特有的管型精巢[7],如金钱鱼(Scatophagusargus)[8]有所不同。但也有其他鲈形目鱼类精巢为小叶型,如中华乌塘鳢(Bostrichthyssinensis)[9]。因此可以认为,鲈形目中鱼类可能存在有多种精巢类型。一般的硬骨鱼类其精巢一般在尾端合并,然后开口于尿殖腔,呈“Y”型[10]。而长鳍裸颊鲷的精巢虽同等大小成对地存在于腹腔后上部,但其后端并未合并连结,而是分别开口通向尿殖腔。
3.3 精子发育特点
硬骨鱼类的精巢结构和精子发生在国内已有诸多报道[11-15]。硬骨鱼类在精子发育过程中,生殖细胞经历了3个主要阶段[16]:由精原细胞分化为精母细胞,精母细胞经过两次减数分裂成精子细胞,精子细胞通过形态变化转变为精子。从组织学切片上可以明显看到,长鳍裸颊鲷的生殖细胞随着发育的不断成熟,同样经历了这3个阶段。长鳍裸颊鲷的初级精原细胞与一般硬骨鱼类[17-18]相似,是精巢中的生精干细胞,由其分裂形成以后各期的生精细胞。
在长鳍裸颊鲷精巢中,精细胞的发育出现了由同步到不同步,最后又同步的分批现象,即第Ⅰ期为单一时相的精原细胞,表现为同步发育;而到了第Ⅱ期则以初级精母细胞为主,第Ⅲ期是以次级精母细胞为主,第Ⅳ期以精子细胞为主,但第Ⅱ、Ⅲ期却仍有不同量的其他时相的精细胞存在,这表现出其精细胞发育的非同步性,非同步性在第Ⅲ期精巢中表现最为明显,精原细胞、初级精母细胞、次级精母细胞3种细胞同时存在;但到了第Ⅳ期后期各时相精细胞反而又表现出一种发育中的追赶现象,待到第Ⅴ期时全部发育为成熟的精子,此时又表现出发育的同步性。这种现象为长鳍裸颊鲷分批排精提供了保障。这种现象在其他鱼类精子发育中也常常出现[19-20]。由于到最后已同步,所以精子同时产出。排精后精巢明显萎缩,未排出的精子以及脱落的细胞质很快退化并被吸收,进入第Ⅵ期后,生殖上皮可明显看到血管变粗,精巢开始充血补充营养并且增厚,开始生殖细胞的又一次发生和形成过程。翌年再形成一个新的繁殖周期。

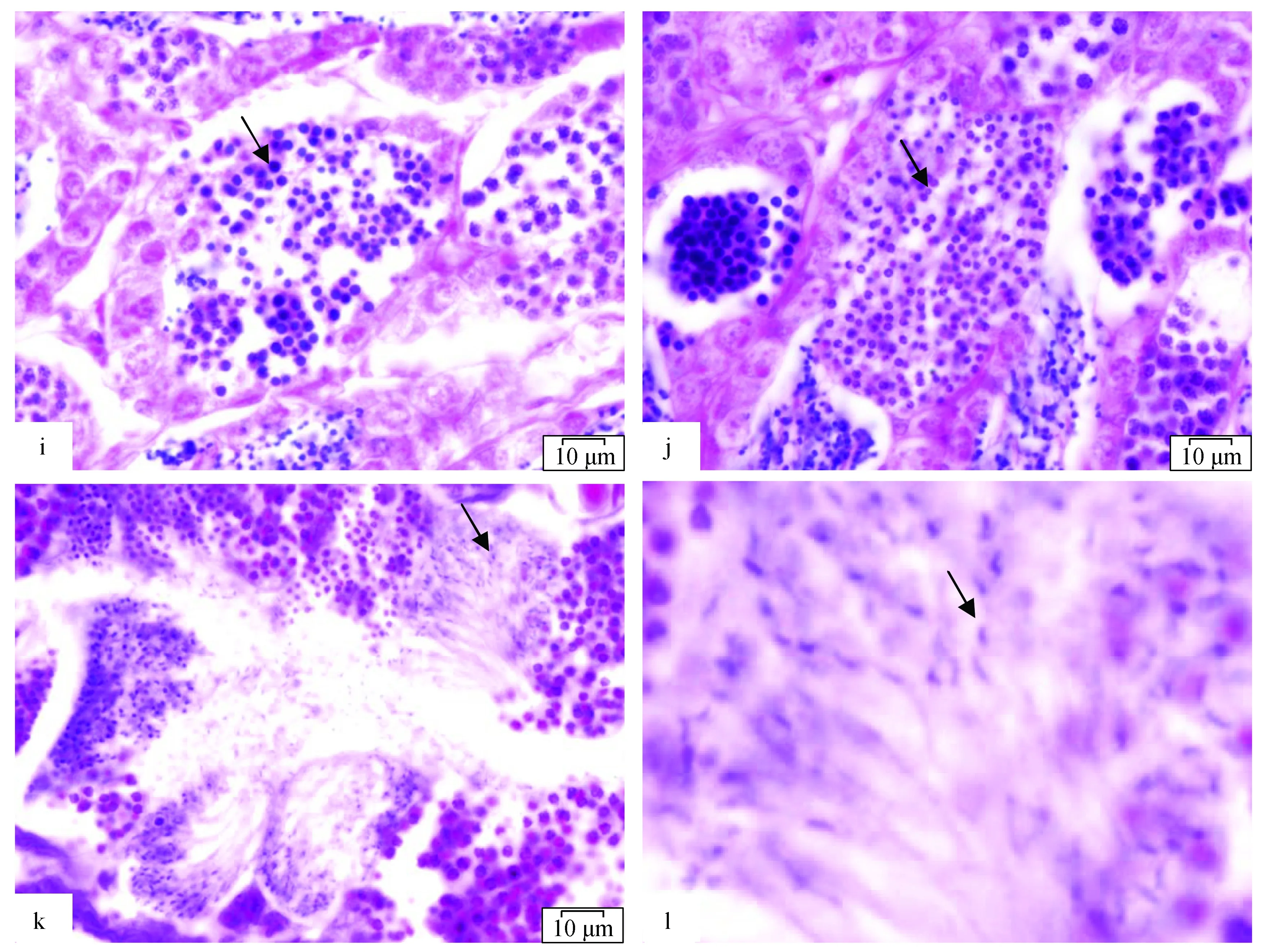
图2 精子发生过程显微观察
a.箭头示附于膜上的精原细胞,×400; b. 箭头示精原细胞,×400; c. 箭头示精原细胞增殖期,×100; d. c图的放大,×200; e. 箭头示初级精母细胞,×100; f. 箭头示初级精母细胞分裂,×400; g. 箭头示次级精母细胞,×100; h. g图的放大,×400; i. 箭头示精子细胞早期,×100; j. 箭头示精子细胞,×100; k. 箭头示精子成熟期,×100; l. k图的放大,×400.
[1] 李明德, 张韶季, 张玉珍. 中国鱼类名录X——鲈亚目(部分)[J]. 海洋通报, 2011, 22(3):45-54.
[2] 陈武文,叶宁,谭保财. 长鳍裸颊鲷全人工繁育和健康养殖技术[J]. 当代水产, 2013(11):76-77.
[3] 刘文彬, 张轩杰. 黄颡鱼精巢发育和周年变化及精子的发生与形成[J]. 湖南师范大学自然科学学报, 2004, 27(1):66-70.
[4] 杨家云. 瓦氏黄颡鱼精巢发育及精子生物学研究[J]. 西南师范大学学报:自然科学版, 2005, 30(4):719-724.
[5] Grier H J, Linton J R, Leatherland J F, et al . Structural evidence for two different testicular types in teleost fishes[J]. Amer J Anat, 1980(159):331-345.
[6] Slveira H P, Azevedo C. Fine structure of the spermatogenesis ofBlenniuspholis(Pisces, Blenniidae) [J] J Submicros Cytol Pathol, 1990, 22 (1):103-108.
[7] 施瑔芳. 鱼类生理学[M]. 北京:农业出版社, 1991:290-291.
[8] 霍丹, 刘志伟, 刘南希,等. 金钱鱼性腺发育及其组织结构观察[J]. 水产学报, 2013, 37(5):796-704.
[9] 江寰新, 尤永隆, 林丹军,等. 中华乌塘鳢鱼精巢的形态结构观察[J]. 福建农林大学学报:自然科学版, 2004, 33(1):89-93.
[10] 温茹淑, 曾德莉, 方展强. 剑尾鱼精巢的显微和超微结构观察[J]. 华南师范大学学报, 2012, 44(4):113-116.
[11] 田照辉,胡红霞,白燕,等.水泥池养殖香鱼性腺发育的观察[J]. 水产科学,2007,26(11):593-596.
[12] 章龙珍,江琪,庄平,等.长鳍篮子鱼繁殖季节性腺的组织学研究[J]. 水产科学,2009,31(2):113-119.
[13] 林君卓,翁幼竹,方永强,等.鲻鱼精子发生的组织学研究[J]. 台湾海峡,2001,20(1):57-60,136.
[14] 王吉桥, 史建国, 姜玉声,等.鸭绿沙塘鳢繁殖习性的观察及性腺发育周期的组织学研究[J]. 水产科学,2008,27(8):379-385.
[15] 陈金涛,谢少林,王超,等.长臀性腺发育及其组织结构观察[J]. 水产科学,2015,34(1):48-52.
[16] Schulz R W, Miura T. Spermatogenesis and its endocrine regulation [J]. Fish Physiol Biochem, 2002(26):43-56.
[17] 林丹军,尤永隆,苏敏.黑脊倒刺鲃精巢结构和精子发生的研究[J].水生生物学报,2003,27(6):564-571.
[18] 刘筠.中国养殖鱼类繁殖生理学[M].北京:农业出版社,1993:25-56.
[19] 张耀光,谢小军.南方鲇的繁殖生物学研究:性腺发育及周年变化[J]. 水生生物学报, 1996,20(1):8-17.
[20] 章龙珍,陈丽慧,庄平,等.长江口纹缟虾虎鱼繁殖季节的性腺组织学[J]. 水产学报, 2009,33(2):245-252.
ObservationonMicrostructureofTestisinLethrinuserythropterus
CAO Fujun, YE Ning, ZHANG Junjie
( Fisheries College, Guangdong Ocean University, Zhanjiang 524025, China )
In this experiment, gonadogenesis and development was histologically observed in testis ofLethrinuserythropterusfrom 2 months old to 2 years old under a light microscope. The results showed that the testis was leaf in shape and located in the abdominal cavity, without connection in caudal portion. Testicular development is divided into six stages:Ⅰ, testis was specific and looked like a thin line;Ⅱ, testis increased slightly with wavy edges; Ⅲ, testis was flat, whose blood vessels were observed without connection in caudal portion;Ⅳ, testis occupied more than 1/2 in abdominal cavity; Ⅴ, testis was milky white in color, more than 2/3 of abdominal cavity; the volume of Ⅵ, testis atrophied. The entire process of sperm cell development ofL.erythropteruswas broadly divided into six periods: multiplication of spermatogonial cells,growth of spermatocytes,maturation of spermatocytes,emergence of spermatoblasts,maturation of spermatozoon, and degeneration of spermatozoon.During the development of sperm, phenomenon appeared in batches from the synchronization to unsynchronization and then the synchronization, which makes the fish in batches ejaculation provide more chances for fertilization.
Lethrinuserythropterus;spermary;histology
10.16378/j.cnki.1003-1111.2017.01.009
Q959.4
A
1003-1111(2017)01-0054-06
2015-12-31;
2016-04-25.
广东省科技计划项目(2012A020602048);广东海洋经济创新发展区域示范项目(粤财农[2013]176号);广东省普通高校省级重大科研项目(GDOU2013050219);湛江市财政资金科技专项(2015A06006).
曹伏君(1967—),女,教授,硕士;研究方向:水产养殖基础生物学.E-mail:1994516@163.com.通讯作者:叶宁(1972—),男,教授级高级工程师,硕士;研究方向:鱼类学、水产养殖学.E-mail:2590713@163. com.
