汉滩病毒包膜糖蛋白嵌合DNA疫苗的细胞免疫评价及其优势表位的筛选
2017-11-13张锦鹏张冠文宣国云郑卓驹欧阳澍毓姜东伯
张锦鹏,张冠文,宣国云,孟 强,张 哲,郑卓驹,欧阳澍毓,张 正,姜东伯,杨 琨
(第四军医大学:1基础部免疫学教研室,2学员旅,陕西西安710032)
汉滩病毒包膜糖蛋白嵌合DNA疫苗的细胞免疫评价及其优势表位的筛选
张锦鹏1,2,张冠文1,2,宣国云1,2,孟 强1,2,张 哲1,2,郑卓驹1,2,欧阳澍毓1,2,张 正1,2,姜东伯1,杨 琨1
(第四军医大学:1基础部免疫学教研室,2学员旅,陕西西安710032)
目的:筛选Gn和Gc抗原优势表位,为汉滩病毒疫苗研究提供方向.方法:通过构建汉坦病毒包膜糖蛋白Gn与Gc嵌合溶酶体相关膜蛋白的DNA疫苗pVAX⁃LAMP/Gn,pVAX⁃LAMP/Gc,利用酶联免疫斑点实验(ELISpot)长期评估了BALB/c小鼠体内的细胞免疫应答.结果:在刺激T淋巴细胞的短效与长效免疫中,pVAX⁃LAMP/Gc组的效应优于其他组,并筛选出优势抗原表位.结论:目前许多新型汉滩病毒疫苗研究缺少细胞免疫应答评价和记忆免疫应答评价.筛选出的Gn和Gc的抗原优势表位,可以为新型汉滩病毒疫苗的研制奠定良好的基础.
汉滩病毒包膜蛋白;LAMP分子;DNA疫苗;表位筛选;免疫学评价
0 引言
汉滩病毒(Hantaan virus,HTNV)是布尼亚病毒科汉坦病毒属的一种有包膜分节段单股负性RNA病毒,主要通过接触啮齿类动物的分泌物传播,导致肾综合征出血热(hemorrhagic fever with renal syndrome,HFRS)[1],亦称流行性出血热.HFRS是一种以发热、疼痛(头痛、腰痛、眼眶痛)、出血、急性肾功能损伤为主要临床表现的综合征.在自然感染过程中,根据病毒CBI等类型,病死率可以从2%变化到15%.在辅助治疗如连续性肾脏替代治疗(continuous renal replacement therapy,CRRT)中,在感染的早期阶段,病死率控制在1%以内.每年全球约有10万人发病,我国是世界上发病最为严重的国家,占总发病人数的90%以上,发病人数多且流行范围广,严重威胁疫区人民的生命健康与安全[2-3].而目前临床上尚无针对HFRS的特效药物,疫苗目前被认为是控制该疾病最为行之有效的办法.我国临床上使用的是双价灭活疫苗,可以产生具有保护作用的中和抗体[4],然而该疫苗在诱导机体产生细胞免疫应答及产生长期免疫记忆能力方面存在不足,大大降低了其在公共卫生安全领域中的推广价值.第四军医大学基础部免疫学教研室长期从事DNA疫苗的研制.DNA疫苗因其能够模拟病原体自然感染激活体内免疫应答,在生产、储存、运输方面的优势使其成为疫苗研制领域中的热点.
汉滩病毒基因组分为大(L)、中(M)、小(S)三个片段,分别编码病毒的RNA聚合酶、包膜糖蛋白(Gn和Gc)和核衣壳蛋白(nucleocapsid protein,NP).其中M片段全长仅有一个开放阅读框,编码包膜糖蛋白复合体,该前体蛋白在细胞内质网被切割加工成Gn与Gc两个糖蛋白[5].Gn与Gc构成病毒吸附蛋白,可识别结合细胞表面病毒受体,在病毒对机体细胞的侵袭与感染过程中发挥重要作用[6].相关文献报道,Gn、Gc上均有中和抗原位点,人自然感染汉滩病毒时,可诱导机体产生特异性免疫应答,刺激机体生成一定量保护性的中和抗体[7-8].中和抗体在清除体内病毒这一过程中扮演重要角色,提示汉滩病毒的Gn与Gc在建立机体对抗病毒感染发病的特异性免疫保护机制中发挥重要作用,可作为DNA疫苗的研究靶点.然而,传统基因疫苗表达内源性抗原,通过MHCⅠ类抗原加工提成途径激活CD8+T细胞,诱导机体产生有效的细胞免疫应答,而CD4+T细胞得不到有效活化.
溶酶体相关膜蛋白(lysosome⁃associated mem⁃brane protein,LAMP)是位于溶酶体膜上的Ⅰ型穿膜蛋白,由N端溶酶体内腔部分、疏水穿膜部分及C末端短胞浆尾构成[9].LAMP分子合成后,其保守的短胞浆尾通过穿膜⁃胞浆区定向地结合亚细胞结构——内吞体/溶酶体,主要发挥靶向转运至溶酶体的重要功能.当编码目的抗原的基因插入到LAMP分子编码溶酶体内腔部分和胞浆尾的基因之间构成嵌合蛋白时,利用LAMP分子胞浆尾靶向至内体/溶酶体膜作用,可将LAMP融合蛋白中目的抗原直接携带进入主要组织相容性复合体(major histocompatibility complex,MHC)Ⅱ类分子器室(MHC classⅡcom⁃partment,MIIC),从而可实现DNA疫苗所表达蛋白从内源性抗原的MHC⁃Ⅰ类加工途径向外源性抗原的MHC⁃Ⅱ类加工提呈途径的转化,有效地激活了CD4+T细胞[10-11].活化的CD4+T既可以分泌大量细胞因子,刺激其他免疫细胞的活化、增殖、分化[12],又能表达CD40L,作为第二信号辅助B细胞活化分泌抗体,从而增强适应性免疫应答以及记忆T细胞和B细胞的发生.
本研究利用汉滩病毒包膜糖蛋白Gn和Gc联合LAMP靶向作用,构建编码LAMP和HTNV包膜糖蛋白Gn与Gc的新型重组嵌合表达载体pVAX⁃LAMP/Gn和pVAX⁃LAMP/Gc,分别在短时间内连续3次免疫接种后和半年加强免疫后评估免疫接种小鼠细胞免疫应答和长效的免疫记忆能力,为进一步研究新型出血热基因疫苗奠定基础.
1 材料和方法
1.1 材料重组质粒为本实验室前期构建保存,含有LAMP分子全长序列的P43⁃LAMP载体为美国约翰·霍普金斯大学馈赠.pVAX 1真核表达载体、LipofectamineTM2000购自美国Invitrogen公司.去内毒素质粒大提试剂盒购自北京天根生物有限公司,新生牛血清购自杭州四季青责任有限公司.细胞培养基RPMI⁃1640为本室配制.三级实验动物成年雌性BALB/c小鼠购自第四军医大学实验动物中心.双价肾综合征出血热灭活疫苗由长春生物制品研究所提供.ELISpot试剂盒(美国BD公司),ELISpot读数仪(美国Cellular Technology Ltd.).
1.2 方法
1.2.1 重组载体的构建及鉴定 前期实验中已对重组表达载体的构建及鉴定作了详细的介绍,在此不作过多论述.并且转染实验显示目的基因能够在真核细胞中稳定表达.
1.2.2 重组基因疫苗的免疫方案 140只小鼠(SPF级成年雌性BALB/c小鼠)分7组(pVAX组、pVAX⁃LAMP组、pVAX⁃Gn组、pVAX⁃LAMP/Gn组、pVAX⁃Gc组、pVAX⁃LAMP/Gc组和灭活疫苗组),每组20只.重组质粒50 μg/只经尾根部皮下免疫,共免疫3次,每次间隔3周.第3次免疫2周后处死部分小鼠取脾脏进行短效酶联免疫斑点实验(enzyme⁃linked immunospot assay,ELISpot)(BD Ptarmigan IFN⁃γ),半年后加强免疫一次,2周后进行长效免疫评价.
1.2.3 HTNV⁃Gn/Gc多肽合成 在Genebank数据库中查找HTNV76⁃118株糖蛋白氨基酸序列(P08668⁃1,Hantaan virus glycoprotein sequence),并委托上海强耀生物科技有限公司合成重叠多肽,每条多肽长度为15aa,相邻两条肽重叠8aa,共合成Gn 89条(其中6条未合成成功),Gc 68条(其中4条未合成成功).每条15肽合成10 mg,经RP⁃HPLC测定纯度>85%,多肽干粉于-20℃长期保存.所有15肽均先用20 μL无菌DMSO溶解,待完全溶解后再加入无血清PRMI⁃1640不完全培养基配制成终浓度为1 mg/mL的溶液,分装后-70℃冻存备用.
1.2.4 ELISpot 取免疫小鼠脾脏,制备脾细胞悬液进行ELISpot检测.用IFN⁃γ捕获抗体5 mg/L包被96孔PVDF滤膜板,4℃过夜.次日用不含血清RPMI⁃1640洗2遍,后用含10%新生牛血清的PRMI⁃1640培养基封闭,200 μL/孔,室温2 h.调整脾淋巴细胞密度为1×107/mL,100 μL/孔;加入抗原肽(分别将Gn的83条肽或Gc的64条肽平均分为9或8组肽池,先进行肽池实验,筛选出阳性肽池后再做单肽实验),100 μL/孔,3个复孔,同时设ConA(终浓度为5 mg/L)阳性对照和完全培养基阴性对照孔,37℃、5%CO2孵育24 h.洗板、拍干后加入1 mg/L生物素化IFN⁃γ检测抗体,100 μL/孔,室温孵育2 h;洗板拍干后加入HRP⁃链霉亲和素100 μL/孔,室温孵育1 h;加入AEC底物显色,室温避光静置15~45 min,待斑点形成合适的大小后用自来水冲洗终止显色过程,晾干,ELISpot自动读板仪计数,结果以每1×106脾细胞中可分泌细胞因子的斑点形成数(spot forming unite,SFU)表示.
1.3 统计学处理检测数据使用均数±标准差(mean±SD)的呈现方法,采用Graphpad Prism 5.0软件分别进行student⁃t检验、Mann⁃Whitney检验、one⁃way ANOVA分析.
2 结果
2.1 Gc刺激短效免疫小鼠脾淋巴细胞特异性分泌IFN-γ的频率通过ELISpot检测免疫小鼠脾细胞分泌细胞因子发现,在Gc的8个肽池中,2、3、5肽池pVAX⁃LAMP/Gc组明显优于pVAX⁃Gc组和灭活疫苗组(Inactivated Vaccine组)(图1A).随后对2、3、5肽池中单肽进行ELISpot,整体上pVAX⁃LAMP/Gc分泌IFN⁃γ高于pVAX⁃Gc组和灭活疫苗组(图1B),进行统计学分析筛选出阳性肽(图1C).

图1 Gc短效ELISpot检测IFN⁃γ
2.2 Gc刺激长效免疫小鼠脾淋巴细胞特异性分泌IFN-γ的频率Gc长效应答结果显示,pVAX⁃LAMP/Gc组明显增强,IFN⁃γ分泌频率为pVAX⁃Gc组和灭活疫苗组的两倍以上(图2).全肽的单肽实验显示,pVAX⁃LAMP/Gc几乎覆盖Gc全长的表位扩展现象,且记忆回应明显优于pVAX⁃Gc组和灭活疫苗组(图3).
2.3 Gn刺激短效免疫小鼠脾淋巴细胞特异性分泌IFN-γ的频率Gn短效应答结果显示:第6、7、8组有明显的阳性应答,且pVAX⁃LAMP/Gn组分泌IFN⁃γ的频率明显较Gn组更高.将筛选得到的混合肽进一步行单肽实验,得到一条阳性肽(图4).
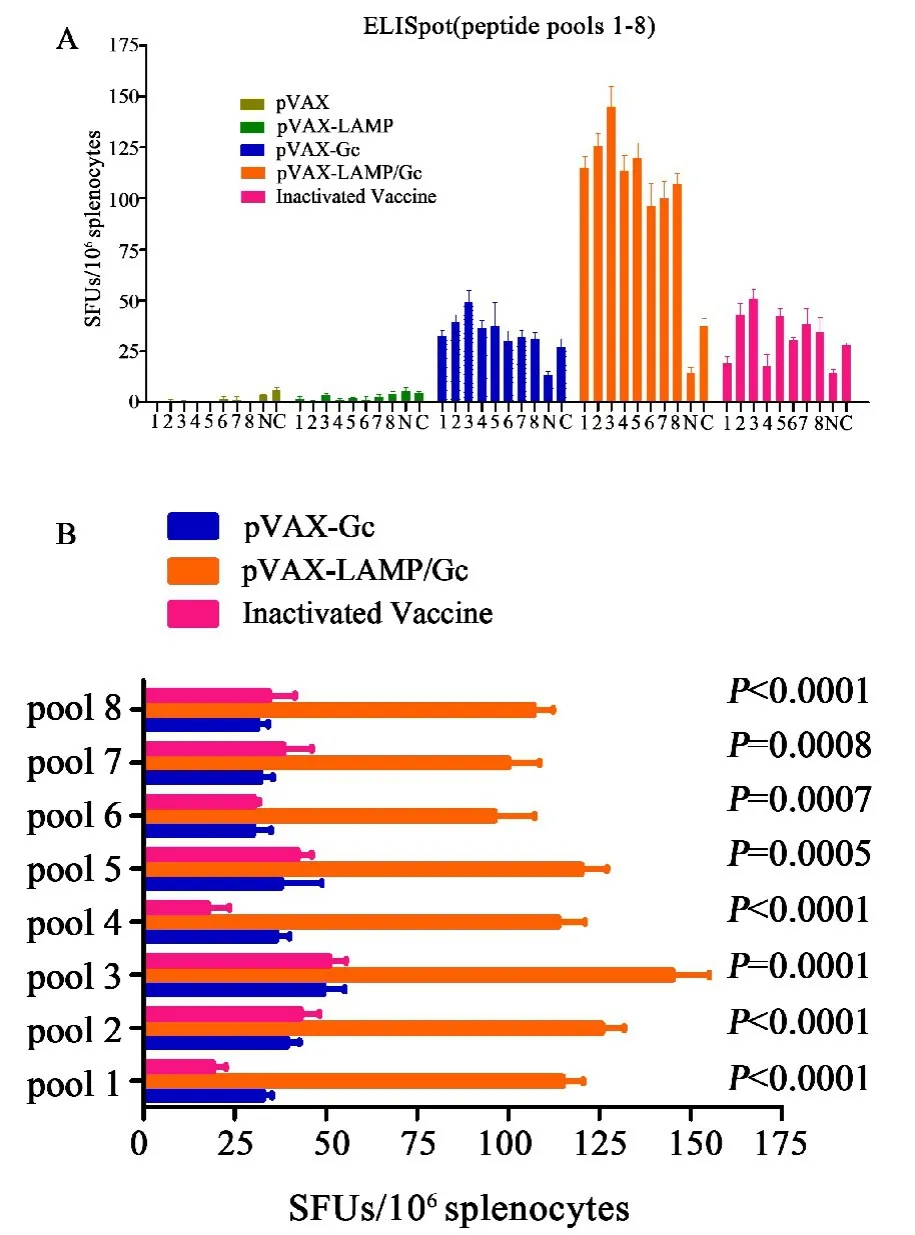
图2 Gc长效肽池ELISpot检测IFN⁃γ
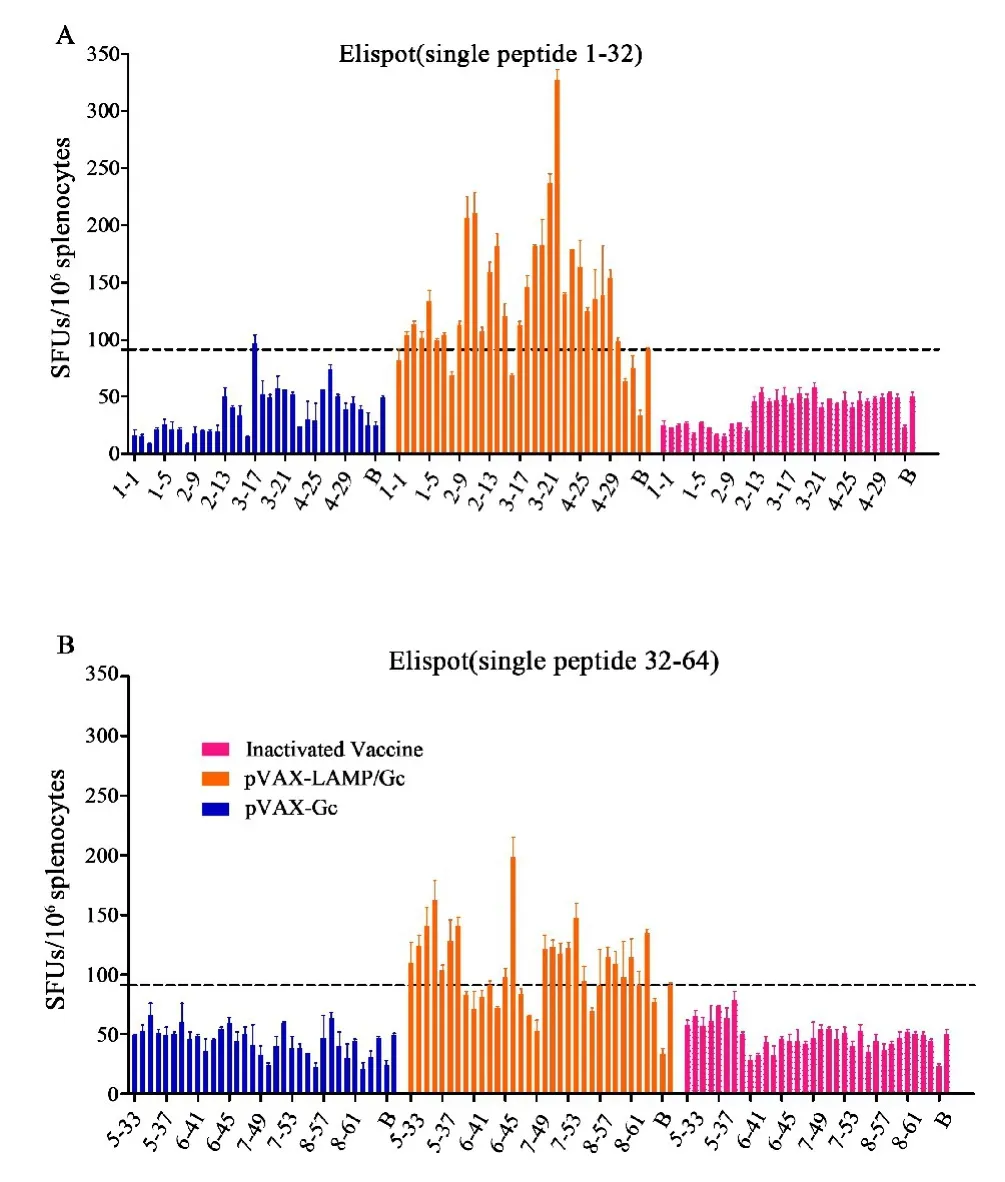
图3 Gc长效单肽ELISpot检测IFN⁃γ

图4 Gn短效ELISpot检测IFN⁃γ
2.4 Gn刺激长效免疫小鼠脾淋巴细胞特异性分泌IFN-γ的频率Gn长效应答结果显示,3、4、6、7和8号肽池均为阳性,单肽实验观察到更为明显的表位扩展现象(图5).
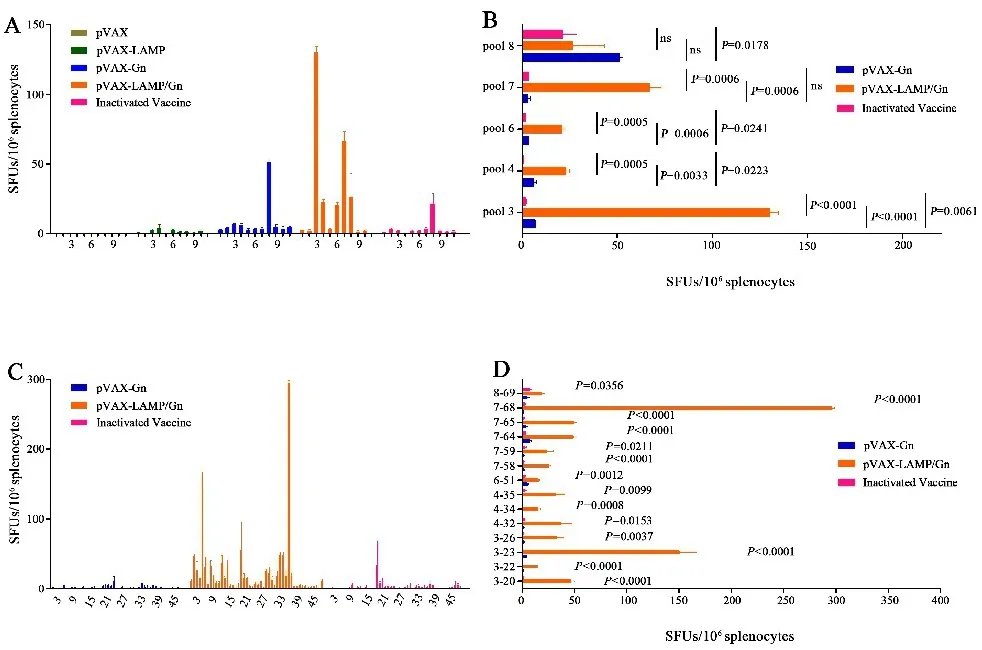
图5 Gn长效ELISpot检测IFN⁃γ
3 讨论
我国是HFRS高发国家,给我国造成了严重的人员伤亡及巨大的经济损失,其疫苗研究有着重要的现实意义.双价灭活疫苗能诱导机体产生一定量的中和抗体,可以在出血热防治方面发挥重要作用,但存在细胞免疫应答不明确和长效免疫记忆能力较差的问题[13].现国内外新型汉滩病毒疫苗研究包括减毒活疫苗、亚单位疫苗、VLP等,其中更集中于DNA疫苗的研制[14-15],但也遇到传统基因疫苗表达内源性抗原,只能通过MHCⅠ类途径活化CD8+T细胞,不能有效活化CD4+T细胞,缺乏长效记忆免疫等问题.
近日,韩国科学家提出记忆性T淋巴细胞可以避免汉滩病毒持续性感染,在预防HFRS复发中具有重要意义.灭活疫苗长期抗体效价低,若需保持一定滴度抗体,需要不断加强免疫,新型疫苗的研究应关注长效免疫记忆能力和细胞免疫评价.实验中进行了汉滩病毒灭活疫苗的长期血清学评价,发现两次初次免疫接种后一年内抗体迅速下降,一年后加强一次免疫,蚀斑减少中和试验的转阳率达40%以上,此后抗体效价逐渐下降[16].研究缺少细胞免疫评价可能与CD8+T在HFRS疾病进展中作用仍存在争议有关,目前认为HFRS主要是由免疫系统介导,比如细胞因子、补体系统和细胞介导的反应.皮损中CTL的募集和浸润可能会侵袭感染的上皮细胞或内皮细胞,造成细胞损伤和漏血.然而研究发现中国中西部汉族人口的HFRS中,CD8+T的升高与疾病的严重程度呈负相关,提示CD8+T在疾病进展的保护作用[17-18].这表明T细胞在HFRS疾病进展的免疫保护与免疫损伤之间有着微妙的平衡,有待进一步研究.
本研究通过分子克隆手段,利用汉滩病毒糖蛋白cDNA成功构建重组真核表达载体pVAX⁃LAMP/Gn、pVAX⁃LAMP/Gc.将目的基因Gn、Gc插入编码LAMP分子溶酶体内腔部分和胞浆尾的基因之间构成嵌合体时,利用LAMP分子的靶向内体/溶酶体作用,可使抗原蛋白直接进入MⅡC,实现MHCⅠ类途径向MHCⅡ类途径转化.这一策略克服了传统DNA疫苗单纯诱导细胞免疫应答的缺陷,实现了抗原加工提呈途径的转换,有效激活CD4+T细胞,增强了体液免疫应答和细胞免疫应答,也获得了良好的长效记忆免疫应答[19-20],为出血热疫苗的研究打下良好的基础.
在免疫评价中,汉滩病毒包膜糖蛋白Gn、Gc显示了很好的免疫原性,可诱导小鼠产生良好的免疫应答.在整体IFN⁃γ释放频率上,Gc优于Gn.这可能与Gn表达量相对较低有关,Gn蛋白尾部存在的WAA⁃SA序列可以激活宿主细胞内部的自噬机制,使得蛋白降解而无法在细胞内稳定存在[21-22].除此之外,Gn上含有5个天冬酰胺连接糖基化位点,而Gc上有2个,这造成Gn糖基化程度高,结构更复杂,表达难度大于Gc.上述原因共同导致Gn表达量低于Gc.
此外本研究对比发现,在ELISpot筛选得到的Balb/c小鼠特异性的优势表位与前期研究筛选出的出血热患者特异的优势表位存在种属间交叉现象[18],其与H2⁃Kd限制性的表位也同样存在交叉[23],这有望在不同实验动物上筛选汉滩病毒的优势表位,寻找多种不同限制性表位间的共同优势表位.这种共同优势表位在表位疫苗研究中有重要意义,可能会增加在不同种族不同人群中应用的广泛性,其意义更有待进一步研究.
[1] Avšcic⁃županc T,Saksida A,Korva M.Hantavirus infections[J].Clin Microbiol Infect,2015.
[2] Mir MA.Hantaviruses[J].Clin Lab Med,2010,30(1):67-91.
[3] Jiang H,Zheng X,Wang L,et al.Hantavirus infection:a global zoo⁃notic challenge[J].Virol Sin,2017,32(1):32-43.
[4] Szabó R.Antiviral therapy and prevention against hantavirus infec⁃tions[J].Acta Virol,2017,61(1):3-12.
[5] Battisti AJ,Chu YK,Chipman PR,et al.Structural studies of Hantaan virus[J].J Virol,2011,85(2):835-841.
[6] Muyangwa M,Martynova EV,Khaiboullina SF,et al.Hantaviral Proteins:Structure,Functions,and Role in Hantavirus Infection[J].Front Microbiol,2015,6:1326.
[7] Hooper JW,Josleyn M,Ballantyne J,et al.A novel Sin Nombre virus DNA vaccine and its inclusion in a candidate pan⁃hantavirus vaccine against hantavirus pulmonary syndrome(HPS)and hemor⁃rhagic fever with renal syndrome(HFRS)[J].Vaccine,2013,31(40):4314-4321.
[8] Higa MM,Petersen J,Hooper J,et al.Efficient production of Hanta⁃an and Puumala pseudovirions for viral tropism and neutralization studies[J].Virology,2012,423(2):134-142.
[9] Marques ET Jr,Chikhlikar P,de Arruda LB,et al.HIV⁃1 p55Gag encoded in the lysosome⁃associated membrane protein⁃1 as a DNA plasmid vaccine chimera is highly expressed,traffics to the major histocompatibility classⅡcompartment,andelicits enhanced immune responses[J].J Biol Chem,2003,278(39):37926-37936.
[10] Anwar A,Chandrasekaran A,Ng ML,et al.West Nile premem⁃brane⁃envelope genetic vaccine encoded as a chimera containing the transmembrane and cytoplasmic domains of a lysosome⁃associated membrane protein:increased cellular concentration of the transgene product,targeting to the MHC II compartment,and enhanced neutralizing antibody response[J].Virology,2005,332(1):66-77.
[11] Chikhlikar P,Barros de Arruda L,Maciel M,et al.DNA encoding an HIV⁃1 Gag/humanlysosome⁃associatedmembraneprotein⁃1 chimera elicits a broad cellular and humoral immune response in Rhesus macaques[J].PLoS One,2006,1:e135.
[12] Nakanishi Y,Lu B,Gerard C,et al.CD8(+)T lymphocyte mobili⁃zation to virus⁃infected tissue requires CD4(+)T⁃cell help[J].Nature,2009,462(7272):510-513.
[13] Hooper JW,Li D.Vaccines against hantaviruses[J].Curr Top Microbiol Immunol,2001,256:171-191.
[14] Boudreau EF,Josleyn M,Ullman D,et al.A Phase 1 clinical trial of Hantaan virus and Puumala virus M⁃segment DNA vaccines for hemorrhagic fever with renal syndrome[J].Vaccine,2012,30(11):1951-1958.
[15] Hooper JW,Moon JE,Paolino KM,et al.A Phase 1 clinical trial of Hantaan virus and Puumala virus M⁃segment DNA vaccines for haemorrhagic fever with renal syndrome delivered by intramuscular electroporation[J].ClinMicrobiol Infect,2014,20Suppl 5:110-117.
[16] Song JY,Woo HJ,Cheong HJ,et al.Long⁃term immunogenicity and safety of inactivated Hantaan virus vaccine(HantavaxTM)in healthy adults[J].Vaccine,2016,34(10):1289-1295.
[17] Ma Y,Wang J,Yuan B,et al.HLA⁃A2 and B35 restricted hantaan virus nucleoprotein CD8+T⁃cell epitope⁃specific immune response correlates with milder disease in hemorrhagic fever with renal syndrome[J].PLoS Negl Trop Dis,2013,7(2):e2076.
[18] Ma Y,Yuan B,Zhuang R,et al.Hantaan virus infection induces both Th1 and ThGranzyme B+cell immune responses that associated with viral control and clinical outcome in humans[J].PLoS Pathog,2015,11(4):e1004788.
[19] Yang K,Sun K,Srinivasan KN,et al.Immune responses to T⁃cell epitopes of SARS CoV⁃N protein are enhanced by N immunization with a chimera of lysosome⁃associated membrane protein[J].Gene Ther,2009,16(11):1353-1362.
[20] Dhalia R,Maciel MJr,Cruz FS,et al.Membrane and envelope virus proteins co⁃expressed as lysosome associated membrane protein(LAMP)fused antigens:a potential tool to develop DNA vaccines against flaviviruses[J].An Acad Bras Cienc,2009,81(4):663-669.
[21] Hussein IT,Cheng E,Ganaie SS,et al.Autophagic clearance of Sin Nombre hantavirus glycoprotein Gn promotes virus replication in cells[J].J Virol,2012,86(14):7520-7529.
[22] Ganaie SS,Mir MA.The role of viral genomic RNA and nucleocapsid protein in the autophagic clearance of hantavirus glycoprotein Gn[J].Virus Res,2014,187:72-76.
[23] Ma RX,Cheng LF,Ying QK,et al.Screening and Identification of an H⁃2Kb⁃Restricted CTL epitope within the glycoprotein of hantaan virus[J].Front Cell Infect Microbiol,2016,6:151.
Cellular immune response evaluation of Hantavirus glycoprotein chimeric DNA vaccine and its screening of dominant epitopes
ZHANG Jin-Peng1,2,ZHANG Guan-Wen1,2,XUAN Guo-Yun1,2,MENG Qiang1,2,ZHANG Zhe1,2,ZHENG Zhuo-Ju1,2,Ou-Yang Shu-Yu1,2,ZHANG Zheng1,2,JIANG Dong-Bo1,YANG Kun11Department of Immunology,2Brigade of Cadet,The Fourth Mili⁃tary Medical University,Xi'an 710032,China
AIM:To screen the dominant epitopes of antigens and provide a direction for further research of Hantaanvirus vaccine.METHODS:DNA vaccine encoding Hantavirus glyco⁃proteins fused with lysosome⁃associated membrane protein was designed in this study.cellular immune response of BALB/c mice was assessed by ELISpot.RESULTS:The pVAX⁃LAMP/Gc group showed great advantages in stimulating short⁃term and long⁃term immunity reaction,and dominant antigen epitopes were found.CONCLUSION:Currently,lots of HFRS vaccine resear⁃ches is limitted in cellular immune response and memory immune response evaluation.Epitope mapping was performed to study the T⁃cell epitopes.This study lays a good foundation for the research of Hantavirus vaccine.
Hantanvirus glycoprotein;LAMP;DNA vaccine;epitope mapping;immune response evaluation
R446.63
A
2017-07-16;接受日期:2017-07-30
国家自然科学基金面上项目(81171977)
张锦鹏.E⁃mail:418110759@qq.com
杨 琨.博士,教授,博导.研究方向:病毒疫苗,肿瘤治疗,生物信息学.E⁃mail:yangkunkun@fmmu.edu.cn姜东伯(共同通讯作者).硕士生.研究方向:病毒疫苗,生物信息学.E⁃mail:superjames1991@foxmail.com
2095⁃6894(2017)10⁃26⁃05
