内蒙古地区布氏田鼠种群MHCⅡ类基因第二外显子的遗传分析
2017-09-26赵素华
赵素华,魏 磊
①
(1.宿州职业技术学院,安徽 宿州 234101;2.毫州学院 生物与化学工程系,安徽 亳州 236800;3.中国科学院 动物研究所动物生态和保护生物学院重点实验室,国家野生动物疫病研究中心,北京 100101)
内蒙古地区布氏田鼠种群MHCⅡ类基因第二外显子的遗传分析
赵素华1,魏 磊2,3
①
(1.宿州职业技术学院,安徽 宿州 234101;2.毫州学院 生物与化学工程系,安徽 亳州 236800;3.中国科学院 动物研究所动物生态和保护生物学院重点实验室,国家野生动物疫病研究中心,北京 100101)
以MHCⅡ类基因第二外显子(MHC classⅡ exon 2)为分子标记,用限制性酶切多态性和直接测序的方法对分布于内蒙古地区的8个布氏田鼠种群共460个个体进行种群遗传结构的分析.结果显示,酶切共检测到6个等位基因,13个酶切多态性位点,卡方检验显示,各种群在6个酶切多态性位点上基因型频率不符合Har⁃dy-Weinberg平衡.序列分析显示,在长度261 bp的核苷酸序列中,有57个变异位点,定义21种单倍型,其中有1个单倍型为不同区域种群所共享,其余20个单倍型均为各区域种群所特有.7个地理种群的单倍型多样性和核苷酸多样性较高,1个地理种群较低.谱系分析显示,8个布氏田鼠地理种群分为3个进化分支,分别与采集的地理种群相吻合:同一地理种群内单倍型之间遗传差异小,而不同地理来源的单倍型之间存在较大区别. Mentel检测表明,布氏田鼠的遗传分化与地理距离呈现正相关.AMOVA分析结果同样表明地理种群之间存在差异:各区域类群间变异组分占总变异比率的60.48%,区域内种群间变异组分占总变异比率的7.78%,种群内个体间变异组分占总变异比率的28.14%;遗传分化系数和基因流分析显示,布氏田鼠种群出现一定程度的分化,但正镶白旗种群与其他种群之间遗传分化显著.布氏田鼠的这种遗传结构特点可能是该物种稳定的地下生活环境和有限的迁移造成的,且浑善达克沙漠形成布氏田鼠分化最强烈的隔离因素.
布氏田鼠;MHCⅡ类基因;遗传分化
0 引言
内蒙古地理环境复杂,独特的地形、地貌和地质历史对物种结构产生较大的影响.布氏田鼠(Lasiopodomys brandtii)是内蒙古草原具有一定代表性的小型哺乳动物,具有扩散能力弱,对环境依赖性强的特点,因此是研究复杂环境对种群遗传特性影响的典型物种代表.Wang等[1]基于微卫星标记对内蒙古4个布氏田鼠地理种群分析表明,栖息地的破碎化、沙漠化、环境隔离使种群间基因交流减少、存在近交繁殖现象,从而影响着布氏田鼠的种群遗传结构.张曼等[2]采用微卫星标记对连续三代布氏田鼠封闭群遗传结构稳定性进行分析,通过平均有效杂合度和多态信息含量测定,该布氏田鼠种群传代过程中具有稳定的杂合度.MHCⅡ类基因包括经典DR、DQ和DP等亚区、非经典DO、DM和DN等亚区,MHCⅡ类基因各经典的亚区间包含有很大的差异,推测其原因是因为不同物种在进化中积累的特异性复制造成的[3].本文尝试以MHCⅡ类基因第二外显子为分子标记,分析布氏田鼠种群的遗传结构特征,及遗传距离与地理距离之间的关系,探讨环境因素对物种进化的影响,具有重要的进化理论研究价值.
1 材料与方法
1.1 样品采集
样品采集来自于正镶白旗(42°25.611′N,115°16.331′E;n=62)、二连浩特(43°53.245′N,112°23.564′E;n=65)、锡林浩特(44°50.351′N,115°13.673′E;n=68)、东乌珠穆沁旗(44°59.454′N,116°10.846′E;n=54)、西乌珠穆沁旗(44°5.211′N,117°57.324′E;n=53)、新巴尔虎右旗(48°21.625′N,118°22.573′E;n=50)、新巴尔虎左旗(49°18.152′N,118°03.776′E;n=56)、陈巴尔虎旗(49°32.652′N,119°44.735′E;n=52),共8个种群460只布氏田鼠,各采样点之间的直线距离均在100 km以上.
1.2 DNA的提取
以Genbank序列(AJ003232,AF084934,U88933,M73960,BC132163,K0012,AJ634270)作为参考,设计一对引物,其中上游引物(MF)为5′-GGAATACGGTACATCTGAG-3′,下游引物(MR)为5′-CAACTCTCCGCCGCACAAGG-3′;PCR反应体系与条件参照文献[4]设置.
1.3 RsaⅠ酶切
按说明书提供的方法,用RsaⅠ(购自宝大连生物工程有限公司)酶切1.2提取的DNA,酶切产物用8%的EB琼脂糖电泳,拍照并观察结果.
1.4 数据分析
对各酶切片段进行基因型频率统计,并作Hardy-Weinberg平衡的卡方适合性检验.运用分析软件DNAstar 5.0中的Editseq、Seqman程序作基因片段拼接,再用ClustalW 1.83排序,然后运用MEGA4软件分析序列特征,计算遗传距离(p-distance),并借用UPGMA法构建种群分化树,参数均采用默认设置.运用GenAlEx计算地理距离与遗传距离之间的相关性(Mantel tests),运用Arlequin 3.1工具计算单倍型多样度(haplotype diversity,h)和核苷酸多样度(nucleotide diversity,π)以及各种群间的遗传分化指标(F-Statis⁃tics(FST)),并作分子变异分析(analysis of molecular variance,AMOVA)和P-value检验分析.
2 结果
2.1 MHCⅡ类基因第二外显子酶切单倍型与等位基因型的频率
在布氏田鼠8个地理种群MHCⅡ类基因第二外显子的RsaⅠ酶切片段中共检测到6个等位基因,13个多态位点,经卡方检验,6个多态性位点上基因型频率不符合Hardy-Weinberg平衡,X2值分别为49.68(P<0.05)、56.31(P<0.05)、66.04(P<0.05)、59.95(P<0.05)、50.52(P<0.05)和48.15(P<0.05).其中,正镶白旗地理种群纯合子基因型频率较高(0.064 5~0.112 9),杂合子基因型频率较低(0.016 2~0.048 4);其他地理种群纯合子基因型频率与杂合子基因型频率基本一致(见表1).
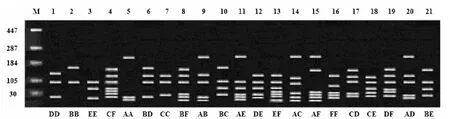
图1 PCR产物RsaⅠ酶切8%琼脂糖电泳检测结果
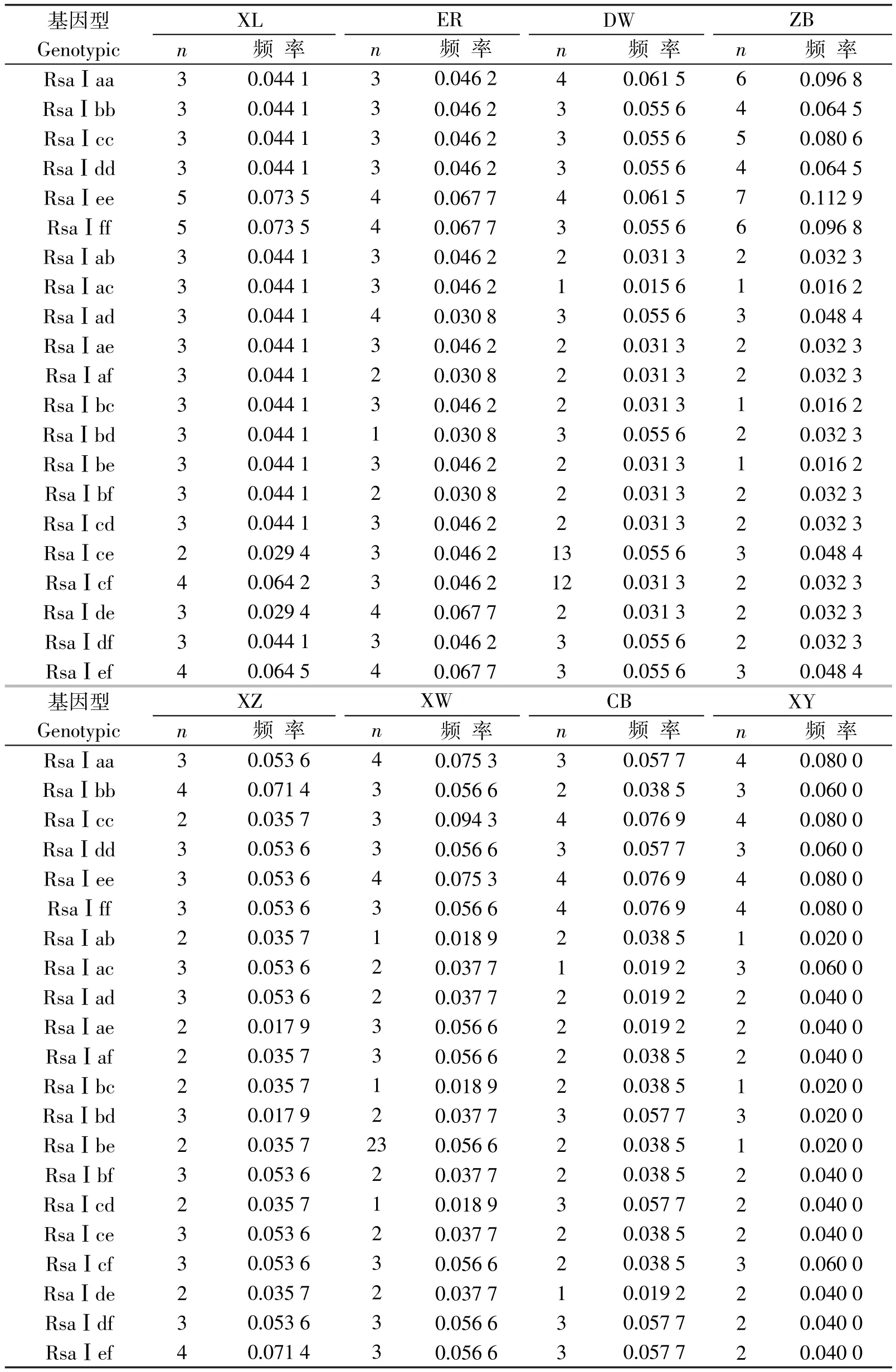
表1 布氏田鼠种群MHCⅡ类基因第二外显子基因型和基因型频率
2.2 MHCⅡ类基因第二外显子序列分析
8个地理种群460只布氏田鼠的MHCⅡ类基因第二外显子的序列均为261 bp,共发现57个变异位点,定义21个单倍型,其中一个单倍型为8个种群共享(见图2).
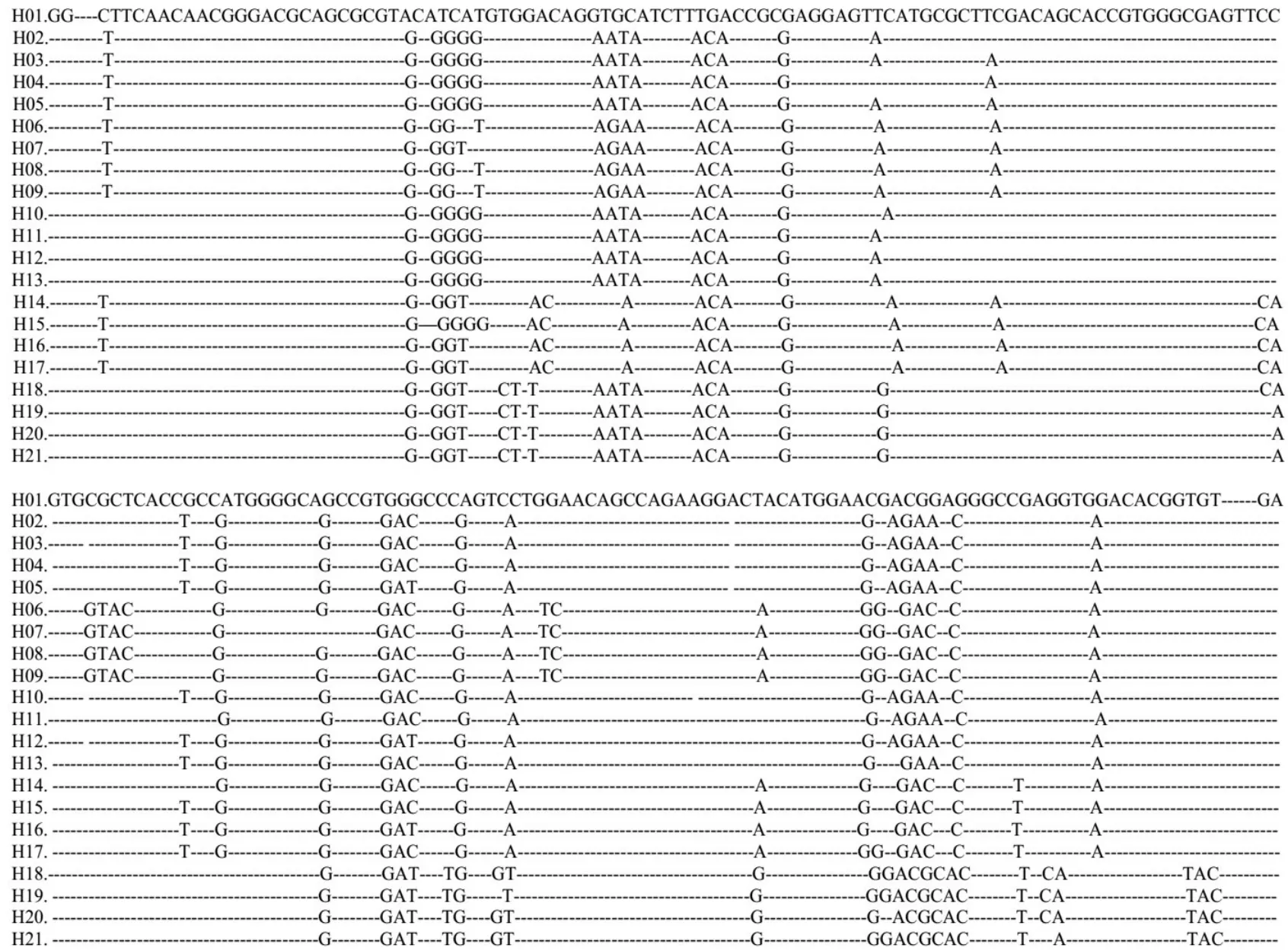
图2 布氏田鼠单倍型变异位点,两端相同的序列被省略
2.3 聚类分析
用UPGMA法对8个布氏田鼠地理种群进行聚类分析,结果显示,8个地理种群分为3个进化分支.把这3个进化单元划分为A、B、C 3个区域(见图3).从图3可以看出,8个地理种群分为3组,分别对应A、B、C 3个区域(见图3),正镶白旗种群(ZB)与其他种群遗传距离最远,为单独一组;XZ、XY、CB被分为一组;ER、XL、XW、DW为一组,其又分为两个亚组,ER、XL为一亚组,XW、DW为一亚组.ER、XL、XW和DW聚为一类,与ZB之间遗传距离较远,但是与XZ、XY和CB有较近的遗传距离.
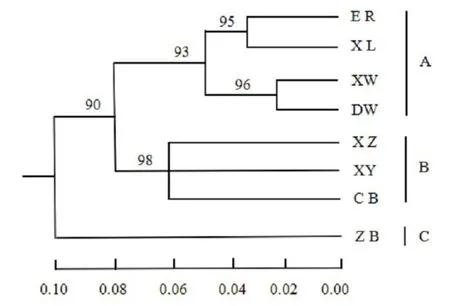
图38 个布氏田鼠种群的UPGMA聚类分析
2.4 布氏田鼠种群遗传多样性及遗传结构分析
种群遗传多样性按系谱发育树A、B、C 3个区域来分析.结果显示,B区域种群单倍型多样度π和核苷酸多样度h最高,A区域种群次之,C区域种群最低(见表2).内蒙古地区布氏田鼠各种群间的遗传多样性差异较大,B区域的3个种群的遗传多样性最高,A区域的4个种群较高,C区域种群的遗传多样性最低.
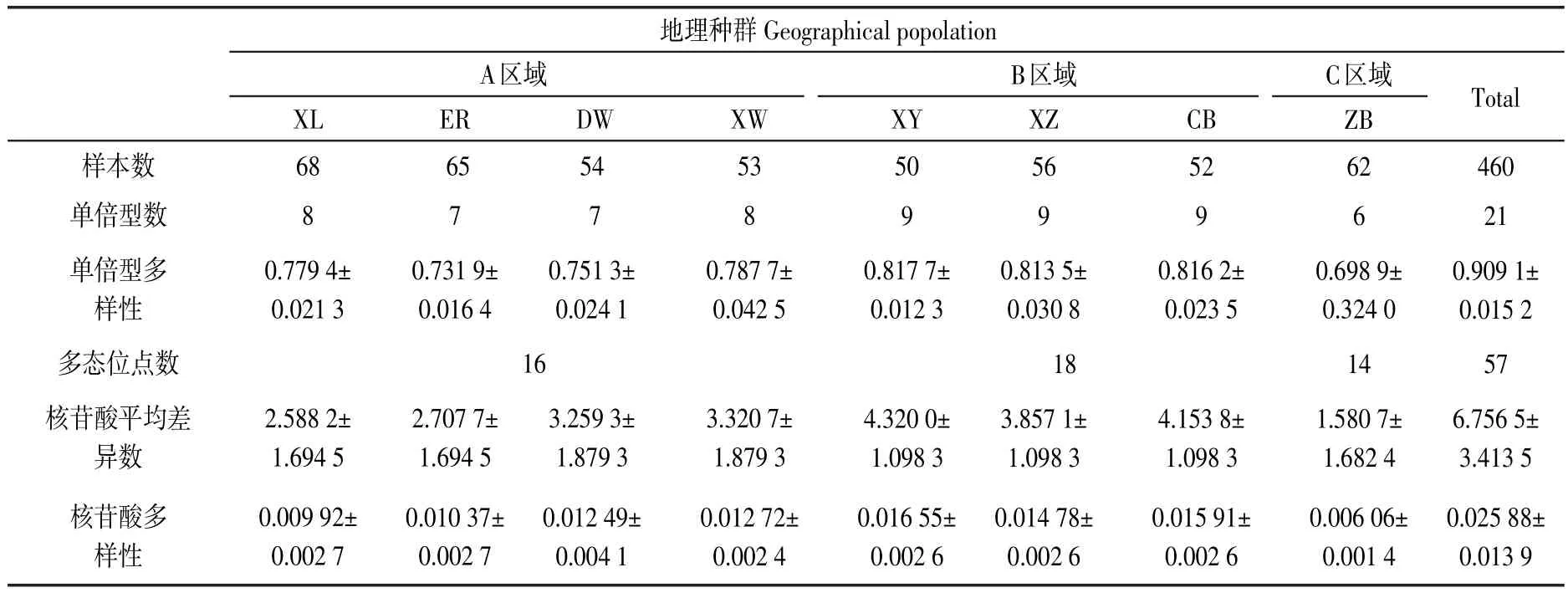
表2 布氏田鼠遗传多样性
AMOVA分析结果表明,种群内个体间变异组分占总变异比率的28.14%,区域内种群间占总变异比率的7.78%,区域类群间占总变异比率的60.48%.说明种区域群间的遗传多样性变化较明显,且核苷酸差异分析表明,C区域种群与其他区域种群之间存在显著的遗传差异.
区域种群间的遗传分化指数FST值(对角线以下)(见表3)分析显示,区域C的正镶白旗种群与A、B区域种群的FST值最大,分别为0.823 2、0.834 7,其FSTP值均小于0.05的检验水平.各区域种群间的基因交流均很小(均小于1),尤其是区域C种群与区域A、B种群的Nm值最低,分别为0.083 5和0.078 8.表明区域C种群具有明显的遗传分化,区域A和区域B种群间出现分化的趋势.遗传分化主要发生在区域类群之间.
同时,分析8个地理种群间的遗传分化指数FST值和种群间的遗传距离(见表4),结果显示,布氏田鼠种群呈现一定程度的分化,分化程度较低,但正镶白旗种群与其他种群间的遗传距离(p-distance)最大,均达到19%以上,分化则较为明显.与上述遗传分化的结果基本一致.

表3 布氏田鼠区域种群的遗传分化系数FST(上三角)和基因流Nm(下三角)
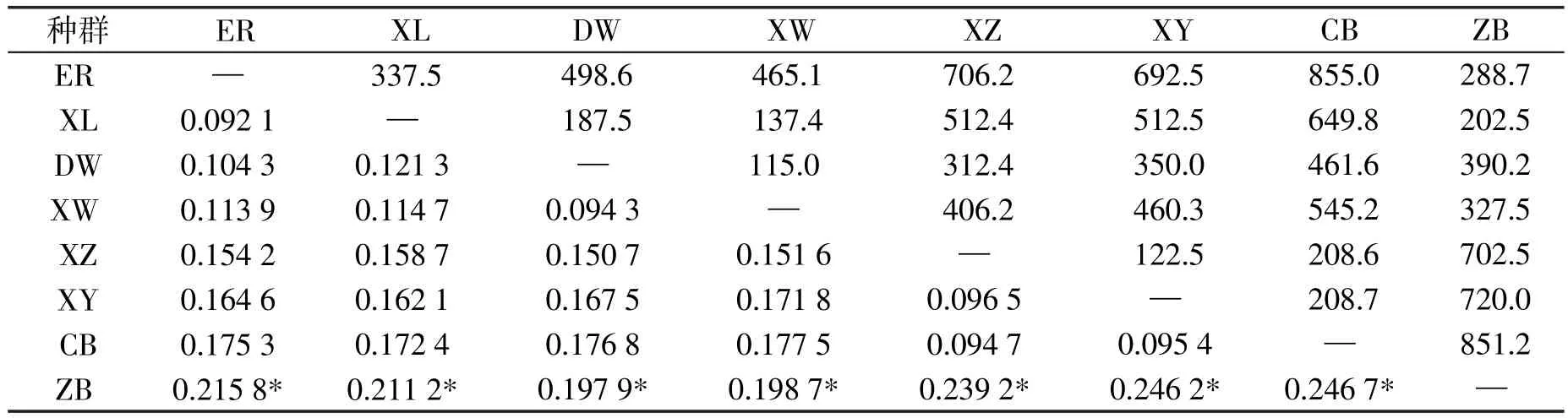
表4 布氏田鼠8个不同地理种群地理距离(上三角)和遗传距离(下三角)
2.5 Mantal tests分析
运用Mantel检验分析,8个小种群的遗传距离(GD)与地理距离(GGD)之间呈现显著的相关性,相关系数R2=0.409 6,且P<0.05的显著水平(如图4).也就是说,布氏田鼠各种群间的地理距离愈大,遗传距离也愈大.
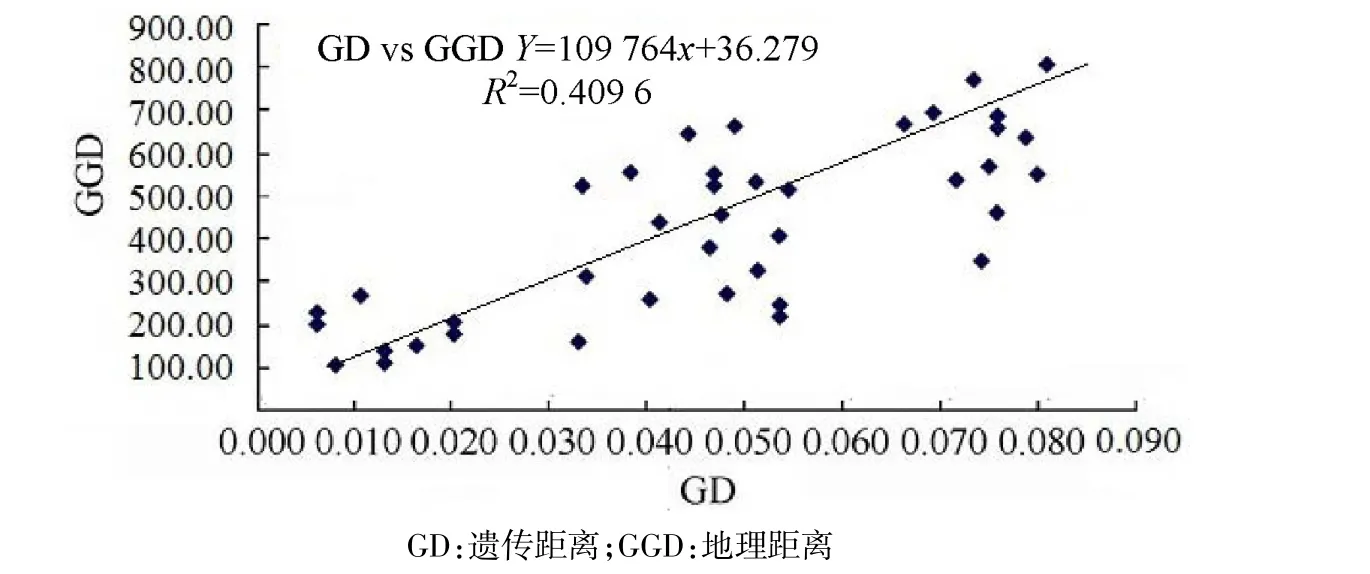
图4 Mantal tests分析结果
3 讨论
3.1 布氏田鼠种群遗传结构
种群的遗传结构受突变、基因流、自然选择和遗传漂变的影响,或者是取决于种群单位间的扩散规模[5-7],研究表明,诸多文献地理隔离和天然地理屏障可引起种群分化[8-13].本研究采集的8个布氏田鼠种群形成3个稳定的分支,分别与采集的地理种群相吻合:同一地理种群内单倍型之间遗传差异小,而不同区域种群间的单倍型之间存在较大区别.根据AMOVA分析,布氏田鼠地理群体间的变异明显大于群体内的变异,该结果说明布氏田鼠存在一定的地理分化.遗传距离、基因流和基因分化系数分析结果说明A和B区域类群间已呈现分化趋势,而C区域的正镶白旗种群与其他种群间的分化较明显.导致种群分化可能有以下因素:首先,内蒙古草原复杂多变的地貌形成的地理隔离在很大程度上影响布氏田鼠的迁徙,从而削弱种群间的基因交流.尤其是浑善达克沙地形成布氏田鼠的迁徙地理屏障,逐渐把C区域的种群与其他区域隔离开来,在长期的进化过程中产生遗传分化,这是引起种群遗传分化的地理因素.其次,布氏田鼠的分布和生存与区域气候、植被、地理等因素有关.内蒙古自治区地形狭长,具有不同地理区划内的气候、地形、植被条件[12].栖息地片段化、草地退化和景观变化是引起布氏田鼠种群遗传分化的生态因素.再次,布氏田鼠是一种营聚集生活的小型哺乳动物,虽然分布区域广,繁殖力强,但种群的扩散能力相对较弱,因此可能较易受到距离隔离的影响而限制种群间基因交流,这是产生遗传分化的物种因素.
3.2 距离隔离对布氏田鼠种群遗传结构的影响
根据Wright距离隔离(isolation by distance)的理论,地理隔离是影响物种遗传分化的重要因素之一,同时其分化的程度也随着地理距离的增大而提高[15].这一观点已有诸多文献证实[16-19].距离隔离对种群遗传分化影响随物种改变而改变[20].在种群间距离2~20 km的范围内,小型哺乳动物可发生明显遗传分化[21-22].布氏田鼠广泛分布在内蒙古中东部,营地下挖掘生活方式,迁移距离小,种群的自由扩散受到限制.本文研究的3大区域类群之间的地理距离都在200 km以上,Mantel检验分析表明,地理距离与布氏田鼠种群遗传分化呈正相关性,表明地理距离是影响布氏田鼠群间遗传分化的因素之一.本文只对8个布氏田鼠种群进行分析,更大范围种群遗传关系和遗传结构有待于进一步的研究.
[1]WANG D,GUO Y W,SHI D Z.Genetic structure of Brandt’s vole(Lasiopodomys brandtii)populations in Inner Mongolia,China,based on microsatellite analysis[J].Conserv Genet,2011,12:659-667.
[2]张曼,施海霞,宋铭晶.微卫星标记在布氏田鼠封闭群遗传结构研究中的应用[J].中国比较医学杂志,2015,25(7):34-38.
[3]WAN Q H,ZHU L,WU H,et al.Major histocompatibility complex class II variation in the giant panda(Ailuropoda melan⁃oleuca)[J].Molecular Ecology,2006,15:2441-2450.
[4]魏磊,王承民,胡奕,等.布氏田鼠和青海田鼠自然种群鼠疫抗性差异[J].四川动物,2013,2(2):167-175.
[5]HUMPHRIES C J,WILLIAMS P H,VANE WRIGHT R I.Measuring biodiversity value for conservation[J].Annual R eview of Ecology and Systematics,1995:93-111.
[6]FRANKHAM R.Genetics and extinction[J].Biol Conserv,2005,126:131-140.
[7]SLATKIN M.Gene flow in natural populations[J].Annu Reu Eeol Syst,1985,16:393-430.
[8]蔡振媛,张同作,慈海鑫,等.高原鼢鼠线粒体谱系地理学和遗传多样性[J].兽类学报,2007,27(2):130-137.
[9]单文娟,刘江,马合木提·哈力克.新疆草兔的种群遗传结构和亚种分化[J].动物学研究,2011,32(2):179-187.
[10]刘翠霞,苏建平,张同作,等.青藏高原的地理屏障在高原鼠兔种群分化中的作用[J].四川动物,2013,32(5):651-657.
[11]RICE W R,HOSTER E E.Laboratory experiments on speciation:What have we learned in 40 years?[J].Evolution,1993,47:637-1653.
[12]FOSTER S A,SCOTT K J,CRESKO W A.Nested biological variation and speciation[J].London:Phil Trans R Soc,1998,353:207-218.
[13]TURELLI M,BARTON N H,COYNE J A.Theory and speciation[J].Trends Ecol Evo,2001,116:330-343.
[14]JI S L,FANG X Y.The plague[M].Beijing:People′s Medical Publishing House,1988:110-117.
[15]WRIGHT S.Isolation by distance[J].Genetics,1943,28:114-138.
[16]GOOSSENS B,CHIKHI L,TABERLET P,et al.Microsatellite analysis of genetic variation among and with in Alpinemar⁃mot populations in the French Alps[J].Molecular Ecology,2001,10(1):41-52.
[17]FLOYD C H,VAN VUREN D H,MAY B.Marmots on great basin moutaintops:using genetics to test a biogeographic paradigm[J].Ecology,2005,86(8):2145-2153.
[18]SELONEN V,PAINTER J N,HANSKI I K.Microsatellite variation in the Siberian flying squirrel in Finland[J].Ann Zool Fennici,2005,42:505-511.
[19]TRIZIO I,CRESTANELLO B,GALBUSERA P,et al.Geographical distance and physical barriers shape the genetic struc⁃ture of Eurasian red squirrels(Sciurus vulgaris)[J].Molecular Ecology,2005,14(2):469-481.
[20]PETIT E,MAYER F.Male dispersal in the noctule bat(Nyctalus noctula):Where are the limits?[J]Proceedings of the Royal Society of London-Series B:Biological Sciences,1999,266(1430):1717-1722.
[21]ANTOLIN M F,HORNE B V,BERGER M D,et al.Effective population size and genetic structure of a Piute ground squirrel(Spermophilusmollis)population[J].Canadian Journal of Zoology,2001,79(1):26-34.
[22]STEWART W A,DALLAS J F,PIERTNEY S B,et al.Metapopulation genetic structure in thewater vole,Arvicola terres⁃tris,in NEScotland[J].Biological Journal of the Linnean Society,1999,68:159-171.
Genetic Analysis of Brandt′s Vole(Lasiopodomys Brandtii)Population Based on MHC ClassⅡexon 2 in Inner Mongolia
ZHAO Suhua1,WEI Lei2,3
(1.Suzhou Vocational and Technical College,234101,Suzhou,Anhui,China;
2.Department of Biological and Chemical Engineering,Bozhou University,236800,Bozhou,Anhui,China; 3.National Research Center for Wildlife Born Diseases,Key Laboratory of Animal Ecology and Conservation Biology,Institute of Zoology,Chinese Academy of Sciences,100101,Beijing,China)
The genetic diversity and genetic structure of 8 geo-populations of Lasiopodomys brandtii were ana⁃lyzed using PCR-RFLP and sequences analysis based on MHC classⅡexon 2.The Results showed that 6 al⁃leles and 13 polymorphic loci were detected.The Chi-square test results argued that genotype frequencies in 6 polymorphic loci were not consistent with the Hardy-Weinberg equilibrium.21 haplotypes were defined based on 57 bases mutations,and one haplotype was shared by all populations.Haplotype diversity(H)and nucleotide diversity(πn)of 7 of L.brandtii populations were higher and another was relatively lower.The UP⁃GMA phylogenetic tree showed that the 8 geo-populations clustered into three groups,corresponding well to their geographical distributions.Mantel test confirmed that genetic differentiation was positively correlated with the geographic distance.Analysis of molecularvariance(AMOVA)similarly recovered the distinct differ⁃entiation between geographical populations:regional genetic variation accounted for 60.48%,intraregional population variation accounted for 7.78%,and individual variation accounted for 28.14%.Analysis of gene flow and FSTsuggested that genetic differentiation already occurred among the 8 populations,but a high level was detected between the Zhengxiangbaiqi and other populations.The unique genetic structure of this spe⁃cies was probably the limited dispersal as well as the stable habitats of the subterranean living,and Hun Shan Da Ke Desert played the most important role in population differentiation of L.brandtii.
Lasiopodomys brandtii;MHC classⅡgene;genetic structure
Q 959.837
A
2095-0691(2017)03-0030-07
2017-05-10
安徽省教育厅自然科学研究基金重点项目(KJ2014A253)
赵素华(1966- ),女,安徽萧县人,副教授,博士生,研究方向:动物防疫与免疫.通信作者:魏 磊(1971- ),男,河南永城人,教授,博士,研究方向:系统进化,种群地理学.
