基于耳石微化学的长江安徽和县江段刀鲚生境履历重建
2017-09-12李孟孟姜涛KHUMBANYIWADavisonDaniel刘洪波杨
李孟孟姜 涛KHUMBANYIWA Davison Daniel刘洪波杨 健,
(1. 南京农业大学无锡渔业学院, 无锡 214081; 2. 中国水产科学研究院淡水渔业研究中心, 中国水产科学研究院长江中下游渔业生态环境评价与资源养护重点实验室, 无锡 214081)
基于耳石微化学的长江安徽和县江段刀鲚生境履历重建
李孟孟1姜 涛2KHUMBANYIWA Davison Daniel1刘洪波2杨 健1,2
(1. 南京农业大学无锡渔业学院, 无锡 214081; 2. 中国水产科学研究院淡水渔业研究中心, 中国水产科学研究院长江中下游渔业生态环境评价与资源养护重点实验室, 无锡 214081)
为了解长江刀鲚(Coilia nasus)的“生境履历”, 利用电子探针微区分析技术研究了2015年5月17日在长江安徽和县江段捕获的刀鲚短颌鲚与长颌鲚类型耳石Sr和Ca微化学特征。短颌鲚的耳石Sr/Ca值变化动态为2类, 一类比值稳定在2.00左右(1.65±0.87—2.03±0.96), 反映了其纯淡水的生境履历; 另一类比值波动显著, 不仅具有对应淡水生境的低值(2.39±1.18—2.63±0.85), 还具有对应半咸水生境的高值(3.22±1.17—3.29±1.14), 显现了其江海生活的生境履历。长颌鲚耳石的Sr/Ca值均波动显著, 同时具有对应淡水生境的低值(1.35±1.07—2.37±1.04)和半咸水、海水生境的高值(3.47±1.09—8.35±1.25), 表现了其均为溯河洄游的生境履历。Sr面分析结果也与上述Sr/Ca值的结果相印证。长江安徽和县江段刀鲚资源组成复杂, 群体可由纯淡水型和江海生活型的个体共同组成。
刀鲚; 生境履历; 电子探针微区分析; 长江; 安徽和县江段
刀鲚(Coilia nasus), 属鲱形目、鳀科、鲚属, 包括溯河洄游型和淡水定居型2种生态型[1,2], 对应的经济价值截然不同。前者经济价值较高, 捕捞强度较大, 其资源量尤其近来年表现为趋于枯竭, 境况已近濒危[3,4]。传统上仅利用上颌骨的长短为依据,认定刀鲚的长颌鲚(C. nasus, 即其上颌骨长度长于头长)类型为洄游型; 而对上颌骨短于头长的短颌鲚(曾命名C. brachygnathus)类型则被认定为只能淡水定居的生态型[5]。然而, 长江流域淡水定居型长颌鲚和江海洄游型短颌鲚的发现[6—8]否定了上述观点, 同时说明仅利用上颌骨长短来判断刀鲚资源的不同生态型的方法的确会产生误判。而基于目前形态学、分子生物学等方法也无法准确判断刀鲚资源的生态型[9,10], 因而也很难正确评价刀鲚资源群体的组成。与此相对照, 耳石作为硬骨鱼类自身携带的硬组织, 其稳定性好, 不易被重吸收, 记录着鱼类一生中所经历的生活史履历和生境过程[11]。其中, 耳石中Sr/Ca值及Sr含量作为鱼类(特别是洄游型鱼类)的生境的“指纹”元素[12—14], 在研究鱼类生境履历的反演上应该具有更大的优势。
长江安徽江段为刀鲚的主要分布区之一, 该江段刀鲚资源量备受关注, 但近年刀鲚在此江段已形不成汛期[15,16]。同时, 该江段是连接长江江苏江段和江西江段的重要水路, 一直被认为是刀鲚的洄游通道[17,18]。长江安徽和县江段作为刀鲚进入安徽江段的最初的水域, 亦为刀鲚从长江口经由江苏江段上溯至安徽江段甚至江西鄱阳湖的必经江段[13,18],其独特地理位置的重要性不言而喻。另外, 在与长江安徽和县江段接壤的南京江段发现了除存在溯河洄游的刀鲚长颌鲚类型外, 还存在淡水定居的长颌鲚类型[6,7]; 但目前对包括和县在内的长江安徽江段刀鲚的生态型及与相邻江段刀鲚资源的关联性尚缺乏了解和确认。因此, 本研究拟基于本实验室前期研究工作[19,20], 利用电子探针微区分析技术对捕获于长江安徽和县江段的刀鲚短颌鲚和长颌鲚类型个体的耳石微化学特征进行分析测定, 以期为较全面地了解该江段刀鲚2种不同生态型的状况,重建该江段刀鲚资源的生境履历和生活史类型的组成动态提供第一手研究资料和理论依据。
1 材料与方法
1.1 材料
刀鲚样品共17尾, 于2015年5月17日采集自长江安徽和县江段(距长江口约440 km)。首先进行样品编号、基本数据测量(表 1), 而后取下左右矢耳石, 用去离子水清洗干净, 晾干备用。
本研究统一使用左矢耳石(以下简称“耳石”)开展微化学研究。耳石样品前处理(包括包埋、碾磨)参考Jiang等[19]的处理方法, 碾磨过后暴露核心的耳石配合抛光液并在装有机织布抛光盘的LaboPol-35磨抛机上抛光至耳石表面没有明显刮痕。接着耳石放于Milli-Q水中超声清洗5min, 晾干, 真空镀膜机(JEE-420, 日本电子株式会社)镀膜(36 A, 25s)。
1.2 耳石微化学分析
参考Yang等[12]的方法, 对耳石进行Sr和Ca的微化学分析, 包括定量线分析和面分布分析。线分析的参数设定: 加速电压为15 kV, 电子束电流为2.0× 10–8A; 束斑直径为5 μm, 每点驻留时间15s; 以间距10 μm连续进行打点测定。标样使用碳酸钙(CaCO3)和钛酸锶(SrTiO3)。所有耳石线分析完后, 进行面分布分析。此时, 加速电压不变, 电子束电流为5.0×10–7A, 束斑直径为5 μm, 像素为7 μm×7 μm, 每点驻留时间为30ms。
1.3 数据分析
Jiang等[19]在研究我国沿海不同水域刀鲚群体时建立了淡水系数(FC), 该值主要反映洄游性鱼类对淡水生境的依存程度。FC=Lf/LT, 其中Lf指从耳石核心开始一直保持Sr/Ca比低值的分析径长, LT指从耳石核心到边缘的整个分析径长。
所有数据主要应用Excel 2008进行汇总和作图, 并参考卢明杰等[21]的STARS (Sequential t-test analysis of regime shifts)方法来客观分析和判定本研究刀鲚耳石Sr/Ca值(按惯例标准化为Sr/Ca×1000,下同)的格局转变(本文设定截断长度为10、Huber权重为1、置信度P为0.1绘制耳石Sr/Ca值)。另外,使用SPSS 20.0分析软件进行非参数检验(Mann-Whitney U-test)。
2 结果
2.1 刀鲚耳石Sr/Ca值的定量线分析
短颌鲚个体共8尾, 其上颌骨/头长均<1, 符合传统上刀鲚短颌鲚类型的定义。全长为(22.2± 3.5) cm, 体重为(35.78±22.29) g, 通过读取耳石上的年轮可知, 1+有2尾, 其余6尾均为2龄。根据耳石定量线分析的Sr/Ca值结果可以将所有样品分为2类(图 1)。第一类耳石样品包括15hxcb01、-cb02、-cb04、-cb06、-cb07和-cb08个体, 耳石Sr/Ca值仅可分为1个阶段。结果显示耳石从核心到边缘的Sr/Ca比均较低, 各测定点的比值几乎不超过3(最小值1.65±0.87, 最大值2.03±0.96)(图 1)。第二类耳石样品包括15hxcb03和-cb05个体, 耳石Sr/Ca值可分为2个阶段。个体15hxcb03在核心至第1年轮形成后(距核心1640 μm)的一段耳石径长上的Sr/Ca值平均在3.0以下(2.39±1.18), 之后至1640 μm处Sr/Ca值升高至耳石边缘结束时比值平稳在某一水平(3.29±1.14)。个体-cb05在耳石核心至第1年轮形成之前(距核心690 μm)的一段耳石径上的Sr/Ca值平均在3.0以下(2.63±0.85), 之后至第一年轮之前(690 μm处)Sr/Ca值升高至第一年轮结束比值降低再到第二年轮比值变化较为平稳在某一水平(3.22± 1.17)。这2个样品的2个阶段测点比值组之间的差异均达显著水平 (P<0.05) (表 2)。
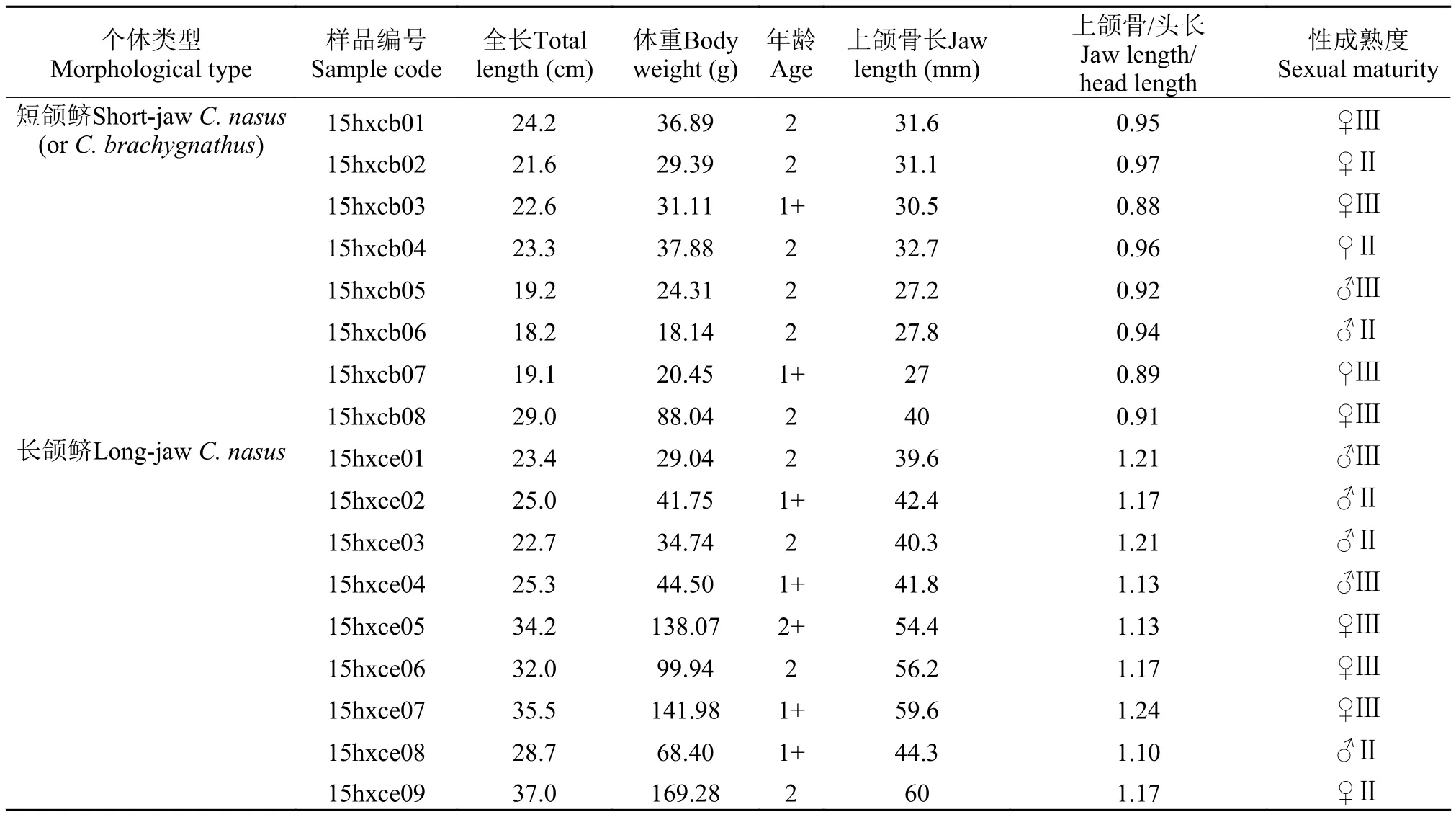
表 1 长江安徽和县江段刀鲚的常规测量数据Tab. 1 Sampling details of Coilia nasus from Hexian section of the Yangtze River in Anhui Province
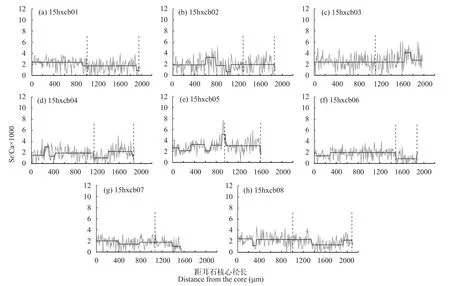
图 1 长江安徽和县江段8尾短颌鲚个体耳石从核心(0 μm)到边缘定量线分析的Sr/Ca值变化
长颌鲚个体共9尾, 其上颌骨/头长均>1, 符合传统上刀鲚长颌鲚类型的定义。全长为(29.3± 5.5) cm, 体重为(85.30±53.42) g, 通过读取耳石上的年轮可知, 1+有2尾, 2龄有6尾, 剩余1尾为2+龄。根据耳石定量线分析的Sr/Ca值结果可将所有样品分为2类(图 2)。第一类包括15hxce01、-ce02、-ce05、-ce06、-ce08和-ce09。它们在核心至第1年轮形成之前(距核心550—1130 μm)的一段耳石径长上的Sr/Ca值均在3.0以下(最小值1.35±1.07, 最大值2.37±1.04), 此低值区为第1阶段。随后, 这些样品Sr/Ca值在第1年轮结束时至第2年轮开始前升高并超过3.0, 其平均值稳定在某一高值(分别为5.20± 1.28、5.04±1.58、6.14±1.53、6.10±1.18、7.99± 1.78和4.09±1.44), 此高值区为第2阶段。在第2年轮阶段Sr/Ca值变化呈现多样, 个体-ce01、-ce02和-ce05和-ce06样品Sr/Ca值先升高后降低。个体-ce08在第1年轮结束后比值先升高后降低, 在最后的一些测定点时比值又再次升高。个体-ce09至第2年轮时比值相对稳定。值得注意的是, 个体-ce08在第2高值阶段为7.99±1.78(>7), 明显高于其他样品的高值阶段(3<Sr/Ca<7)。另一类样品根据其Sr/Ca值的变化可以将其分为低值区和高值区2个阶段, 高值区又可以分为不同的水平, 包括15hxce03、-ce04和-ce07。第1阶段为<3的低值区, 最小为1.99±1.05, 最大为2.28±0.90; 此后比值开始上升, 分别至780—1060、780—1140 和870—1280 μm, 比值分别为3.47±1.09、4.44±1.43和4.90±1.01, 之后比值又再次上升且>7(最小值为7.94±1.14, 最大值为8.35±1.25)至第2年轮时比值降低。各个水平间差异显著(P<0.05)。
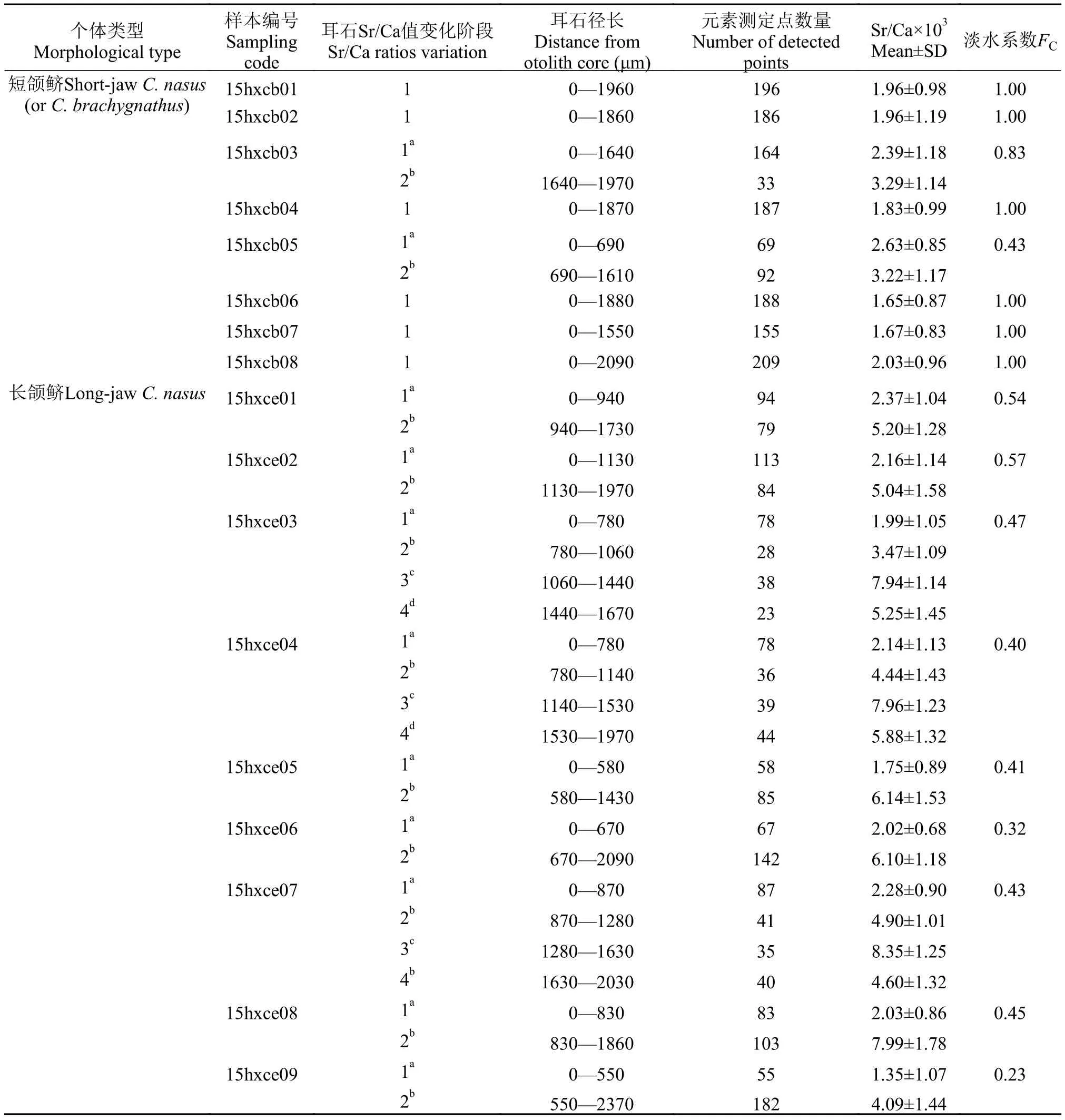
表 2 长江安徽和县江段刀鲚样品耳石中锶和钙的微化学变化Tab. 2 Fluctuation of Sr/Ca ratios along line transects from the core (0 μm) to the edge in otoliths of Coilia nasus from Hexian section of the Yangtze River in Anhui Province
注: 同一样本中相同字母表示不同阶段差异不显著(P>0.05); 不同字母代表差异显著(P<0.05)
Note: Phases in one otolith sample having the same letter indicate not significant differences (P>0.05); whereas different letters indicate significant differences (P<0.05)
2.2 刀鲚耳石Sr含量的面分布分析
耳石Sr元素含量面分布分析图能够更为直观清晰地反映短颌鲚和长颌鲚个体间的差异(图 3)。结果显示, 短颌鲚个体中耳石存在2种类型: 类型一是耳石中核心至边缘具有统一的蓝色的低Sr浓度图, 类型二是耳石除了核心至边缘有蓝色圆环, 蓝色圆环外还有黄绿色的代表高Sr浓度的的圆环。长颌鲚个体中耳石一类是核心的低Sr浓度蓝色核心圆环和外周的高Sr浓度红色圆环, 另一类是核心低Sr浓度蓝色核心圆环, 以及高Sr浓度邻近黄绿色、较高Sr浓度红色圆环和最外周蓝绿色圆环。
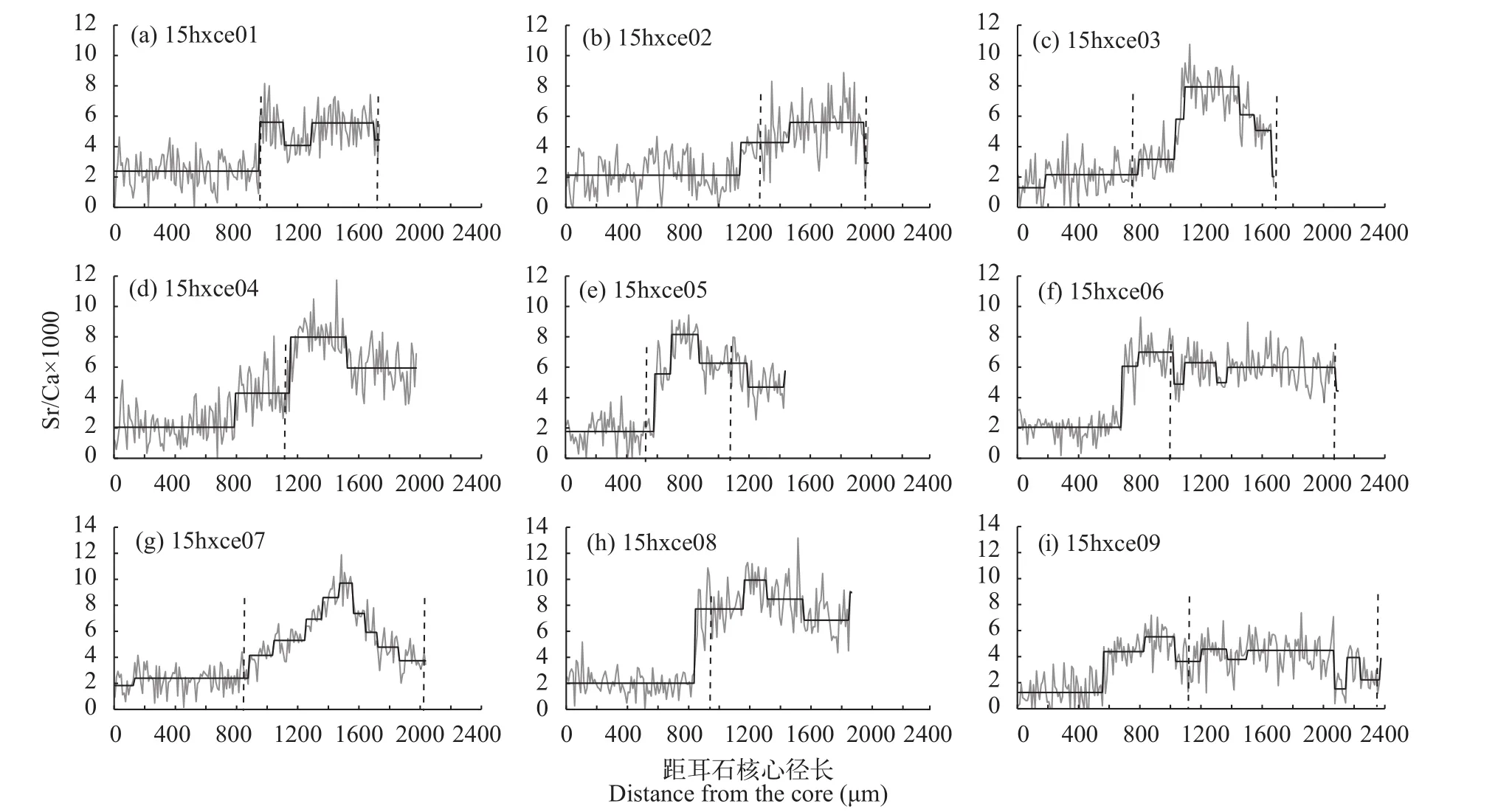
图 2 长江安徽和县江段9尾长颌鲚个体耳石从核心(0 μm)到边缘定量线分析的Sr/Ca值变化
3 讨论
3.1 长江安徽和县江段刀鲚短颌鲚类型的生境履历特征
长江干流中分布有刀鲚的一类短颌鲚群体, 它与长江中分布的另一类长颌鲚群体形态很相似, 过去常将上颌骨长短是否大于头长作为一个重要判断标准[1]。袁传宓等[1]曾将两者归类为不同的物种,近年研究发现两者未达到不同物种2%的遗传距离阈值, 应属同一个物种的两种类型[2]。短颌鲚一直被认为是淡水定居的生态型[1], 但随着溯河洄游短颌鲚个体的被发现证明了利用耳石微化学应该更能客观而真实地判定不同刀鲚生态型的生境履历[8]。在本研究中通过耳石微化学测定并分析发现长江安徽和县江段短颌鲚个体的生活史较为复杂。Yang等[12]曾利用EPMA技术研究刀鲚的耳石生境履历发现, 其耳石对应淡水、咸水生境的Sr/Ca值高低差异显著, 并进一步推定了对应不同盐度生境的比值范围及其相应的Sr元素含量面分布分析图像标准, 即确定了分别对应淡水(<3)、半咸水(3—7)、海水(>7)不同生境“履历”的蓝色、绿黄色、红色等直观的Sr元素图谱。参考该标准, 本研究对长江安徽和县江段的短颌鲚的生境履历进行了重建。结果发现, 类型一(包括15hxcb01、-cb02、-cb04、-cb06、-cb07和-cb08个体)耳石Sr/Ca值从核心至边缘均为<3的低值, 显示出这些个体的全生活史都是在淡水生境中完成, 未曾进入过河口半咸水。Jiang等[20]的研究发现溯河洄游生态型的刀鲚不足1龄时就会离开淡水进入高盐度(河口甚至海区)生境生活。而本研究中短颌鲚个体均在1龄以上(甚至2龄), 说明这6尾短颌鲚应该不存在尚未到开始洄游阶段的可能性。类型二(包括15hxcb03、-cb05个体)和前者仅在淡水中生活截然不同。其耳石Sr/Ca值不仅具有<3的低值, 还有>3的高值, 反映其早期生活史在淡水中完成, 之后会转换生境进入河口半咸水水域生活, 然后再溯河进入长江繁殖,并在和县江段被捕获, 表现出了溯河洄游的倾向。短颌鲚的这种现象已不是个例。前人分别在长江江苏靖江江段和通江江西鄱阳湖湖口等水域也有发现[8,22], 充分说明了长江中可参与江海洄游短颌鲚个体的存在。以上结果不仅显现出单纯利用上颌骨长短会造成刀鲚生境履历的误判, 同时也暗示了长江安徽和县江段的短颌鲚资源群体组成复杂,可由纯淡水型和江海生活型的个体共同组成。因此, 今后对包含本江段在内的长江刀鲚资源组成和资源量的评价上应注意区分两者的不同; 同时, 也需要采集更多的江海生活型短颌鲚个体来进一步确证其溯河洄游的习性。
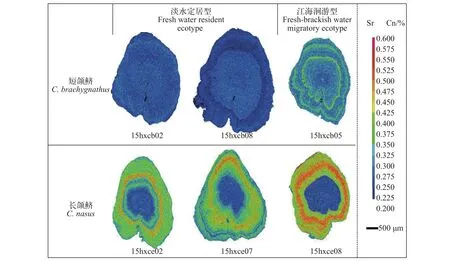
图 3 长江安徽和县江段刀鲚耳石Sr含量的面分析
3.2 长江安徽和县江段刀鲚长颌鲚类型的生境履历特征
洄游型刀鲚亲鱼在2月初开始上溯进行生殖洄游, 孵化的幼鱼会降河入海生长、肥育和越冬[17]。过去常以海洋性寄生虫中国上斧颚虱(Epiclarella chinensis)作为见证其生殖洄游的主要标志, 但在检查安庆江段以上刀鲚时此寄生虫完全消失掉, 该方法仅可有效指示芜湖江段以下的刀鲚群体的洄游[23]。本研究突破上述局限, 通过利用刀鲚耳石微化学的方法更为客观准确地重建其生境履历。结果显示耳石Sr/Ca值不仅具有对应淡水低值, 还存在对应河口半咸水甚至海水的高值, 说明这些长颌鲚个体均参与了江河洄游。通过比较生境履历又不难发现各个体离开淡水后继续肥育时选择的生境并不完全相同, 有的选择在河口生活(15hxce01、-ce02、-ce05、-ce06和-ce09, 其第2阶段Sr/Ca大于3小于7),有的是在河口生活一段时间后进入近海(15hxce03、-ce04、-ce07和-ce08), 这说明个体15hxce01、-ce02、-ce05、-ce06和-ce09在降河后选择了中等的盐度,形成一种生活类型, 而个体15hxce03、-ce04、-ce07、-ce08降河后则需要先在中等盐度生活一段后再过渡至较高盐度生境, 形成第二种生活类型。在Jiang等[19]的研究中也发现刀鲚群体有以上生活模式,这一方面说明一部分刀鲚个体并不降至海里, 而是选择在河口区生活, 可能与长江口沿岸饵料丰富,为刀鲚幼鱼乃至成鱼提供了良好觅食场所有关[24],另一方面也反映了刀鲚个体降河后并不集成大群而是分散生活在沿岸及附近海域[25]。
从淡水系数可知长颌鲚的FC是0.42±0.10, 虽然钱塘江刀鲚可以与长江刀鲚混栖[1], Jiang等[19]发现长江刀鲚的FC值为0.48±0.16, 而毗邻的钱塘江刀鲚的FC值仅是0.22±0.13。由此可见, 本研究中长颌鲚个体的生境模式更符合长江刀鲚群体的生活史特征, 更可能为源自长江产卵场刀鲚资源群体的后代。
目前, 对于刀鲚的长颌鲚和短颌鲚类型的分类地位尚存在争议, 本研究仍将两者作为同种刀鲚的不同类型来研究。但以上结果清楚地表明了, 在长江安徽和县江段捕获的刀鲚不仅生态型复杂, 而且资源群体的组成亦体现出多样性, 即由纯淡水型和江海生活型短颌鲚和溯河洄游型长颌鲚类型个体共同组成。由于纯淡水和溯河的刀鲚资源分属经济价值截然不同的生态型, 很难主观地判别和区分,这也对于今后正确区分长江中现存这两类生态型群体组成, 以便评价其正确的资源量提出了更高要求。同时, 现阶段长江刀鲚(尤其长颌鲚)资源量已显著降低, 前期资料显示, 刀鲚繁殖群体已倾向于由1—2龄个体组成[17]。本研究中和县江段的刀鲚个体正是以1+—2龄为主。这反映出, 近些年长江上所实施刀鲚按证专项定时捕捞的措施虽起到一定的作用, 但资源的低龄化和小型化现象仍日渐明显[24]; 再加上水利工程、拦河建坝等在导致江段的刀鲚洄游通道的破坏[26], 刀鲚资源量减少将愈发严峻[27]。我们认为今后应更加重视研究和保护长江包括和县在内的安徽江段作为长江刀鲚洄游通道不可替代性的作用, 以利保证长江名贵的刀鲚亲鱼可溯河经本江段到达其更上游的产卵场, 而补充群体又可经其到达长江口并入海索饵生长, 从而对整个长江中下游江段和东、黄海区刀鲚资源的养护和可持续利用做出更加实质性的贡献。
[1]Yuan C M, Qin A L, Liu R H, et al. On the classification of the anchovies, Coilia, from the lower Yangtze River and the southeast coast of China [J]. Journal of Nanjing University (Natural Sciences), 1980, (3): 67—82 [袁传宓,秦安黔, 刘仁华, 等. 关于长江中下游及东南沿海各省的鲚属鱼类种下分类的探讨. 南京大学学报(自然科学), 1980, (3): 67—82]
[2]Zhou X D, Yang J Q, Tang W Q, et al. Species validities analyses of Chinese Coilia fishes based on mtDNA COI barcoding [J]. Acta Zootaxonomica Sinica, 2010, 35(4): 819—826 [周晓犊, 杨金权, 唐文乔, 等. 基于线粒体COI基因DNA条形码的中国鲚属物种有效性分析. 动物分类学报, 2010, 35(4): 819—826]
[3]Li Y X, He W P, Liu J S, et al. Annulus validation and age and growth estimation of anadromous Coilia ectenes in the Yangtze estuary [J]. Acta Hydrobiologica Sinica, 2010, 34(4): 787—793 [黎雨轩, 何文平, 刘家寿, 等. 长江口刀鲚耳石年轮确证和年龄与生长研究. 水生生物学报, 2010, 34(4): 787—793]
[4]Mao C Z, Jiao X M, Zhong J S, et al. Research progress on resource status and protection of Coilia nasus in the Yangtze River estuary [J]. Journal of Huaihai Institute of Technology (Natural Science Edition), 2015, 24(3): 78—83 [毛成责, 矫新明, 钟俊生, 等. 长江口刀鲚资源现状及保护研究进展. 淮海工学院学报: 自然科学版, 2015, 24(3): 78—83]
[5]Yuan C M, Lin J B, Qin A L, et al. On the classification history and status quo of genus Coilia in China [J]. Journal of Nanjing University (Natural Sciences), 1976, (2): 1—12 [袁传宓, 林金榜, 秦安舲, 等. 关于我国鲚属鱼类分类的历史和现状—兼谈改造旧鱼类分类学的几点体会. 南京大学学报(自然科学版), 1976, (2): 1—12]
[6]Chen T T, Jiang T, Li M M, et al. Inversion of habitat history for the long-jaw ecotype Coilia nasus collected from Nanjing section of t?he Yangtze River [J]. Journal of Fisheries of China, 2016, 40(6): 1—11 [陈婷婷, 姜涛, 李孟孟,等. 长江南京江段长颌鲚生境“履历”的反演研究. 水产学报, 2016, 40(6): 1—11]
[7]Chen T T, Jiang T, Liu H B, et al. Do all long supermaxilla type estuarine tapertail anchovies Coilia nasus anadromously migrate[J]? Journal of Applied Ichthyology, 2017, 33(2): 270—273
[8]Xu G C, Gu R B, Liu H B, et al. Fluctuation of Sr/Ca in otoliths of Coilia nasus in the Yangtze River and the validation for the anadromous migratory history [J]. Journal of Fisheries of China, 2014, 38(7): 939—944 [徐钢春, 顾若波, 刘洪波, 等. 长江短颌鲚耳石Sr/Ca值变化特征及其江海洄游履历. 水产学报, 2014, 38(7): 939—944]
[9]Cheng W X, Tang W Q. Some phenotypic varieties between different ecotypes of Coilia nasus in Yangtze River [J]. Chinese Journal of Zoology, 2011, 46(5): 33—40 [程万秀, 唐文乔. 长江刀鲚不同生态型间的某些形态差异. 动物学杂志, 2011, 46(5): 33—40]
[10]Liu D, Li Y Y, Tang W Q, et al. Population structure of Coilia nasus in the Yangtze River revealed by insertion of short interspersed elements [J]. Biochemical Systematics & Ecology, 2014, (54): 103—112
[11]Campana S E, Thorrold S R. Otoliths, increments, and elements: keys to a comprehensive understanding of fish populations[J]? Canadian Journal of Fisheries and Aquatic Sciences, 2001, 58(1): 30—38
[12]Yang J, Arai T, Liu H B, et al. Reconstructing habitat use of Coilia mystus and Coilia ectenes of the Yangtze River estuary, and of Coilia ectenes of Taihu Lake, based on otolith strontium and calcium [J]. Journal of Fish Biology, 2006, 69(4): 1120—1135
[13]Jiang T, Yang J, Lu M J, et al. Discovery of a spawning area for anadromous Coilia nasus Temminck et Schlegel, 1846 in Poyang Lake, China [J]. Journal of Applied Ichthyology, 2017, 33(2): 189—192
[14]Jiang T, Liu H B, Huang H H, et al. Otolith microchemistry of Coilia grayii from the Parl River Estuary, China [J]. Acta Hydrobiologica Sinica, 2015, 39(4): 816—821 [姜涛, 刘洪波, 黄洪辉, 等. 珠江口七丝鲚耳石微化学初报. 水生生物学报, 2015, 39(4): 816—821]
[15]Wan Q, Lai N Y, Li F, et al. Analysis on the change of reproductive population composition of Coilia ectenes in Wuwei section of Anhui in Yangtze River [J]. Journal of Hydroecology, 2009, 30(4): 60—65 [万全, 赖年悦, 李飞,等. 安徽无为长江段刀鲚生殖洄游群体年龄结构的变化分析. 水生态学杂志, 2009, 30(4): 60—65]
[16]Zhu X F, Gao C H, Yao C Y, et al. Saury resource protection in Yangtze River [J]. Chinese Fisheries Economics, 2009, 27(3): 41—44 [朱孝锋, 高成洪, 姚彩媛, 等. 长江刀鱼资源的危机和保护对策. 中国渔业经济, 2009, 27(3): 41—44]
[17]Yuan C M. Spawning migration of Coilia nasus [J]. Bulletin of Biology, 1987, (12): 1—4 [袁传宓. 刀鲚的生殖洄游. 生物学通报, 1987, (12): 1—4]
[18]Li Y X, Xie S G, Li Z J, et al. Gonad development of an anadromous fish Coilia ectenes (Engraulidae) in lower reach of Yangtze River, China [J]. Fisheries Science, 2007, 73(6): 1224—1230
[19]Jiang T, Liu H B, Shen X Q, et al. Life history variations among different population of Coilia nasus along theChinese coast inferred from otolith microchemistry [J]. Journal of the Faculty of Agriculture Kyushu University, 2014, 59(2): 383—389
[20]Jiang T, Yang J, Liu H, et al. Life history of Coilia nasus from the Yellow Sea inferred from otolith Sr: Ca ratios [J]. Environmental Biology of Fishes, 2012, 95(95): 503—508
[21]Lu M J, Liu H B, Jiang T, et al. Preliminary investigations on otolith microchemistry of Odontamblyopus rubicundus in the Daliao River Estuary, China [J]. Marine Fisheries, 2015, 37(4): 310—317 [卢明杰, 刘洪波, 姜涛,等. 大辽河口红狼牙鰕虎鱼耳石微化学的初步研究. 海洋渔业, 2015, 37(4): 310—317]
[22]Lu M J. Studies on otolith morphometry and microchemistry of Coilia nasus collected from the Poyang Lake [D]. Thesis for Master of Science. Shanghai Ocean University, Shanghai. 2015 [卢明杰. 鄱阳湖水域刀鲚耳石的形态学和微化学研究. 硕士学位论文, 上海海洋大学,上海. 2015]
[23]Xu H N, Sun C B, Tong Y R, et al. The biological indicator of the spawning migration of anchovy (Coilia ectenes Jordan et Seale) in Yangtze River [J]. Journal of Nanjing University (Natural Sciences), 1978, (3): 85—91 [徐㞧南, 孙超白, 童远瑞, 等. 长江流域刀鲚鱼生殖洄游的“生物指标”. 南京大学学报(自然科学), 1978, (3): 85—91]
[24]Guo H Y, Zhou T S, Tang W Q, et al. Temporal pattern of Coilia nasus biomass along the Jingjiang section of the Yangtze River over the last decade [J]. Resources and Environment in the Yangtze Basin, 2015, 24(4): 565—571 [郭弘艺, 周天舒, 唐文乔, 等. 长江近口段沿岸刀鲚生物量的时间格局. 长江流域资源与环境, 2015, 24(4): 565—571]
[25]Yuan C M, Qin A L. Ecological habits and distribution of Coilia along the Chinese coast and its changes of output [J]. Marine Sciences, 1984, 8(5): 35—37 [袁传宓, 秦安舲. 我国近海鲚鱼生态习性及其产量变动状况. 海洋科学, 1984, 8(5): 35—37]
[26]Shi W G, Zhang M Y, Liu K, et al. Stress of hydraulic engineering on fisheries in the lower reaches of the Yangtze River and compensation [J]. Journal of Lake Sciences, 2009, 21(1): 10—20 [施炜纲, 张敏莹, 刘凯, 等. 水工工程对长江下游渔业的胁迫与补偿. 湖泊科学, 2009, 21(1): 10—20]
[27]Tian S Q, Tian Z Q, Gao C X, et al. Analyzing of annual changes for the stock status of Coilia nasus in fishing season in Yangtze River estuary [J]. Journal of Shanghai Ocean University, 2014, 23(2): 245—250 [田思泉, 田芝清, 高春霞, 等. 长江口刀鲚汛期特征及其资源状况的年际变化分析. 上海海洋大学学报, 2014, 23(2): 245—250]
RECONSTRUCTING HABITAT HISTORY OF COILIA NASUS FROM THE HEXIAN SECTION OF THE YANGTZE RIVER IN ANHUI PROVINCE BY OTOLITH MICROCHEMISTRY
LI Meng-Meng1, JIANG Tao2, KHUMBANYIWA Davison Daniel1, LIU Hong-Bo2and YANG Jian1,2
(1. Wuxi Fisheries College, Nanjing Agricultural University, Wuxi 214081, China; 2. Key Laboratory of Fishery Eco-environment Assessment and Resource Conservation in Middle and Lower Reaches of the Yangtze River, Freshwater Fisheries Research Center, Chinese Academy of Fishery Science, Wuxi 214081, China)
Coilia nasus is a valuable fish in the middle and lower reaches of the Yangtze River. In order to reconstruct its habitat history, an electronic probe microanalyzer (EPMA) was applied to analyze the microchemistry patterns of element Sr and Ca in otoliths of 17 individuals of two phenotypes belonging to short-jaw C. nasus (once named as C. brachygnathus) and long-jaw C. nasus. Au sample were collected in Hexian section of the Yangtze River in Anhui Province at May 2015. Microchemical results showed that C. brachygnathus individuals could be divided into two types: (1) Sr/Ca ratio was consistently at low phase (<3.0) throughout the otolith, indicating that the individuals only experienced freshwater habitat during their whole life history; (2) Sr/Ca ratio fluctuated between low (2.39±1.18—2.63±0.85) and high (3.22±1.17—3.29±1.14) phases in the otoliths, suggesting the individuals experienced not only freshwater, but also brackish water habitats at different stages in their life history. The otolith Sr/Ca ratio of long-jaw C. nasus fluctuated significantly between low (<3, reflecting freshwater habitat) and high (>3, reflecting estuarine brackish water or sea water habitat) value phases. The 16- color-Sr concentration profiles are in the line with the srlca ratios. All of the above results revealed that the resource composition of Coilia nasus from Hexian section of the Yangtze River in Anhui Province was much more complex by consisting of freshwater resident and freshbrackish water migratory short-jaw C. nasus, as well as anadromous long-jaw C. nasus individuals.
Coilia nasus; Habitat history; EPMA; The Yangtze River; Hexian section of the Yangtze River in Anhui Province
SG31.5
A
1000-3207(2017)05-1054-08
10.7541/2017.132
2016-11-10;
2017-04-11
国家自然科学基金项目(31372533); 中央级公益性科研院所基本科研业务费专项(2016PT01); 江苏省自然科学基金项目(BK20131089)资助 [Supported by the National Natural Science Foundation of China (31372533); the Central Research Institutes of Basic Research and Public Service Special Operations, China (2016PT01); the Natural Science Foundation of Jiangsu Province (BK20131089)]
李孟孟(1991—), 女, 河南商丘人; 硕士研究生; 主要研究方向为渔业生态环境监测与保护。E-mail: 18015357281@163.com
杨健, E-mail: jiany@ffrc.cn
猜你喜欢
杂志排行

水生生物学报的其它文章
- 剩余污泥资源化利用新工艺研究进展
- 草鱼野生与选育群体线粒体DNA控制区D-loop遗传变异分析
- 在患CyHV-2病的异育银鲫肠道黏膜中胆固醇、胆汁酸代谢通路基因的差异表达
- 黄颡鱼20β-羟基类固醇脱氢酶Ⅰ和Ⅱ基因特征分析和表达模式研究
- 池蝶蚌β-连环蛋白基因cDNA的克隆及表达特征分析
- MOLECULAR CLONING, CHARACTERIZATION, AND EXPRESSION ANALYSIS OF TWO ISOFORMS OF ANTI-LIPOPOLYSACCHARIDE FACTOR FROM THE ORIENTAL RIVER PRAWN, MACROBRACHIUM NIPPONENSE