在患CyHV-2病的异育银鲫肠道黏膜中胆固醇、胆汁酸代谢通路基因的差异表达
2017-09-12叶元土蔡春芳林秀秀吴代武张宝彤萧培珍
叶元土 吴 萍 蔡春芳 林秀秀 吴代武 何 杰 张宝彤 萧培珍
(1. 苏州大学基础医学与生物科学学院, 江苏省水产动物营养重点试验室, 苏州 215123; 2. 北京营养源研究所水产动物系统营养研究开放试验室, 北京 100000)
在患CyHV-2病的异育银鲫肠道黏膜中胆固醇、胆汁酸代谢通路基因的差异表达
叶元土1吴 萍1蔡春芳1林秀秀1吴代武1何 杰1张宝彤2萧培珍2
(1. 苏州大学基础医学与生物科学学院, 江苏省水产动物营养重点试验室, 苏州 215123; 2. 北京营养源研究所水产动物系统营养研究开放试验室, 北京 100000)
为了探讨CyHV-2疾病条件下异育银鲫胆汁酸肠肝循环代谢途径主要基因表达活性的变化, 以患CyHV-2病的、正常的异育银鲫肠道黏膜为材料, 提取总RNA, 采用RNA-Seq测序、对单一基因进行注释, 并进行KEGG富集分析、单一基因差异表达分析。结果显示, 肠道黏膜组织有7770个基因显著性差异表达, 其中, 差异表达上调基因数为3335个、差异表达下调的基因数为4435个, 表明CyHV-2病的发生对肠道黏膜组织基因表达产生了重大的影响。病鱼胆固醇、胆汁酸生物合成代谢途径的酶、蛋白质的基因显著性差异表达, 显示肠道黏膜胆固醇、胆汁酸合成代谢受到显著性影响。参与胆固醇、胆汁酸合成代谢调节作用, 以及胆固醇、胆汁酸分泌、吸收、转运等生理过程的蛋白质、酶的基因也是差异表达下调, 胆汁酸肠肝循环有出现代谢障碍的趋势。结果表明, CyHV-2病的发生对肠道黏膜组织基因表达产生了严重影响, 对胆固醇、胆汁酸的合成代谢途径、胆汁酸肠肝循环途径的基因表达产生了严重影响, 将导致病鱼体内胆固醇、胆汁酸量的不足。患CyHV-2病病鱼血清胆汁酸含量下降了99%、胆固醇含量下降了10%, 证实了上述基因差异表达的趋势。
氧化鱼油; 黏膜; 胆固醇; 胆汁酸; 异育银鲫
油脂是水产动物饲料的主要营养物质和能量物质, 而油脂易氧化, 其氧化的中间产物、终产物对水产动物具有氧化损伤作用[1,2]。在草鱼饲料中添加氧化鱼油[3]或者对草鱼灌胃氧化鱼油[4], 导致草鱼肝胰脏、肠道黏膜胆固醇生物合成通路基因差异表达, 血清、肠道内胆汁酸含量显著下降; 胆汁酸的肠肝循环受到严重的障碍, 而胆汁酸的肠肝循环对于维护肝胰脏和肠道组织结构完整性和功能完整性具有重要的生理作用[4]。异育银鲫(Carassius auratus gibelio)受到鲤疱疹Ⅱ型病毒(CyHV-2)感染后出现全身性出血症状, 并导致主要器官组织如肝胰脏、脾脏、肾脏、肠道黏膜、血液等组织严重损伤; 同时, 病鱼血清的胆固醇和胆汁酸含量显著下降[5—7]。肠道黏膜是动物体内代谢活跃的器官组织, 也是胆固醇、胆汁酸合成的重要器官组织之一, 本文分别以正常的、患CyHV-2病的异育银鲫肠道黏膜为材料, 提取总RNA后采用高通量测序技术(RNA-Seq)进行转录组分析, 以正常异育银鲫的结果为对照, 在转录组水平上, 对参与胆固醇、胆汁酸生物合成通路的蛋白质、酶的基因进行功能注释和差异表达活性分析, 探讨在患CyHV-2病的情况下, 异育银鲫肠道黏膜组织中胆固醇、胆汁酸合成代谢的酶、蛋白质基因表达活性的变化, 以及参与胆汁酸肠肝循环的酶、蛋白质基因表达活性的变化, 从一个侧面、从转录组水平较为宏观的视觉探讨异育银鲫患CyHV-2病的病理代谢变化, 为该病的病理分析和防治提供参考。
1 材料与方法
1.1 试验鱼
江苏盐城大丰市池塘养殖的异育银鲫。正常鱼无CyHV-2病发病特征, 病鱼则是具有CyHV-2病发病特征、经过PCR定量检测带有CyHV-2病毒的异育银鲫[5—7]。
参照林秀秀等[5—7]对异育银鲫CyHV-2病鉴定方法, 用于血清胆固醇、胆汁酸含量分析的试验鱼包括正常(H)组(CyHV-2的PCR检测结果阴性)、携带CyHV-2(CyHV-2的PCR检测结果阳性)但未出现病症(IH)组、典型的CyHV-2病症(I)组(CyHV-2的PCR检测结果阳性)异育银鲫, 体重为350—510 g,每组各9尾、共27尾。
1.2 血清采集与总RNA提取
按照常规方法采集正常组(H)、携带CyHV-2但无病症组(IH)、患CyHV-2病并具有典型病症组(I)的异育银鲫血清, 用雅培C800全自动生化分析仪测定血清胆固醇和胆汁酸含量。
正常组、患CyHV-2病组异育银鲫肠道黏膜各9个样本(每尾鱼为1个样本), 分别用试剂盒(EASYspin Plus)每尾鱼独立用于提取总RNA, 电泳检测RNA质量。正常组、病鱼组分别选取电泳条带亮度高、清晰、无拖尾的高质量RNA进行等量混合, 正常组、病鱼组各1个样本(每个样本至少有5尾试验鱼的总RNA样本组成), RNA质量约4 μg。同样方法再混合一个RNA样本, 其余的RNA舍弃。正常组、病鱼组分别得到2个(n=2)平行样本用于RNA-seq分析。
1.3 转录组分析
RNA-Seq 样品总RNA用DNaseⅠ消化DNA后, 用Oligo d (T)磁珠纯化总RNA中的mRNA,向得到的mRNA中加入适量打断试剂, 在高温条件下使其片段化, 再以片段后的RNA为模板, 合成cDNA, 经过磁珠纯化、末端修复、3'末端加碱基A、加测序接头后, 进行PCR扩增, 从而完成整个文库制备工作。构建好的文库用Agilent 2100 Bioanalyzer和ABI Step One Plus Real-Time PCR System质检和测序。读段长度125 bp。
无参转录组的转录本拼接与基因功能注释
综上所述,在经济社会发展的过程中,建筑施工企业想要保障自身的持续稳定发展,必须要对自身成本管控工作进行完善,结合新时期发展趋势转变思想观念,健全成本管控体制,加强专业人才培养,完善风险防控,如此才能为成本管控工作的实施创造良好的条件,为企业持续稳定发展奠定坚实基础。
经过质控(QC)、过滤后得到读段(clean reads)用Trinity (版本: v2012-10-05, min_kmer_ cov为2)软件进行无参考基因组的转录本拼接。
对拼接好的转录本利用不同的数据库进行unigene基因注释。各数据库及功能注释所用到的方法: ①与SwissProt、KOG、KEGG GENES序列数据库的比对采用NCBI blast 2.2.27+, 经过Swissprot注释得到了38071个、KOG注释得到32345个单一基因。②PFAM蛋白结构域预测采用HMMER 3.0 package-hmmscan, 经过Pfam注释得到10227个单一基因。③GO功能注释为基于Swissprot和Pfam两部分的蛋白注释结果, 软件为Blast2GO v2.5, 经过GO注释得到36838个单一基因。
基因差异表达分析与KEGG通路分析 采用RSEM软件, 以Trinity拼接得到的转录组作为参考序列, 将每个样品的clean reads对参考序列做比对进行基因表达定量。差异表达分析用edgeR软件包进行分析。对于所有基因, 当其表达量在正常组、病鱼组两个不同样品间具有差异且统计分析中调整P值(假阳性率)<0.05时, 认为其在两个不同样品中具有显著差异表达。
以KEGG Pathway为单位, 应用超几何检验, 找出与整个基因组背景相比、在差异表达基因中显著性富集的Pathway。以correctedP-value≤0.05为阈值, 满足此条件的定义为在差异表达基因中显著富集的KEGG条目。
2 结果
2.1 血清胆固醇、总胆汁酸含量
由表 1可知, 异育银鲫携带CyHV-2后, 在IH组、I组, 其血清中胆汁酸和胆固醇含量显著下降。与正常鱼体的结果比较, 病鱼血清总胆汁酸含量下降了99%、血清胆固醇含量下降了10%。
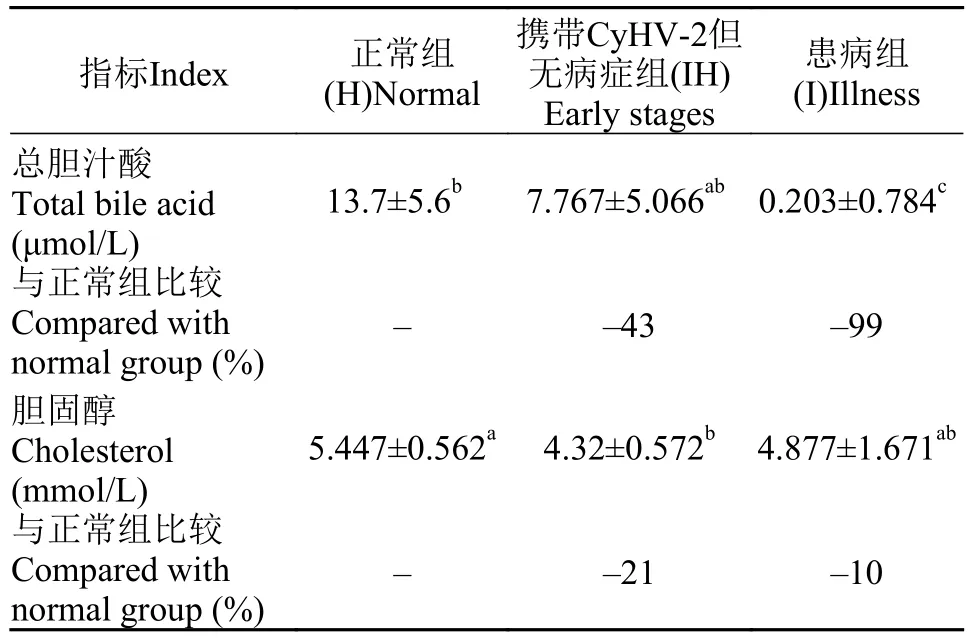
表 1 异育银鲫血清胆固醇和总胆汁酸含量与比较Tab. 1 Gibel carp serum total cholesterol and bile acid content
2.2 基于转录组分析的基因差异表达
基于通过注释的单一基因结果, 在正常、患CyHV-2病异育鲫鱼样本间进行了基因差异表达分析, 总共有7770个显著差异表达的基因, 其中, 显著差异表达上调基因数为3335个, 占差异表达基因总数的43%; 显著表达下调的基因数为4435, 占57%。由表 2可知, 在KEGG类固醇合成通路的21个基因中, 有15个基因差异表达, 占通路基因总数的71%;在KEGG初级胆汁酸合成通路的20个基因中, 有14个基因显著差异表达, 占通路基因总数的70%;上述2个KEGG通路差异表达均达到显著性水平(P<0.05)。上述结果显示, 异育银鲫在患CyHV-2病后, 肠道黏膜组织的基因发生了显著性的差异表达,其中胆固醇、胆汁酸的生物合成通路基因也发生了显著性的差异表达。

表 2 基因显著差异表达的KEGG路径富集Tab. 2 Differentially expressed genes of KEGG enrichment
2.3 胆固醇合成通路基因的差异表达
基于患CyHV-2病与正常的异育银鲫肠道黏膜转录组分析结果, 统计了胆固醇生物合成途径有18个具有差异表达的基因(表 3)。将注释得到的单一基因序列, 利用NCBI/ Blast进行相对于不同物种相同基因的同源性分析。表 3中的18个基因与斑马鱼、安水金线鲃、犀角金线鲅、滇池金线鲃中相同基因的同源性较高, 同源性达到78%—100%。
log2(I/H)是将患病组单一基因的表达量与对应的正常组基因表达量比对, 进行以2为底的对数值计算的结果, 该值越大表明基因表达量的差异越大,正值为差异表达上调、负值则为差异表达下调;当≥2、或≤–2时可以视为显著差异表达。
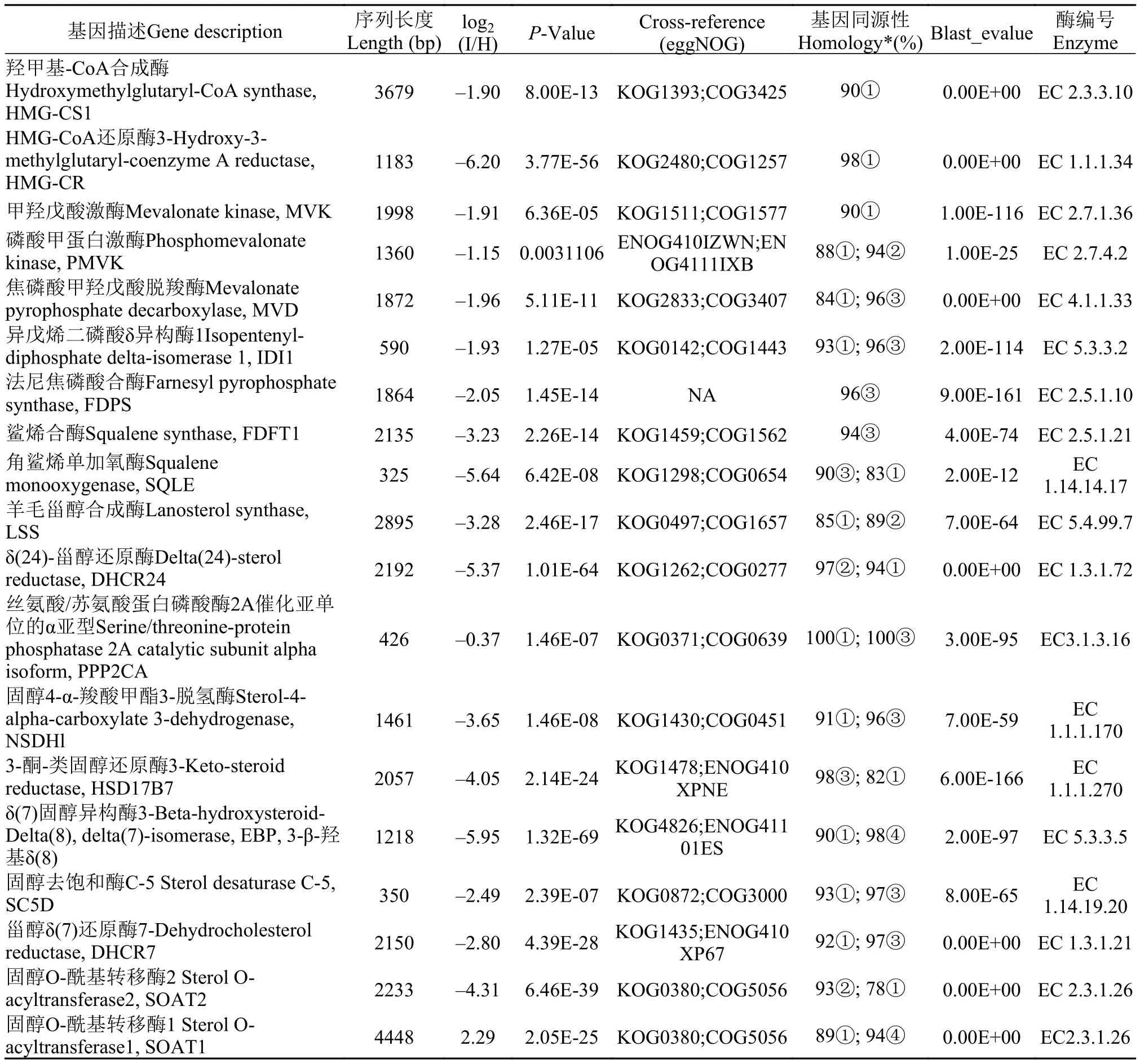
表 3 CyHV-2病异育银鲫肠道黏膜组织胆固醇生物合成通路基因差异表达信息Tab. 3 Differentially expressed genes of cholesterol biosynthetic pathway in intestinal mucosa
在表 3中, 仅有固醇O-酰基转移酶1(SOAT1)的log2(I/H)为正值, 且大于2, 表明为显著差异表达上调; 其他17个基因的log2(I/H)值全部为负值, 表明为差异表达下调, 其中, 胆固醇生物合成通路关键酶HMG-CoA还原酶等12个基因的log2(I/H)值小于“–2”, 为差异表达显著下调。
上述结果表明, 异育银鲫在患CyHV-2病后, 肠道黏膜组织的胆固醇生物合成通路的绝大多数基因显著差异表达下调, 可能影响到肠道黏膜组织的胆固醇生物合成量的不足。
2.4 胆汁酸合成通路基因的差异表达
表 4中9个基因序列相对于斑马鱼、安水金线鲃、犀角金线鲅相同基因的同源性为83%—98%。由表 4可知, 参与胆汁酸合成的9个基因的log2(I/H)值均为负值, 均为差异表达下调; 其中, 胆汁酸合成通路关键基因胆固醇7-α单加氧酶(CYP7A1)、固醇26-羟化酶(CYP27A1)等7个基因的log2(I/H)值小于“–2”, 达到差异表达显著下调的水平。
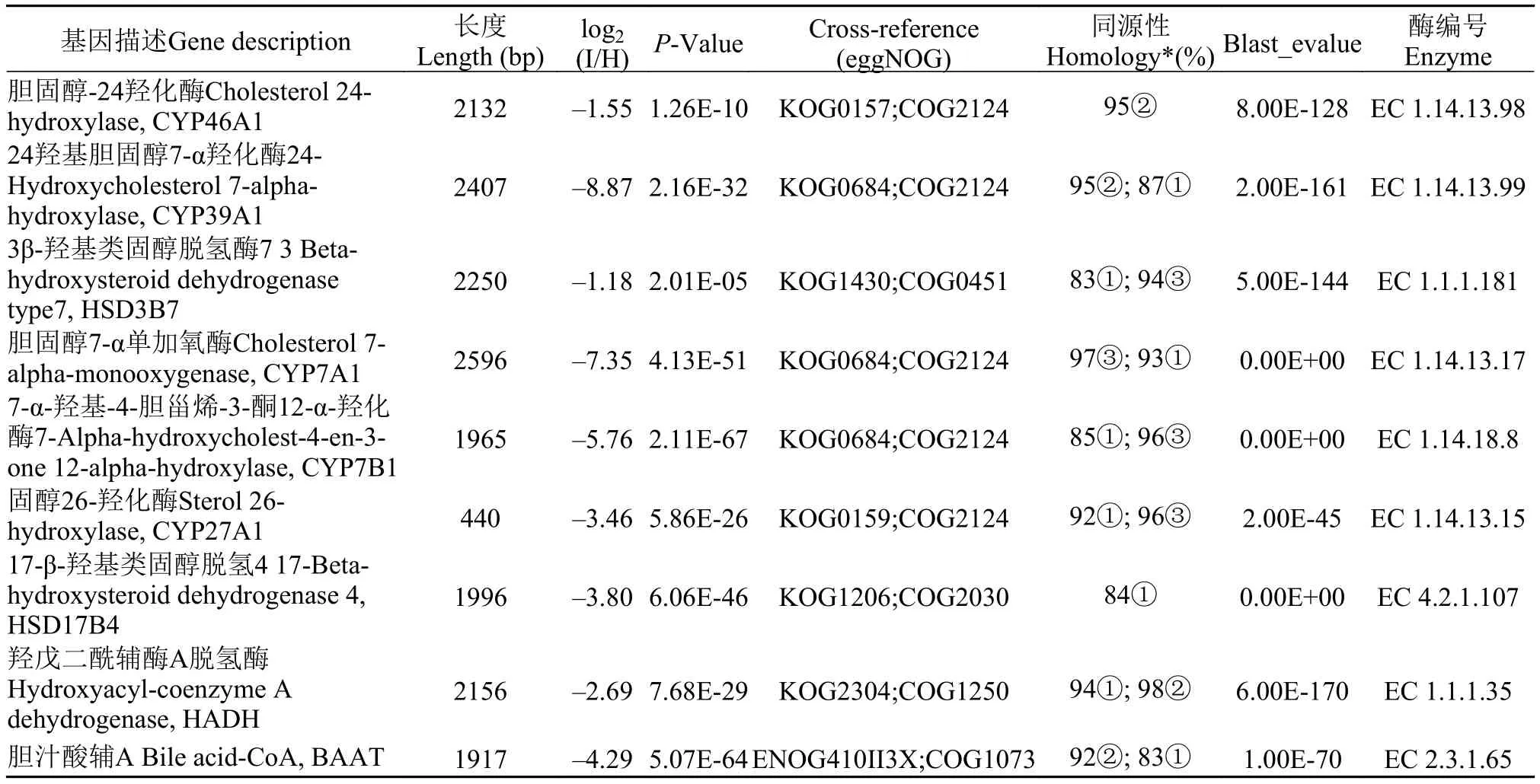
表 4 CyHV-2病的异育银鲫肠道黏膜组织胆汁酸生物合成通路基因差异表达信息Tab. 4 Differentially expressed genes of bile acid biosynthesis pathway in intestinal mucosa
上述结果显示, 在异育银鲫患CyHV-2病后, 肠道黏膜组织中胆汁酸合成通路的基因差异表达均为下调, 可能影响到胆汁酸生物合成量的不足, 表明CyHV-2病的发生将严重影响肠道黏膜组织中胆汁酸生物合成代谢途径基因的表达活性。
2.5 参与胆固醇、胆汁酸生物合成代谢调节的蛋白质、酶基因的差异表达
表 5 中11个基因序列相对于斑马鱼、安水金线鲃、犀角金线鲅、鲤鱼、草鱼相同基因的同源性为68%—99%。由表 5可知, 涉及胆固醇转运的胆固醇酯转移蛋白(CETP)的log2(I/H)值为–1.64, 调节胆固醇生物合成通路的限速酶HMG-CoA还原酶表达的固醇调节元件结合蛋白(SREBF1、SREBF2)的log2(I/H)值为–2.55、–1.51, 低密度脂蛋白受体(LDLR)的log2(I/H)值为–3.85, 氧固醇受体LXR-α的log2(I/H)值为–1.40, 均为负值, 显示在患CyHV-2病后, 这些基因表达活性下调, 将会引起肠道黏膜组织胆固醇的生物合成量可能下降。
在涉及胆汁酸分泌、吸收和转运的蛋白质中,仅有胆汁酸受体(FXR)的log2(I/H)值为2.63, 为差异表达上调, 其他的如胆盐输出泵(B S E P)的log2(I/H)值为–2.02, 回肠钠/胆汁酸协同转运蛋白(ASBT)的log2(I/H)值为–8.51, 肠胆汁酸结合蛋白(IBABP)的log2(I/H)值为–5.63, 肝胆汁酸结合蛋白(LFABP)的log2(I/H)值为–5.63, 胆汁酸辅酶A(BAAT)的log2(I/H)值为–1.06基因表达活性均为表达下调。
上述结果表明, 在肠道胆固醇、胆汁酸生物合成通路基因差异表达下调的同时, 涉及胆固醇生物合成调节、胆固醇转运的蛋白质基因差异表达下调, 涉及胆汁酸分泌、吸收和转运的蛋白质基因差异表达下调, 其结果可能导致胆固醇、胆汁酸生物合成量不足。而血清胆固醇、胆汁酸含量在患Cy-HV-2病后显著下降, 也证实了上述结果。
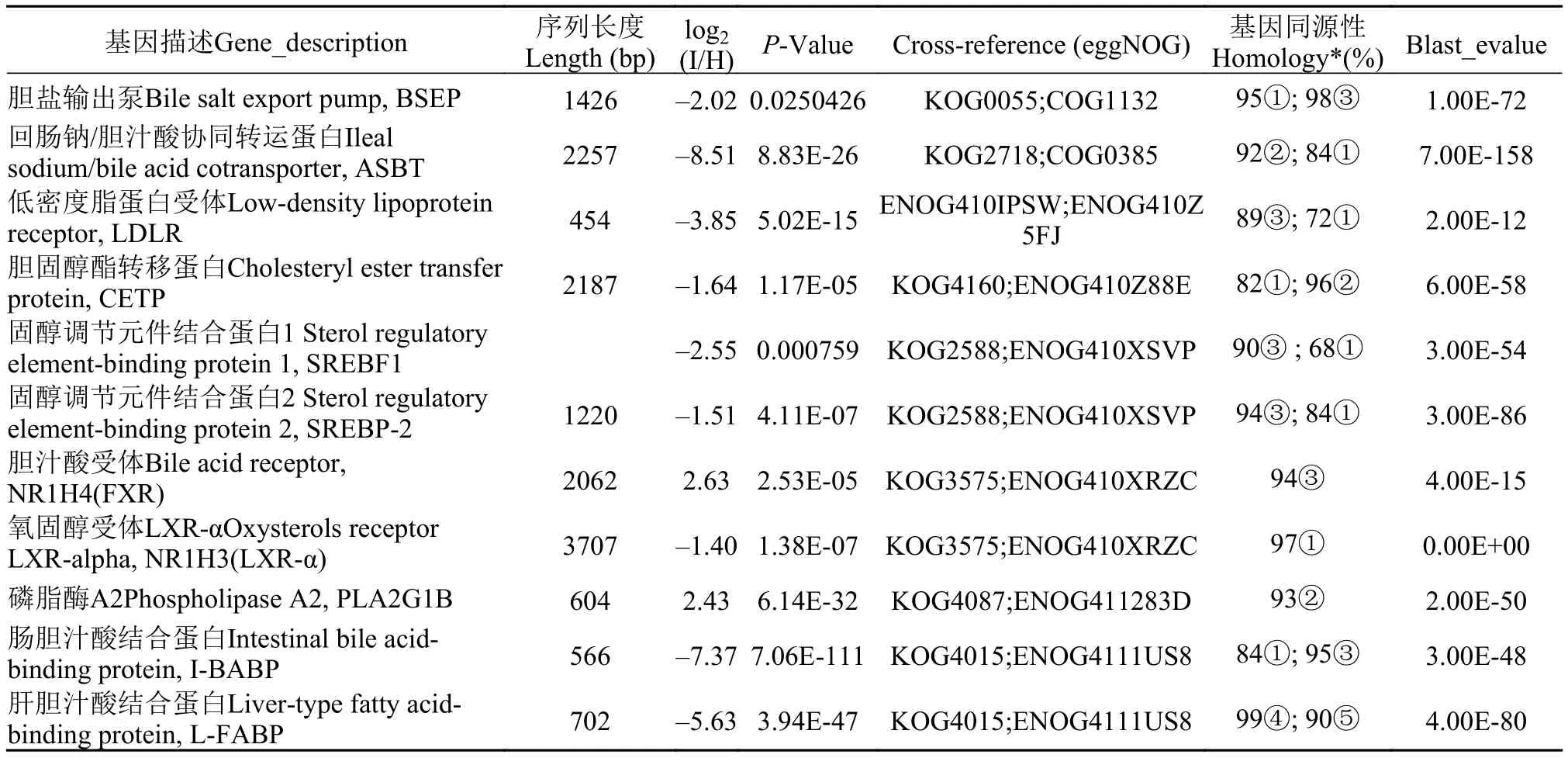
表 5 参与胆固醇、胆汁酸代谢的蛋白质、酶基因的差异表达信息Tab. 5 Differentially expressed genes of cholesterol, bile acid metabolism
3 讨论
3.1 患CyHV-2病的异育银鲫肠道黏膜组织胆固醇生物代谢通路基因的差异表达下调
内环境稳定(内稳态)是动物正常生理代谢的基础。一些生理活性物质如胆固醇、胆汁酸在动物体内保持动态稳定具有重要的生理意义[4,8,9]。胆固醇的来源包括2个方面, 一是由体内组织细胞释放的胆固醇、从食物中摄取并进入血液的胆固醇;另一个来源则是, 鱼类与其他动物一样, 能够以乙酰CoA为原料合成胆固醇。胆固醇在体内不能被彻底氧化分解为CO2和H2O, 而经氧化和还原反应转变为其它含环戊烷多氢菲母核的化合物, 主要代谢去路包括作为甾醇类激素生物合成原料、作为胆汁酸合成的原料、转化为维生素D等[8]。
固醇O-酰基转移酶(SOAT)又称酰基转移酶(Cholesterol acyltransferase, ACAT), 是细胞内已知的、唯一催化游离胆固醇与长链脂肪酸连接形成胆固醇酯的酶, 有2种同工酶SOAT1、SOAT2或称ACAT1、ACAT2。SOAT1在各种组织和细胞中都有表达, 主要作用是催化胆固醇的酯化反应、维持细胞内胆固醇的代谢平衡。同时, 有研究表明[10],在血液中的单核细胞分化成巨噬细胞后SOAT1大量表达, 导致细胞内胆固醇酯的过量堆积而形成泡沫细胞。林秀秀等在异育银鲫CyHV-2病研究中发现, 病鱼血液细胞发生显著变化, 产生过度的炎症反应[5,6,7]。在本文中, 病鱼肠黏膜中SOAT1的 log2(I/H)值为2.29, 是表 3中唯一差异表达显著上调的基因, 这是否与血液单核细胞分化为巨噬细胞有关值得进一步的研究。SOAT2则只在肝脏和小肠细胞(主要在小肠上皮细胞绒毛的顶端)中表达, 主要参与胆固醇的吸收以及脂蛋白的装配。食物来源的胆固醇是以游离的形式被肠道黏膜细胞被动吸收, 胆固醇进入肠黏膜细胞后受SOAT2催化, 胆固醇与脂酰辅酶A结合生成胆固醇脂肪酸酯, 并与载脂蛋白组装为乳糜微粒进入淋巴系统或血液系统。病鱼肠黏膜中SOAT2的log2(I/H)值为–4.31, 显示肠道黏膜细胞内利用胆固醇形成胆固醇酯、组装为乳糜微粒的SOAT2作用下降。也表明, 从食物、其他细胞转运来的胆固醇量可能不足。在表 5中, 影响胆固醇吸收的低密度脂蛋白受体(LDLR)的log2(I/H)值为–3.85, 影响胆固醇转运的胆固醇酯转移蛋白(CETP)为–1.64, 显示均为差异表达下调, 这些结果也表明, 异育银鲫患CyHV-2病后,从其他器官组织吸收、转运胆固醇能力下降。
细胞内胆固醇的另一个主要来源是以乙酰辅酶A为原料合成胆固醇, HMG-CoA还原酶(HMGCR)是胆固醇生物合成代谢途径的限速酶, 该酶位于内质网上的膜整合糖蛋白, 是体内胆固醇生物合成的关键酶, 其log2(I/H)值为–6.20, 差异表达显著下调。从表 3也可以发现, 经过基因注释、差异表达分析得知, 胆固醇生物合成途径的多个酶蛋白基因均是差异表达下调的, log2(I/H)值为(–6.20)—(–0.37)。在表 5中, 参与胆固醇生物合成代谢调节作用的固醇调节元件结合蛋白(SREBF1、SREBP-2)的log2(I/H)值为–2.55、–1.51, 也是差异表达下调的。
已经有研究表明, 在饲料氧化油脂刺激下[3,4],以及草鱼肠道疾病病理条件下[11], 都会导致肠道黏膜组织的胆固醇、胆汁酸合成代谢发生显著的变化。综合上述分析可以发现, 异育银鲫在患CyHV-2病后,一方面显示胆固醇代谢受到严重影响, 食物吸收和转运胆固醇的能力下降, 以乙酰辅酶A为原料合成胆固醇的能力下降, 参与胆固醇合成代谢起调节作用的蛋白、酶的基因表达活性下降, 可能导致鱼体器官组织、血液中胆固醇含量的不足。病鱼血清中胆固醇含量较正常鱼下降10%也证实了这种趋势。另一方面, 在对异育银鲫CyHV-2病进行预防或治疗的生产实践中,有必要通过饲料途径补充一定量的胆固醇, 这值得进一步的深入研究和验证。
3.2 患CyHV-2病的异育银鲫肠道黏膜组织胆汁酸代谢通路基因差异表达下调
以胆固醇为原料合成胆汁酸有2种途径[8,9]: 经典途径和替代途径。经典途径是由胆固醇7α羟化酶(CYP7A1)催化, CYP7A1是此反应的限速酶; 替代途径是由甾醇27α羟化酶(CYP27A1)和甾醇12α羟化酶(CYP7B1)催化的途径。胆固醇7-α单加氧酶(CYP7A1)是以胆固醇为原料合成胆汁酸代谢经典途径的限速酶, 胆汁酸的合成速度与CYP7A1的活性呈正相关关系, 决定了胆固醇向胆汁酸转化的速度[3,4]。由表 4可知, CYP7A1的log2(I/H)值为–7.35, 显示其差异表达显著下调。代谢途径中后续的胆汁酸合成代谢酶的log2(I/H)值均为负值, 显示胆汁酸合成路径的催化酶基因差异表达均为下调。CYP7A1的活性和表达量受固醇受体(LXR-α)的调节, 而LXR-α的log2(I/H)值为–1.40, 也是差异表达下调。由表 4可知, 胆汁酸合成替代途径有甾醇27α羟化酶(CYP27A1) log2(I/H)值为–3.46、甾醇12α羟化酶(CYP7B1) log2(I/H)值为–5.76, 也是差异表达显著下调。结果表明, 无论是经典途径或是替代途径, 胆汁酸合成途径的限速酶、参与催化代谢途径的酶, 其基因差异表达均为下调。
关于胆汁酸的分泌、重吸收途径, 胆汁酸在肝脏合成后由胆盐输出泵(BSEP)泵入肠道, 游离胆汁酸在肠道由黏膜细胞通过扩散作用被动重吸收, 而结合胆汁酸通过小肠刷状缘的钠盐依赖的胆汁酸转运体(ASBT)被黏膜细胞主动重吸收, 黏膜细胞内的胆汁酸与回肠胆汁酸结合蛋白(IBABP)结合, 由黏膜基侧膜吸收入静脉、回到肝胰脏, 完成胆汁酸的肠肝循环[8,9]。在表 5中, BSEP的log2(I/H)值为–2.02, 显示涉及胆汁酸分泌的蛋白质基因差异表达显著下调。ASBT、肠胆汁酸结合蛋白(I-BABP)、肝胆汁酸结合蛋白(L-FABP)的log2(I/H)值分别为–8.51、–7.37、–5.63, 显示涉及胆汁酸重吸收、转运蛋白质的基因表达活性也是显著性下调, 胆汁酸有在肝胰脏淤积的发展趋势。这些结果显示, 患CyHV-2病后, 异育银鲫胆汁酸的分泌、胆汁酸的肠肝循环途径中主要蛋白质基因的表达活性也发生了显著性变化, 均为显著性下调, 可能导致胆汁酸的肠肝循环障碍。病鱼血清胆汁酸含量下降99%也证实了这种趋势。
胆汁酸受体(FXR)是胆汁酸的感受器, 属于核受体, 受FXR调控的靶基因主要有CYP7A1、BSEP、IBABP等。当胆汁酸浓度过大时, 使CYP7A 1转录受到抑制, 胆汁酸合成速度下降。由表 4、表 5可知, FXR、CYP7A1、BSEP、IBABP的log2(I/H)值分别为2.63、–7.35、–2.02、–7.37, 结果显示, 即使FXR差异表达显著上调, 但其下游涉及胆汁酸合成、重吸收、转运的蛋白质基因还是差异表达显著下调。
上述结果表明, 异育银鲫在患CyHV-2病后, 肠道黏膜组织胆汁酸生物合成代谢能力下降, 可能导致胆汁酸含量的不足。胆汁酸的分泌、重吸收、转运的蛋白质基因差异表达下调, 可能导致胆汁酸的肠肝循环障碍。病鱼血清总胆汁酸含量下降99%也证实了这种发展趋势。对于异育银鲫Cy-HV-2病的预防、治疗, 以及病后鱼体生理健康的恢复, 通过饲料途径补充适量胆汁酸可能具有可行性, 值得进一步的研究。
[1]Ye Y T, Cai C F, Wu P. Injury of Oxidized Dietary Oil on Growth and Health of Grass Carp [M]. Beijing: China Agricultural Science and Technology Press. 2015, 137—625 [叶元土, 蔡春芳, 吴萍. 氧化油脂对草鱼生长和健康的损伤作用. 北京: 中国农业科技出版社. 2015, 137—625]
[2]Chen K Q, Ye Y T, Cai C F, et al. Effects of dietary oxidizes fish oil on growth and muscle fatty acid composition of grass carp (Ctenopharyngodon idellus) [J]. Chinese Journal of Animal Nutrition, 2015, 27(6): 1698—1708 [陈科全, 叶元土, 蔡春芳, 等. 饲料中氧化鱼油对草鱼生长及肌肉脂肪酸组成的影响. 动物营养学报, 2015, 27(6): 1698—1708]
[3]Huang Y W, Ye Y T, Cai C F, et al. The effect of cholesterol and bile acid metabolism synthesis pathway related enzymes in the liver and intestine under oxidized fish oil in Ctenopharyngodon idellus [J]. Genomics and Applied Biology, 2015, 34(8): 1636—1646 [黄雨薇, 叶元土, 蔡春芳, 等. 氧化鱼油对草鱼(Ctenopharyngodon idellus)肝胰脏、肠道胆固醇、胆汁酸合成代谢的影响. 基因组学与应用生物学, 2015, 34(8): 1636—1646]
[4]Ye Y T, Cai C F, Xu F, et al. Feeding grass carp (Ctenopharyngodon idellus) with oxidized fish oil up-regulates the gene expression in the cholesterol and bile acid synthesis pathway in intestinal mucosa [J]. Acta Hydrobiologica Sinica, 2015, 39(1): 90—100 [叶元土, 蔡春芳, 许凡, 等. 灌喂氧化鱼油使草鱼肠道黏膜胆固醇胆汁酸合成基因通路表达上调. 水生生物学报, 2015, 39(1): 90—100]
[5]Lin X X, Ye Y T, Wu P, et al. The injury effect of infection by cyprinid herpesvirus 2 (CyHV-2) on tissues and organs of gibel carp (Carassius auratus gibelio) [J]. Genomics and Applied Biology, 2016, 35(3): 587—594 [林秀秀, 叶元土, 吴萍, 等. 鲤疱疹Ⅱ型病毒(CyHV-2)感染对异育银鲫(Carassius auratus gibelio)组织器官的损伤作用. 基因组学与应用生物学, 2016, 35(3): 587—594]
[6]Lin X X, Ye Y T, Wu P, et al. Ultra-pathological observation of cyprinid herpesvirus 2 (CyHV-2) of a gibel carp (Carassius auratus gibelio) with hematopoietic necrosis [J]. Chinese Journal of Fisheries, 2016, 29(1): 17—23 [林秀秀, 叶元土, 吴萍, 等. 异育银鲫造血器官坏死症病鱼体内鲤疱疹病毒Ⅱ型的电镜观察与超微病理学特征.水产学杂志, 2016, 29(1): 17—23]
[7]Lin X X, Ye Y T, Wu P, et al. Variation in blood biochemical indices and histopathology in gibel carp (Carassius auratus gibelio) Infected with cyprinid herpesvirus Ⅱ [J]. Chinese Journal of Fisheries, 2016, 29(2): 16—23 [林秀秀, 叶元土, 吴萍, 等. 感染鲤疱疹病毒Ⅱ型 (CyHV-2) 的异育银鲫血液生理生化指标及相应组织学变化. 水产学杂志, 2016, 29(2): 16—23]
[8]Zhang J C, Nie Q H. Metabolism of bile acid and related diseases progression [J]. China Journal Gastroenterology and Hepatology, 2008, 17(11): 953—956 [张久聪, 聂青和. 胆汁酸代谢及相关进展. 胃肠病学和肝病学杂志, 2008, 17(11): 953—956]
[9]Zhang L, Niu S M. The progress of bile acids and metabolic syndrome [J]. Medical Recapitulate, 2016, 22(5): 964—967 [张柳, 牛尚梅. 胆汁酸与代谢综合征的研究进展. 医学综述, 2016, 22(5): 964—967]
[10]Yao X M, Song B L, Wang C H, et al. Human Acyl-coenzyme A: cholester of Aacyltr ansfer ase (ACAT) [J]. Journal of Shanghai Jiaotong University (Agricultural Science), 2006, 24(1): 108—115 [姚晓敏, 宋保亮, 王灿华, 等. 人酰基辅酶A: 胆固醇酰基转移酶(ACAT). 上海交通大学学报 (农业科学版), 2006, 24(1): 108—115]
[11]Huang Y W, Ye Y T, Cai C F, et al. The effect of the gene expression in metabolism of cholesterol synthesis pathway after intestine injuried on Ctenopharyngodon idellus [J]. Journal of Nanjing Agricultural University, 2015, 38(3): 497—503 [黄雨薇, 叶元土, 蔡春芳, 等. 肠道损伤对草鱼胆固醇代谢通路基因表达的影响. 南京农业大学学报, 2015, 38(3): 497—503]
GENE DIFFERENCE EXPRESSION OF CHOLESTEROL AND BILE ACID METABOLISM PATHWAY IN INTESTINAL MUCOSA WITH THE CYHV-2 DISEASE CARASSIUS AURATUS GIBELIO
YE Yuan-Tu1, WU Ping1, CAI Chun-Fang1, LIN Xiu-Xiu1, WU Dai-Wu1, HE Jie1, ZHANG Bao-Tong2and XIAO Pei-Zhen2
(1. Key Laboratory of Aquatic Animal Nutrition, School of Basic Medicine and Biological Science, Soochow University, Suzhou 215123, China; 2. Open Lab for Aquatic Animal Nutrition, Beijing Research Institute for Nutritional, Beijing 100000, China)
Gibel carp (Carassius auratus gibelio) infected with Cyprinid herpesvirus 2 (CyHV-2), damaged many organs including intestinal mucosa. Total RNA of intestinal mucosa from CyHV-2 diseased and normal gibel carp were extracted for RNA-Seq. Transcriptome assembly from RNA-Seq data without a reference genome was conducted by Trinity (version: v2012-10-05, min_kmer_cov 2) software. Single gene is annotated by using different databases. Software and the database of the functional gene annotation are SwissProt (NCBI blast 2.2.27+, Blast2GO v2.5), KOG (NCBI blast 2.2.27+), PFAM (HMMER 3.0 package), KEGG (KOBAS, NCBI blast 2.2.27+). Gene expression was analyzed by edgeR package. 3335 up-regulated genes and 4435 down-regulated genes were observed, indicating a significant injury in the intestinal mucosa by CyHV-2 infection. The metabolic disorder for enterohepatic circulation of bile acids was supported by the reduced gene expression that regulate cholesterol and bile acid synthesis metabolism and other physiological processes, secretion, absorption and transportation. As a consequence, CyHV-2 infection in Crucian carp decreased the serum bile acid content by 99% and cholesterol content by 10%.
Oxidized fish oil; Mucosa; Cholesterol; Bile acid; Gibel carp
S917
A
1000-3207(2017)05-0956-07
10.7541/2017.119
2016-09-07;
2016-12-24
江苏省水产三新工程项目(D2015-12)资助 [Supported by the Jiangsu Province, Three New Aquatic Projects (D2015-12) Funding]
叶元土, 教授, 硕士生导师, E-mail: yeyt@suda.edu.cn, Tel./Fax: +86-13912629760
