人体运动控制理论及计算模型的研究进展
2017-09-03王爱文罗冬梅
王爱文 ,罗冬梅
人体运动控制理论及计算模型的研究进展
王爱文 ,罗冬梅
人体运动控制是一个复杂的、综合了多门学科的领域,先后纳入了物理学、解剖学、生理学、心理学、工程学、神经学等学科的内容。人体运动控制的研究从最初的对人体外在的机械运动的描述逐渐深入到肌肉骨骼,神经控制的层次。人体运动控制领域要解决的基本问题是人体运动的冗余控制、富余控制及运动变异,它们代表了人体运动的多样性,围绕这几个基本问题,学者们提出了各种基于信息控制理论和基于物理过程的各种人体运动控制的假说(动作编码、层级控制、平衡点假说、对照设置假说、动态行为理论等)及多种用于人体运动评估研究的计算框架(黎曼流形、非控制流形、容错-噪声-共变模型、适应性模型理论等)。这些理论假说有的偏重于控制信息的处理过程,有的偏重于人体运动的物理过程,其中每个理论假说都有其意义及局限性,提供研究者不同的思考角度,有的理论,如平衡点假说,则是沉寂一段时间后又被重新发现其实践意义,并受到越来越多的关注。对人体运动的研究,一开始人们关注于单一关节、单一肌肉的状态,然而人体的运动是一个整体,一个关节出现损伤可能主要的原因并不是在这一个关节而是受到其余关节活动的影响,一块肌肉的损伤也可能是由于其余肌肉的影响,甚至人体的损伤也受到人体认知状况的影响,这就需要从整体的角度去重新审视人体运动。人体运动是复杂的,对人体运动的描述不能仅仅是对单一关节、单一肌肉的描述,而是需要更为复杂的计算框架,黎曼流形、非控制流形、容错-噪声-共变模型、适应性模型理论则对人体运动的描述做出了有益的探索。人体运动控制理论对康复及运动训练有着重要的影响,根据运动控制理论学者们提出和更新了传统的康复及训练方法,从运动控制角度设计的训练方法有效地降低了损伤的发生率,延长了运动员的运动寿命。对人体运动控制的进一步研究可以帮助对损伤机制、运动表现、装备器械对人体的影响机制等的深入理解,促进相关领域的发展。
运动控制;人体运动;运动生物力学;康复理论
1 引言
人有能力完成各种精巧的动作,例如:走钢丝、花样跳伞、骑独轮车等,人体运动控制的研究目的在于了解隐藏在人类与众不同的能够完成精准动作能力背后的机制。人体运动控制领域的研究先后纳入了物理学、解剖学、生理学、心理学、工程学、神经学等学科的内容,是一个综合的领域,国外关于人体运动控制的系统研究始于19世纪早期并一直延续到今天。
人体运动控制的研究从对人体运动外在的机械运动的研究逐渐深入到神经控制层次的研究,尤其神经生理学基于电信号对中枢及周围神经系统的研究提供了人体活动与相关神经部位的信息,为以后的控制模型的研究提供了理论基础,其中,由Laszlo实施的,利用现代技术进行的追踪性研究从心理物理学的角度对人体的运动进行了阐述[42]。1995年,Neilson 等人首次应用系统工程学的方法对追踪任务相关的人体运动进行了解读[60]。20世纪末,研究者试图通过寻找直接的人体运动控制的神经生理学证据对心理物理学模型给予支持,以达到整合人体运动控制的物理属性与心理属性的目的[79,81,82]。在进行上述人体运动控制理论的结构性研究的同时,研究者们也对神经系统中信息的处理方式进行了研究,提出了统合心理观察、神经心理学数据和系统工程学视角的概念化框架理论—适应性模型理论(Adaptive Model Theory,AMT)并进行了评估[60,62,63,68]。
2 人体运动中的控制问题
2.1 冗余控制与富余控制
冗余控制的概念在不同的领域有着不同的侧重,如,在工程领域中,冗余控制的概念是采用一定或成倍量的设备或元器件的方式组成控制系统来参加控制。当某一设备或元器件发生故障而损坏时,可以通过硬件、软件或人为方式相互切换后备设备或元器件替代因故障而损坏的设备或元器件保持系统正常工作,使控制设备因意外而导致的停机损失降到最低[1]。在通信理论中,冗余是指以多于所传递内容的信息量传送信息。在语义学中,冗余指一词双意或一词多义。冗余的优点在于使得所要控制或表达的内容具有了相当的弹性,从而具有了相当的稳定性。在人体的运动控制领域,对人体进行的从人体整体运动到某一个肢体环节运动的各种层面上的描述,都会涉及冗余控制的问题,冗余控制的解算是一个核心的问题。
在保持身体部分(躯干、下肢、头部等)静止的状态下移动一侧上肢控制食指达到空间中的某一个位置的问题中[47],食指所要达到的空间位置点在笛卡尔坐标系中可以用3个坐标点{X,Y,Z}进行描述,而在整个过程中人体所要协调控制的上肢自由度至少为7个,即描述肩关节的3个角度{α1,α2,α3},描述肘关节运动的{α4},描述腕关节运动的{α5,α6}以及描述前臂在腕关节和肘关节处联合运动的{α7}即前臂的旋前运动与旋后运动。上述问题等价于解算方程组(1),式中x、y、z为食指末端的空间位置坐标,αi为描述关节运动的角度,ki为由上肢解剖特点所决定的关节角度与食指末端位置的映射关系。在上述数学描述中为了完成移动任务{X,Y,Z}而能够利用的{α1,α2,α3,α4,α5,α6,α7}的组合是无限的。在人体运动中每一次的过程都是确定的,人体遵循何种规则完成这种控制是未知的,这种规则涉及生理学(肌肉、神经等)、心理学(意识、认知等)、解剖学(骨骼、肌肉等)等学科。

人体运动控制中的冗余控制问题远超机械概念上的自由度的概念,总数大约5 000 000从脑干发出的纤维与总数在150 000左右的运动神经元相联系,从而控制700余块肌肉[64]。人体运动系统中自由度的总数在100~150之间[61]。人体通过这些复杂的生理、解剖结构对3维空间中的运动实施控制,其冗余度极其庞大。
富余控制是由冗余控制的解算延伸出的概念,Gelfand和Tsetlin 于1966年提出基于经济的原则纳入最少数目的自变量从而避免冗余控制的问题[30]。但是,这样做的缺点在于使得生物体过于机械化从而丧失了生物体的优势。Gelfand和Latas后来在文章中提出纳入更多的自变量以使得系统更具有弹性,认为不同的变量以权重组成了不同的模式从而控制着系统,强调不要把系统限制于唯一的解上[47,29]。

图 1上肢控制示意图Figure1.An Illustration of the Kinematic Redundancy of the Human Arm
人体作为一种冗余控制的客体使得人体的运动在神经支配下表现出极强的灵活性(完成各种精巧动作的能力)和适应性(在受到意外干扰时保持适应的能力),同时,冗余控制的解算也变成了人体运动控制中的核心问题。
2.2 运动变异
人体运动的变异是由人体运动系统的冗余引起的,由于人体的运动控制是冗余控制,所以当完成同样的任务时可以采用不同的方式。运动变异的问题最早由Bernstein在1926年提出[11],如图2所示,他观察到当一个技术熟练的铁匠在保持身体其余部分不同的状态下重复的敲打铁块时,每次完成动作的轨迹精确的来看都是不重合的。这表明,当人在完成重复的动作时并不存在一个严格固定的模式,而是有弹性的。

图 2 铁匠上肢控制示意图Figure2.The Illustration of the Arm Controlling when a Blacksmith Hits an Object with the Tip of the Hammer
产生运动变异的原因存在两种观点[49]:1)人体运动系统中存在噪音,运动变异是一种不完美的控制。2)运动变异是人体运动系统的控制策略。对运动变异的研究从最初的对变异性的简单描述发展到利用非控制流形[75](UCM, Uncontrolled Manifold)、主成分分析法[43,17](PCA,Principle Component Analysis)、独立成分分析法[36,16](ICA,Independent Component Analysis)等技术进行分析,进一步揭示了隐藏在运动变异之后的生理学机制。UCM 方法是将变异程度分解于两个无关空间——控制空间(以铁匠问题为例是由肩关节、肘关节以及腕关节所构成)与其正交补空间(代表与控制无关的噪声空间),UCM方法揭示了中枢在运动变异中所起到的作用,利用UCM方法可以对运动变异中的受神经控制的部分进行解释,可以用来解释动作学习的原理,同UCM类似的方法还有TNC[56-58](Tolerance,Noise,Control)方法。Winter应用PCA方法研究了16条腿的肌肉的肌电信号,发现可以将其分为4个基本模式[85]。Ivanenko利用PCA方法对多个步态周期采集的25块肌肉的肌电信号进行分析,发现25块肌肉共享5种模式,通过5种模式的组合可以解释总体差异的90%[37]。
3 经典的人体运动控制假说
3.1 基于信息控制理论的假说 (动作编码)
随着对生物的中枢神经系统和外周神经系统研究的加深,基于控制理论的人体运动假说逐渐完善。最早的人体运动控制假说-反应链假说(response-chaining)由James(1891)提出[38],他认为,外部刺激激发第1个动作,随后产生的信息被本体感觉接受,从而通过反馈引起第2个动作,继而引起整个反应链的响应。在James的假说中不需要认知注意的参与,因为后一个动作是被前一个动作激发的,这样一旦整个链条激活便会执行到底。这种控制模式是开环的称之为前馈执行。
在人体运动中存在一种闭环的控制模式,Adams(1971)提出[4],在人体运动中通过反馈方式最大可能的降低实际运动与“认知”过程中的期望运动之间的差异。Adams的理论很好地解释了人体运动的调节,尤其是对于慢速运动的修正。后来Adams的理论受到一些实验事实的挑战,如,在人们看到东西然后用手去取的视觉反馈中,手到脑的信息传递时间需要150~200 ms的延迟时间大大超出手移动的时间,无法进行比较。虽然如此,反馈机制在人体运动的控制中仍很重要[39]。
3.2 人体运动控制中的层级结构
Craik(1948)最早提出把人脑看作可输入信息并输出执行的计算机[18],而人们在实际中观察到神经信号的传导是脉冲式的而不是连续的支持了他的想法。Schmidt认为[73],人体运动中的信息处理是层级式的,信息是从高的层级向低的层级传送,越高的层级需要越多的注意力,如图3所示的信息控制模型中,占据最高层级的是“认知”,需要耗费最多的注意力,与心理状态密切相关。在“认知”以后需要传递指令给运动系统从而实现一个具体的动作,当运动系统接收到信息后由运动神经中枢发出信息,控制对应的骨骼肌肉产生运动,运动的实施效果又通过反馈机制传递信息到“认知”层级和运动控制层级,对具体动作进行调控。

图 3人体运动控制中的层级结构简图[21]Figure3.Simplif i ed Hierarchy in a Typical Information Processing Model
动作编码认为,人体运动中的控制信息通过层级结构以反馈和前馈的方式得到实现,在此基础上,Erlhagen 和Schoner, Schoner 和 Thelen提出了动态神经元场理论[23,76],试图解释神经元中的信息处理过程。动作编码理论使得人体运动的机制变得清晰,但却存在一个不可回避的问题,在人的运动中存在无数的变量(自由度)需要同时进行控制,人体不可能同时细节化处理所有的变量。
3.3 基于物理过程的假说
3.3.1 平衡点假说
1.阈值控制理论:在人体中神经元细胞以及细胞膜的信号传递(极化,去极化)都具有阈值的一面,这使得在人体运动控制系统中人体并不是对所有的信号产生应答,而是表现出一个综合的效应。如图 4 所示,IA是传入神经的信号,IC是中枢神经的信号,N是依赖于 IA的输出信号。当IC小于阈值Th时,输出表现为 N;当IC大于阈值Th时,输出不表现为N。通过阈值的设定,神经元完成了对不同信息的综合。
解决人体运动的冗余问题最直接的方法是限定各种优化标准,如,最小能耗方程、最优轨迹等,阈值控制是解决人体运动的冗余问题的另一种可能途径。
研究证明,大脑皮质脊髓束、网状脊髓束、红核脊髓束、前庭脊髓束都可以对肢体的阈值位置(把某一个肢体的位置看作动作的阈值)进行重置[25,69]。 阈值控制理论把人体运动的控制看作神经系统对不同阈值的重置,例如,肌肉在肢体处于某一个位置时肌肉的活动并不明显,但是,在这个位置的肌肉已经对可能遭受的干扰及运动做好了准备,其状态类似系统的伺服机制[7,27]。阈值控制本身并没有去解算冗余控制问题,而是通过阈值的设置以及重置从而对人体运动系统中近乎无限的运动的可能性进行限制,在阈值控制的理论框架内可以对人的抓握活动、坐-起以及行走做出合理的解释。

图 4阈值控制理论示意图[49]Figure4.Illustration of Threshold Theory[49]
2.平衡点假说:在物理上,平衡点指合力以及合力矩为零的状态,在人体运动中平衡点假说侧重于阐述物体的运动是在不同的平衡点之间的转化,平衡点假说是由Feldman(1966)提出的[26]。Feldman最早提出的版本称为λ-平衡点假说(图 5 ),λ1和λ2与不同的神经兴奋性相关联,确定λ相当于确定了一条肌肉的力-长度曲线,从λ1到λ2的变化将导致肌肉长度的变化,即L1到L2。Bizzi等(1976)提出肌肉工作时具有类似的表现,称之为α-平衡点假说[13](图 6),肌肉激活程度α的改变将导致肌肉力、肌肉长度的综合变化,继而使得肌肉处于不同的平衡状态EP1-3从而表现为肌肉的活动,与λ-平衡点假说相比,α-平衡点假说更适用于肌肉。
根据平衡点假说,一旦肢体的最终位置被神经系统确认,中途过程将不受干扰的自动完成,所以,基于平衡点假说可以设计出简单、稳定的控制系统。在后来,平衡点假说遭受到了越来越多的质疑[42,83],根据平衡点假说,随着速度的增加肌肉将表现出很大的粘弹阻力,但是,Gomi和Kawato (1996)的实验说明,关节硬度并没有随着移动速度的增加有明显的改变甚至降低[33]。Shadmehr和Mussa-Ivaldi (1994)将受试者置于阻尼力场中,当撤去力场后即刻,受试者表现出和力场存在时相同的镜面效应,这与平衡点假说不一致,但是却符合内部模型理论[80]。

图 5 λ-平衡点假说示意图[47]Figure 5.Equilibrium-point Hypothesis-λ Model
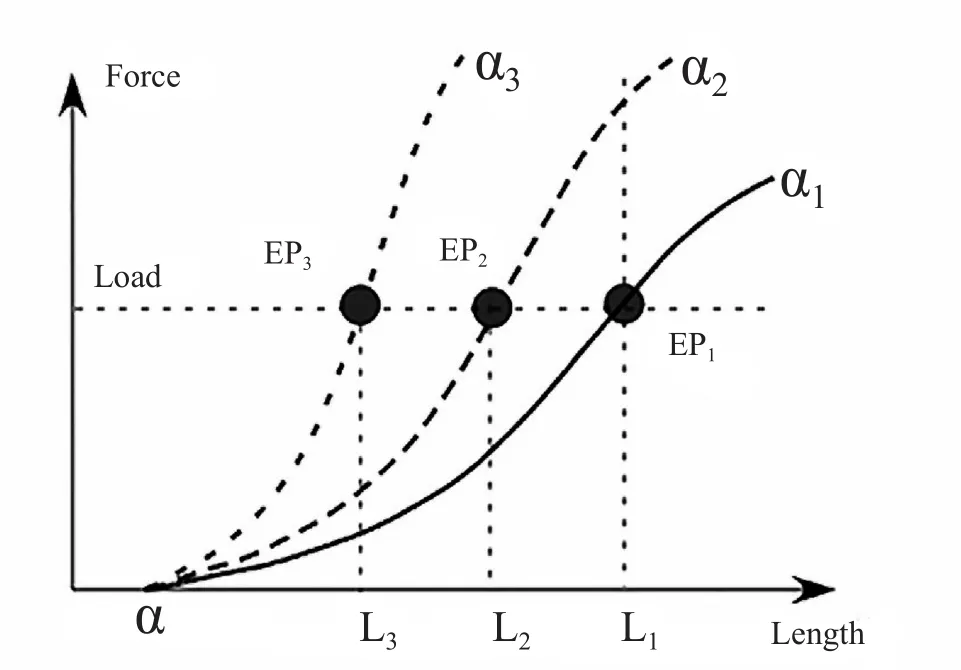
图 6 α-平衡点假说示意图[47]Figure 6.Equilibrium-point Hypothesis-α Model
随着研究的发展,平衡点假说又重新被重视起来,平衡点假说一开始被用来解释单关节的运动,如肘关节、膝关节。现在,平衡点假说被发展为解释多关节的复杂运动。平衡点假说能够很好地帮助理解中枢神经系统受到损伤的患者表现出的运动控制的不足。当患者试图进行主动活动时,关节活动范围内紧张性牵张反射的阈值的改变被证明(tonic stretch ref l ex threshold ,TSRT)与不正常的肌肉激活模式——肌肉的过度共同激活——相关[50]。在临床上可以利用平衡点假说判断运动控制中的不足,通过改善紧张性牵张反射的阈值从而改善患者的运动能力。
3.3.2 对照设置假说
对照设置假说是平衡点假说的一种发展,此假说被用来对有多肌肉群参与,多关节包括全身动员的运动(比如跑步)进行描述[24]。对照设置(Referent conf i guration,RC)按照定义是这样一种设置,当处于此种设置时,所有的肌肉都处于激活的阈值状态,此时的肌肉长度等于由λ决定的肌肉长度(图 5)。对照设置假说如图 7所示:N1代表神经元细胞组中的细胞膜活化水平,Central input 代表外部条件,决定了RC。对照设置假说认为,RC与AC(Actual conf i guration,AC)的差别导致了肌肉激活产生肌肉力,从而维持姿势与产生运动。

图 7 对照设置假说示意图[49]Figure 7.Illustration of Referent Conf i guration Hypothesis
对照设置假说还可以用来解释人是如何抓住一只杯子的,图8表示的是一个截面图,TH表示拇指,VF代表其余4指(I-食指,M-中指,R-无名指,L-小指),L 表示外力。
整个过程需要人对3种对照设置进行控制:
1.拇指与其余4指的距离,APREF
2.杯子的纵向位置,ZREF
3.杯子的方向αREF
人通过控制拇指与其余4指的距离从而生成手与杯壁之间的正压力Fn,继而产生向上的摩擦力Ft,从而抵消掉杯子受到的向下的外力,通过控制杯子的方向α从而保证水不会洒出来,通过控制杯子的纵向位置Z保证杯子固定在一个位置。根据对照设置假说,与控制一个水杯的原理相似,人通过一系列的设置从而完成各种复杂的动作,而人从婴儿到成年的神经发育过程重要的一面有可能是各种对照设置数量的增多和完善,从而导致了人的动作趋向成熟[19]。

图 8人握杯子时的示意图[46]Figure8.Referent Con fi guration Components for a Static Prehensive Task
3.3.3 动态行为理论
动态行为理论[40]的核心概念基于Bernstein(1967)对协调性的研究[34]和Gibson(1979)对感知的研究[31]。动态行为理论对传统的以信息处理为重点的动作控制理论是一种彻底的颠覆,因而引起了巨大的争议。
动态行为理论的观点来自于在物理领域对力学或协同作用更广泛的研究,试图把人的运动看作与客观世界的内化——内部模式、动作编程或者记忆——完全无关,人的运动被看作基于人体与周围环境相互作用而产生的自组织行为。例如,动物步行的越来越快,到达某点时,突然转换为小跑。当动物继续快速移动达到第2点时,则转换为疾跑。动态行为理论认为,新的动作的出现是由于在多系统中有一个发生了关键性的改变,被称为“控制参数”。与侧重于信息处理的其他运动控制理论不同,在动态行为理论中,运动学的参数占据了主导地位而不是神经系统的信息处理过程[3]。
用动态行为理论的观点来描述动作的一个重要内容是吸引状态。吸引状态即生物平时喜欢的状态,如行进速度,当以这种速度行进时是最高效节能的,其对康复实践的指导意义在于可以利用人自身的物理特性进行训练,如,通过对运动失控患者的速度进行控制从而帮助虚弱的患者更加容易的活动[2]。
4 人体运动控制的计算框架
4.1 人体运动的黎曼流形计算框架
依照生物力学的观点,人体是一个由多个环节组成的具备多生物力学自由度的物理系统。在人体运动的实际状态中,参与运动的各个要素之间以及各个要素与环境的交互严格来说不存在简单的线性关系,它们之间的关系往往是非线性的。黎曼几何空间不同于欧几里得几何空间,欧几里得几何空间是黎曼空间中截面曲率为常数的特例。黎曼流形是非线性的空间流形,通过黎曼流形天然的更加方便对人体运动进行研究。Biess (2013)指出[12],在研究人体运动中使用的坐标类型对研究结果存在影响,因为各向异性、协矩阵的正交性都是坐标系依赖的。在黎曼几何的架构中,由于在不同的坐标系之间进行转化的张量方程是保持不变的,例如,在一个坐标体系中张量方程等于零,那么,在剩余的所有坐标体系中张量方程都等于零,这样,对人体运动的描述可以独立于坐标系进行,从而不受不同坐标体系的影响。Arimoto (2010)对多关节机器人的研究表明,由运动自由度张成的空间可以用黎曼空间表示,并且黎曼度量等于机器人的质量-惯性矩阵[6]。
为了研究整个人体在具体环境中的运动,3个光滑的流形是必需的。1)根据Neilson的研究[65-67],人体的运动可以用110个元素运动代表,所以第1个流形是包含110个元素的流形Ɵ,流形中的点代表人体姿态。2)三维流形P中的每一个点代表头部在外部坐标系中的位置。3)三维流形O中的点的坐标是头部的方向矢量,代表头部的旋转姿态。头部在外部空间中的活动会形成O、P、Ɵ 3个流形之间的映射,如,芭蕾舞者头部的运动引起全身姿态的改变。笛卡尔外积C = Ɵ×P×O定义了一个116维度的空间C,其中的点c∈C的坐标(c1,...,c116)在外部坐标系中唯一定义了身体姿态,头部的位置、方向。C称之为人体运动的流形空间,点c∈C的改变代表着人体姿态的改变,而运动流形中依赖于时间的轨迹c(t)则代表了人体的运动。曲线c(t)的切空间代表其速度变化,记做TcC,TcC的切空间代表其加速度的变化,记做TvTcC。C的黎曼度量J(c)对应于质量-惯性矩阵,据Bullo和Lewis的研究[51],J(c)具有动力学-能量的属性,一旦获知人体运动的黎曼度量J(c)便可以计算得出TvTcC、TcC,从而确定人体的运动。
4.2 运动变异的分解
4.2.1 非控制流形分解框架
运动变异是广泛存在的一种事实,在Bernstein 对铁匠打铁轨迹的研究中,他观察到上肢运动的变异远远大于锤子的运动变异[15],由此他做出动作不会机械的、严格不变地重复的推论,但是,这种变异是一种来源于人体运动系统的噪声还是反映了人体运动系统的灵活性、适应性未有定论。Schöner和Scholz提出了一种非控制流形的理论(Uncontrolled Manifold,UCM),用以区别运动变异是来源于运动控制的噪音还是运动系统灵活性的体现[75,77]。在一个由n个自由度控制的实现在m个自由度空间中的目标(n>m),存在一个与移动无关的(n-m)维度的空间,称之为非控制流形[75],非控制流形理论认为,在人体的运动控制中,作为控制部的神经系统没有必要对所有的运动变异,进行控制,对于位于非控制流形中的变异,由于它们与最终的运动无关,所以没有必要进行控制,只有位于与非控制流形正交补空间中的变异才有必要加以控制。
UCM与“自运动流形”[22]、“无运动流形”[41]、“等效运动流形”[20]在本质上是相同的。如图9所示,在利用两个手指(F1,F2)产生一个目标力FTASK任务中,其控制的自由度为2,其目标自由度为1,故其UCM空间的维度为1,即图中所示虚线。当处于UCM正交补空间中的运动变异分量VBAD大于处于UCM空间中的运动变异分量VGOOD时,说明需要神经系统费力加以控制,反之则说明无须过多的神经控制。运动技能学习的过程就是从需要神经系统费力控制到无须费力控制直至动作自动化的过程。
UCM分析可区分有神经功能障碍人群和健康人群的运动协同模式。有研究显示,相对于年轻健康人群,一些人群如老年健康群体、患有唐氏综合征或帕金森病或小脑有疾病的人群,由于运动协调能力受损,表现出相对更少的VGOOD变异[34,71]。

图 9变异分量示意图Figure 9.Illustration of Decomposition of Variability in UCM Space
在控制单侧上肢移动末端手指的任务中,以简单模型为例,控制的自由度包括肩关节的3个旋转、肘关节的屈伸、腕关节的屈伸共计5个自由度,其控制目标为手指末端的空间位置,为3个自由度。控制空间和目标空间的映射为X=Jα,X代表目标空间即手指末端的位置向量,α代表控制空间的肩、肘、腕的角度的组合代表的向量,J为雅克比矩阵。JX的零子空间JX=0即为UCM空间[75,74,44]。
4.2.2 容错-噪声-共变分解框架
不同于非控制流形理论在控制空间上对运动变异的分解,TNC是在目标空间上对运动变异进行分解。以投掷飞镖为例,如图10所示,A、B、C、D、E代表5组练习过程中的测试,每组包含9次测试。灰度代表偏离靶心的程度,白色代表击中靶心。根据物理原理,当我们知道了投出时刻飞镖的速度以及出手角度、出手位置,那飞镖的投掷轨迹便被唯一的确定了。假设每次都是在同一位置出手,对飞镖轨迹进行分析,我们便会得到由出手速度和出手角度张成的二维空间,对其目标空间的分析结果如图10所示。
1. 容错 。A组测试是最初阶段的测试,在这个阶段由于技能不熟练,所有的飞镖都偏离靶心。随着练习的进行,在B组中有些测试能够击中靶心,阶段C与阶段B的区别在于随着练习的进行,在C阶段,即使受试者没有刻意地去试图击中靶心,但是能够更容易的击中靶心,因为在C阶段有更多的角度—速度组合可资利用(C阶段所处位置的白色区域比B阶段所处位置的白色区域要宽),表现出在C阶段具有更强的容错能力。Müller 和 Sternad给出了容错能力的计算方法[59]。
2. 噪声。19世纪末,Fullerton、Cattell和Woodworth 分别进行了一项画直线的研究[28,86],结果显示,就算在如此简单的任务中,运动变异或者噪声也是不可避免的,尽管运动变异通过练习可以减小,但是无法完全消除。如图10所示,从阶段C到阶段D的过程中,随着练习的增加,受试者的表现更加稳定便是噪声减少的原因。
3. 共变。随着练习的深入,受试者的技能更加纯熟(阶段D到阶段E),此时不佳的出手角度可以通过出手速度进行补偿从而命中靶心,反过来,不佳的出手速度可以通过对出手角度的调整得到补偿从而同样命中靶心。Müller和Sternad给出了计算共变的方法[59]。

图 10在目标空间上对飞镖轨迹的变异进行分解的结果图示Figure10.The Decomposition of Variability of Throwing Learning Task in Task Space
容错、噪声、共变3个因素之间互不重叠,完整的解释了在目标空间的运动变异。与UCM框架相比,TNC分解有着自己的优点和缺点。UCM框架进行的分析基于雅克比矩阵的零空间,其分析是线性的,如果对一个非线性比较强的空间应用线性理论,很明显会造成错误。但是,由于UCM理论是对控制空间进行分析,其结果可以对生理过程(比如神经系统、反射、反馈)等进行分析[45,53]。
4.3 适应性模型理论AMT
AMT 是一种综合的基于逆动力学计算的人体运动控制模型,概念来源于自适应控制理论,其核心假设是大脑能够根据周围环境计算和储存逆动力学参数,并利用计算结果指导身体在三维空间中产生平滑、协调的运动。AMT中的逆动力学模型具有对身体非线性运动信息的输入和输出的稳定适应性[62]。AMT具有基于神经生理的层级结构,处于高层级的结构(认知等 )可以以最优化的方式组织底层级结构的表现。
Neilson提出的AMT模型结构(图11),对应于人体的控制过程,此结构分为脑、身体、外部环境3个主要区域,3个区域间的信息传递结构是正交的,采用的是格拉姆施密特正交化(Gram-Schmidt orthogonalisation technique),MSC(肌肉控制系统muscle control system),BM(生物力学系统biomechanical system),E(外部系统external system)。利用此框架可以对人体的运动进行模拟并具有相当的可靠性,此框架充分考虑了人体运动的生理结构,但相对于实际的灵活、复杂、多态的人体运动来说,此框架还是显得过于机械。

图 11 Neilson等提出的适应性模型框架图[63]Figure11.Summary of Information Flow for Adaptive Model Theory
5 人体运动控制理论的实践应用
传统上人们一直认为,前馈控制与反馈控制在产生动作的作用上是二分对立的,人们对动作产生的主要控制机制一直在前馈和反馈之间摇摆,然而,Seidler等(2004)的研究[78]表明,前馈控制与反馈控制之间的关系是相互延续的而非二分的存在,大脑皮质与皮质下结构共同完成动作的功能,Wanger和Smith (2008)的研究[84]表明,前馈控制与反馈控制在某种程度上共享同种内部模式。Bastos等(2015)[8]对视觉皮质的研究表明,与传统的理论认为反馈是通过对传输过程中的前馈信号进行调节从而实现与前馈协同作用的过程不同,前馈控制与反馈控制使用不同的信号频率进行信息交换,这就使得前馈控制与反馈控制之间的交互有着更为复杂的结构。Yousif和Joern (2015)[87]对恒定力场与可变力场中的人体运动的研究结果表明,前馈控制与反馈控制在结构上至少是可以部分分离的。在这些研究的基础上,Pan等(2016)[70]提出了更为复杂的综合了前馈控制与反馈控制的模型,相对于传统模型在简化结构的同时具备了更强的抗干扰能力。对前馈控制与反馈控制关系认识的深入也指导康复师设计更为合理的康复训练计划。
人体运动控制理论对康复及运动训练有着重要的影响,应用运动控制理论模拟人体控制的康复用医疗机器人可以对病人适时地施加电刺激从而帮助病人完成动作,促进病人的康复[5,72]。Bobath康复训练方法就是基于反射的动作控制理念进行设计的,其核心是针对性的加强有益的反射,抑制不利于动作的反射。临床上会基于人体运动控制的层级结构理论,通过对病人反射功能的分析从而推断病人的神经功能状况,也会依据对病人运动功能的分析从而得出其神经发育程度及功能现状。根据运动控制理论,对病人的康复治疗应当关注在主要的动作上面而不是对单独肌肉的训练上,这样才能建立新的神经控制模式,运动控制理论还提示,康复治疗师对病人的康复不应单独地关注某一个系统与某一个部位,而应该从多个系统综合的角度对病人进行康复与评估;康复师可以利用对称或相似的动作实施康复训练,这种方法可以影响相应的神经控制中枢,从而有利于病人的康复[14]。
在体育运动中,运动相关损伤的机制是非常复杂的,包含内部与外部各种因素,有学者从运动控制的角度设计了运动感觉训练方法对运动员进行干预,结果表明,这种训练方法能够有效地降低损伤的发生率[32]。对于前交叉韧带的损伤来说,损伤史是其再次损伤的一个重要因素,从运动控制的神经肌肉训练角度出发,学者们利用反馈技术改善运动员的技术水平延长了运动寿命,在长期来看,也降低了前交叉韧带的损伤率[9,10]。在运动中,运动技能除了受生理学及生物力学因素的影响外,认知对其的影响也非常重要,优秀运动员与一般运动员在感知觉的能力上存在明显差异[52]。利用运动控制训练的生物反馈系统可以提高足球运动员的运动能力,同时降低罹患损伤的风险[35,54],而通过对高尔夫运动员运动控制能力的分析可以帮助其进行动作控制训练提高其成绩[55]。
6 小结
人体的运动是复杂的,对人体运动控制的研究有交叉综合的特点,涉及数学、物理、计算科学、工程学、解剖学、生理学等多门学科,对人体运动控制中现象的描述会借用各个学科的术语。由于其繁杂的研究,到现在为止,甚至连术语体系都不完整统一,但是,运动控制的研究是非常重要的。人体运动控制的研究从过去单一的运动生物力学对独立关节、肌肉的研究,发展到现在对人体运动整体包括神经生理机制的研究,从简单的关节角度、角速度的计算,发展到由神经支配控制的正动力学及正、逆动力学相结合的综合运用,人体运动控制研究的计算手段、评估方法变得越来越丰富。它让我们能够更深入的了解人体运动的神经生理、运动损伤机制、竞技过程中的运动表现。通过对人体运动控制过程的深入研究,可以帮助有运动障碍的人群制定针对性的康复计划或开发适当的辅助器械,帮助运动员制定更有效的训练方法以提高运动成绩。
[1] 陈子平. 浅谈控制系统冗余控制的实现[J]. 自动化仪表,2005,26(9):4-6.
[2] 沙姆韦•库克. 运动控制原理与实践[M]. 北京:人民卫生出版社,2009.
[3] ABERNETHY B,SPARROW W A. The rise and fall of dominant paradigms in motor behaviour research.[J]. Adv Psychol,1992,84:3-45.
[4] ADAMS J A. A closed-loop theory of motor learning[J]. J motor behav,1971,3(2):111-150.
[5] ALIBEJI N A,KIRSCH N A,SHARMA N. A muscle synergy-inspired adaptive control scheme for a hybrid walking neuroprosthesis[J]. Fronts Bioeng Biotech,2015,3.
[6] ARIMOTO S. Modeling and Control of Multi-Body Mechanical Systems:Part I A Riemannian Geometry Approach[M]// Advances in the Theory of Control,Signals and Systems with Physical Modeling. Springer Berlin Heidelberg,2010:3-16.
[7] ASATRYAN D G,FELDMAN A G. Biophysics of complex systems and mathematical models. Functional tuning of nervous system with control of movement or maintenance of a steady posture. I. Mechanographic analysis of the work of the joint on execution of a postural task[J]. BioFiz,1965,10(5):837-846.
[8] BASTOS A M,VEZOLI J,BOSMAN C A,et al. Visual areas exert feedforward and feedback inf l uences through distinct frequency channels[J]. Neuron,2015,85(2):390-401.
[9] BENJAMINSE A,GOKELER A,DOWLING A V,et al. Optimization of the anterior cruciate ligament injury prevention paradigm:novel feedback techniques to enhance motor learning and reduce injury risk[J]. J orthop Sports Phys Ther,2015,45(3):170-182.
[10] BENJAMINSE A,WELLING W,OTTEN B,et al. Novel methods of instruction in ACL injury prevention programs,a systematic review[J]. Physi Ther Sport,2015,16(2):176-186.
[11] BERNSTEIN N A. General biomechanics[J]. Izd. VTsSPS,1926.
[12] BIESS A. Shaping of arm conf i guration space by prescription of non-Euclidean metrics with applications to human motor control.[J]. Phys Rev E Stat Nonlin Soft Matter Phys,2013,87(1):309-318.
[13] BIZZI E,POLIT A,MORASSO P. Mechanisms underlying achievement of fi nal head position[J]. J Neurophysiol,1976,39(2):435-444.
[14] CANO-DE-LA-CUERDA R,MOLERO-SÁNCHEZ A,CARRATALÁ-TEJADA M,et al. Theories and control models and motor learning:clinical applications in neurorehabilitation[J]. Neurología,2015,30(1):32-41.
[15] CARPENTER M B. The co-ordination and regulation of movements[J]. Neuropathol Exp Neurol,1968,27(2):348.
[16] CELONE K A,CALHOUN V D,DICKERSON B C,et al. Alterations in memory networks in mild cognitive impairment and Alzheimer’s disease:an independent component analysis[J]. Neurosci,2006,26(40):10222-10231.
[17] CHAPIN J K,MOXON K A,MARKOWITZ R S,et al. Real-time control of a robot arm using simultaneously recorded neurons in the motor cortex[J]. Nat Neurosci,1999,2(7):664-670.
[18] CRAIK K J W. Theory of the human operator in control systems[J]. Bri J Psychol,1947,38(2):56-61.
[19] CROIX M D S,KORFF T. Paediatric Biomechanics and Motor Control:Theory and Application[M]. Routledge,2013.
[20] CUSUMANO J P,CESARI P. Body-goal variability mapping in an aiming task[J]. Bio Lcybern,2006,94(5):367-379.
[21] DAVIDSON P R. Computational modelling of the human motor control system:Nonlinear enhancement of the adaptive model theory through simulation and experiment[D]. University of Canterbury Electrical & Comput Enging,2001.
[22] DEMERS D,KREUTZ-DELGADO K. Canonical parameterization of excess motor degrees of freedom with self-organizing maps[J]. IEEE Trans Neural Netw,1996,7(1):43-55.
[23] ERLHAGEN W,SCHÖNER G. Dynamic fi eld theory of move-ment preparation.[J]. Psychological Rev,2002,109(3):545.
[24] FELDMAN A G,LEVIN M F. The origin and use of positional frames of reference in motor control[J]. Behav brain sci,1995,18(4):723-744.
[25] FELDMAN A G. Once more on the equilibrium-point hypothesis(lambda model) for motor control.[J]. J Motor Behav,1986,18(1):17.
[26] FELDMAN A G. Functional tuning of the nervous system with control of movement or maintenance of a steady posture. II. Controllable parameters of the muscle[J]. Biophys,1966,11:565-578.
[27] FOISY M,FELDMAN A G. Threshold control of arm posture and movement adaptation to load[J]. Exp Brain Res,2006,175(4):726-744.
[28] FULLERTON G S,CATTELL J M K. On the Perception of Small Differences:With Special Reference to the Extent,Force and Time of Movement[M]. University of Pennsylvania Press,1890.
[29] GELFAND I M,LATASH M L. On the problem of adequate language in biology[J]. Prog Motor Control,2002,2:209-228.
[30] GELFAND I M, TSETLIN M L.On mathematical modeling of the mechanisms of the central nervous system.[M/C]// GURFINKEL I M,FOMIN V S,TSETLIN S V(Eds.).Models of the Structural-Functional Organization of Certain Biological Systems.Gelfand,Nauka,Moscow, 1966: 9-26.
[31] GIBSON B J J. The Ecological Approach to Visual Perception[J]. J Br Soc Phenomenol,1979,13(1):98-99.
[32] GOLLHOFER A,GRANACHER U,TAUBE W,et al. Motor control and injury prevention[J]. Deutsche Z Für Sportmedizin,2006,57(11):266-270.
[33] GOMI H,KAWATO M. Learning control for a closed loop system using feedback-error-learning[C]//Decision and Control. Proceedings of the 29th IEEE Conference on. IEEE,1990:3289-3294.
[34] HARMARK T,OBERS N A. Uncontrolled manifold analysis of single trials during multi-f i nger force production by persons with and without Down syndrome.[J]. Exp Brain Res,2003,153(1):45-58.
[35] HIDES J A,STANTON W R,MENDIS M D,et al. Effect of motor control training on muscle size and football games missed from injury[J]. Med Sci Sports Exerc,2012,44(6):1141.
[36] IVANENKO Y P,POPPELE R E,LACQUANITI F. Motor control programs and walking[J]. Neuroscientist,2006,12(4):339-348.
[37] IVANENKO Y P,POPPELE R E,LACQUANITI F. Five basic muscle activation patterns account for muscle activity during human locomotion[J]. Physiol,2004,556(1):267-282.
[38] JAMES W. The Principles of Psychology[M]. Read Books Ltd,2013.
[39] KAWATO M. Internal models for motor control and trajectory planning[J]. Curr Opin Neurobiol,1999,9(6):718-727.
[40] KELSO S J A. Human Motor Behavior:An Introduction[M]. 1982.
[41] LABOISSIÈRE R,OSTRY D J,FELDMAN A G. The control of multi-muscle systems:human jaw and hyoid movements[J]. Biol cybern 1996,74(4):373-384.
[42] LASZLO J I. Motor control and learning:how far do the experimental tasks restrict our theoretical insight?[J]. Adv Psychol,1992,84:47-79.
[43] LATASH M L,SCHOLZ J P,SCHÖNER G. Motor control strategies revealed in the structure of motor variability[J]. Exerc sport Sci Rev,2002,30(1):26-31.
[44] LATASH M L,SCHOLZ J P,SCHÖNER G. Toward a new theory of motor synergies.[J]. Motor Control,2007,11(3):276-308.
[45] LATASH M L,SHIM J K,SMILGA A V,et al. A central back-coupling hypothesis on the organization of motor synergies:a physical metaphor and a neural model[J]. Biol Cybern,2005,92(3):186-191.
[46] LATASH M L,ZATSIORSKY V. Biomechanics and Motor Control[M].2016.
[47] LATASH M L. The bliss (not the problem) of motor abundance (not redundancy)[J]. Exp brain Res,2012,217(1):1-5.
[48] LATASH M L. Motor synergies and the equilibrium-point hypothesis[J]. Motor control,2010,14(3):294-322.
[49] LATASH MARK L. Fundamentals of Motor Control[M]. Elsevier/Academic Press,2012.
[50] LEVIN M F. Sensorimotor def i cits in patients with central nervous system lesions:explanations based on the λ model of motor control[J]. Hum mov Sci,2000,19(1):107-137.
[51] MARTINEZ S. Geometric Control of Mechanical Systems[M]. Springer,2005.
[52] MANN D T Y,WILLIAMS A M,WARD P,et al. Perceptual-cognitive expertise in sport:a meta-analysis.[J]. J Sport Exerc Psychol,2007,29(4):457.
[53] MARTIN V,SCHOLZ J P,SCHÖNER G. Redundancy,self-motion,and motor control[J]. Neural Comput,2009,21(5):1371-1414.
[54] MENDIS M D,HIDES J A. Effect of motor control training on hip muscles in elite football players with and without low back pain[J]. Sci Med Sport,2016,19(11):866-871.
[55] MORRISON A,MCGRATH D,WALLACE E S. Motor abundance and control structure in the golf swing[J]. Hum Mov Sci,2016,46:129.
[56] MÜLLER H,STERNAD D. A randomization method for the calculation of covariation in multiple nonlinear relations:illustrated with the example of goal-directed movements[J]. Biol Cybern,2003,89(1):22-33.
[57] MÜLLER H,STERNAD D. Accuracy and variability in goal oriented movements-decomposing gender differences in children[J]. Hum Kinet,2004,12:31-50.
[58] MÜLLER H,STERNAD D. Decomposition of variability in the execution of goal-oriented tasks:three components of skill improvement[J]. Exp Psychol Hum Percept Perform,2004,30(1):212.
[59] MÜLLER H,STERNAD D. A randomization method for the calculation of covariation in multiple nonlinear relations:illustrated with the example of goal-directed movements[J]. Biol Cybern,2003,89(1):22-33.
[60] NEILSON P D,NEILSON M D,O'DWYER N J. Evidence for rapid switching of sensory-motor models[M/C]//PIEK J P.Motor control and human skill: A multidisciplinary perspective. Perth,Australia:Human kinetics,box5076,champaign,IL 61825-5076 USA, 1998: 105-126.
[61] NEILSON P D,NEILSON M D,O’DWYER N J. Adaptive optimal control of human tracking[J]. Adv Psychol,1995,111:97-140.
[62] NEILSON P D,NEILSON M D,O’DWYER N J. Adaptive model theory:Application to disorders of motor control[J]. Adv Psychol,1992,84:495-548.
[63] NEILSON P D,NEILSON M D,O’DWYER N J. Adaptive model theory:Central processing in acquisition of skill[J]. Clin Deval Med,1997 (143):346-370.
[64] NEILSON P D,NEILSON M D,O’DWYER N J. What limits high speed tracking performance?[J]. Hum Mov Sci,1993,12(1):85-109.
[65] NEILSON P D,NEILSON M D. A new view on visuomotor channels:the case of the disappearing dynamics.[J]. Hum Mov Sci,2004,23(3-4):257-283.
[66] NEILSON P D,NEILSON M D. Motor maps and synergies[J]. Hum Mov Sci,2005,24(5-6):774-797.
[67] NEILSON P D,NEILSON M D. On theory of motor synergies[J]. Hum Mov Sci,2010,29(5):655-683.
[68] NEILSON P D,O’DWYER N J,NEILSON M D. Stochastic prediction in pursuit tracking:An experimental test of adaptive model theory[J]. Biol Cybern,1988,58(2):113-122.
[69] NICHOLS T R,STEEVES J D. Resetting of resultant stiffness in ankle fl exor and extensor muscles in the decerebrate cat.[J]. Exp Brain Res,1986,62(2):401-10.
[70] PAN Y,LIU Y,XU B,et al. Hybrid feedback feedforward:An eff i cient design of adaptive neural network control[J]. Neural Netw,2016,76:122-134.
[71] PARK J,LEWIS M M,HUANG X,et al. Effects of olivo-ponto-cerebellar atrophy (OPCA) on fi nger interaction and coordination[J]. Clin Neurophysiol,2013,124(5):991-998.
[72] RESQUÍN F,GONZALEZ-VARGAS J,IBÁÑEZ J,et al. Feedback error learning controller for functional electrical stimulation assistance in a hybrid robotic system for reaching rehabilitation[J]. Eur J Transl Myol,2016,26(3).
[73] SCHMIDT R,LEE T. Motor Control and Learning[M]. 5th ed. Champaign:Human Kinet,1998.
[74] SCHOLZ J P,SCHÖNER G,LATASH M L. Identifying the control structure of multijoint coordination during pistol shooting.[J]. Exp Brain Res,2000,135(3):382-404.
[75] SCHOLZ J P,SCHÖNER G. The uncontrolled manifold concept:identifying control variables for a functional task[J]. Exp Brain Res,1999,126(3):289-306.
[76] SCHÖNER G,THELEN E. Using dynamic field theory to rethink infant habituation[J]. Psychol Rev,2006,113(2):273-299.
[77] SCHONER G. Recent developments and problems in human movement science and their conceptual implications[J]. Ecol Psychol,1995,7(4):291-314.
[78] SEIDLER R D,NOLL D C,THIERS G. Feedforward and feedback processes in motor control[J]. Neuroimage,2004,22(4):1775-1783.
[79] SHADMEHR R,HOLCOMB H H. Neural correlates of motor memory consolidation[J]. Sci,1997,277(5327):821-825.
[80] SHADMEHR R,MUSSA-IVALDI F A. Adaptive representation of dynamics during learning of a motor task[J]. Neurosci,1994,14(5):3208-3224.
[81] SMITH M A,BRANDT J,SHADMEHR R. Motor disorder in Huntington’s disease begins as a dysfunction in error feedback control[J]. Nat,2000,403(6769):544-549.
[82] THOROUGHMAN K A,SHADMEHR R. Electromyographic correlates of learning an internal model of reaching movements[J]. J Neurosci,1999,19(19):8573-8588.
[83] VAN INGEN SCHENAU G J,VAN SOEST A J,GABREËLS F J M,et al. The control of multi-joint movements relies on detailed internal representations[J]. Hum Mov Sci,1995,14(4):511-538.
[84] WAGNER M J,SMITH M A. Shared internal models for feedforward and feedback control[J]. J Neurosci,2008,28(42):10663-10673.
[85] WINTER D A. Biomechanics and Motor Control of Human Gait:Normal,Elderly and Pathological[M].1991.
[86] WOODWORTH R S. The Accuracy of Voluntary Movement..[M]. Columbia University.,1899.
[87] YOUSIF N,DIEDRICHSEN J. Structural learning in feedforward and feedback control[J]. J Neurophysiol,2012,108(9):2373-2382.
Review on Theories and Computational Modeling of Motor Control
WANG Ai-wen,LUO Dong-mei
Human motor control is a comprehensive fi eld which absorbs knowledge gradually from physics,anatomy,physiology,psychology,engineering,neurology,etc. and the researches of human motor control are also from the preliminary human movement describing to the now neuromuscular mechanism. The main problems of human motor control field are redundancy control,abundance control and motor variability which are the abstractions of human movement fl exibilities. Researchers proposed several hypotheses and theories about how to solving these problems including Motor Coding,Hierarchy Control,Equilibrium Point Hypothesis,Reference Configuration Hypothesis,Dynamical Action Theory,Riemann manifold,Uncontrolled Manifold,Tolerance-Noise-Covariation,Adaptive Model Theory etc. Some of these hypotheses are focusing on the information transferring in human motor control proceeding and some of them are focusing on the physical processing. Every hypothesis has its limitation and effect which supplies researchers with different view about the human movement control. In the fi eld,researcher focused only the sole articulation,single muscle but human moves as a whole body so if one articulation or muscle were got problem,it perhaps is caused not by the problem articulation but the other articulations or muscle even by the human recognition. For this reason,we need to evaluate the human movement from the whole prospective. Human motor control hypotheses and theories have the very important affections on the training and rehabilitation that physical therapists proposed new rehabilitation methods and renewed the old methods. According to the training method based on the motor control theory can signi fi cantly decreased the injury risk of athletes. The research in human motor control fi eld can help to understand themechanism of sport injuring and the interaction between equipment and human body,to improve the sport performance.
motor control;human movement;sport biomechanics;rehabilitation theory
G804.6
A
2017-04-14;
2017-07-15
北京体育大学自主科研课题(2016SYS006)。
王爱文,男,在读博士研究生,主要研究方向为运动生物力学、人体运动控制;Tel:(010)62989580,E-mail:ivan.alvan.king@gmail.com;罗冬梅,女,教授,博士,博士研究生导师,主要研究方向为幼儿动作发展,Tel:(010)62989580,E-mail:dmluo02@sina.com。
北京体育大学 运动人体科学学院,北京 100084
Beijing Sport University,Beijing 100084,China.
