基于耳石微结构的南海春季鸢乌贼日龄、生长与种群结构的研究
2017-08-16招春旭陈昭澎何雄波邓玉淑冯颜云榕
招春旭陈昭澎何雄波邓玉淑冯 波,颜云榕,
(1.广东海洋大学水产学院, 湛江 524088; 2.广东海洋大学南海渔业资源监测与评估中心, 湛江 524088; 3.广东省南海深远海渔业管理与捕捞工程技术研究中心, 湛江 524088)
基于耳石微结构的南海春季鸢乌贼日龄、生长与种群结构的研究
招春旭1陈昭澎1何雄波1邓玉淑1冯 波1,2,3颜云榕1,2,3
(1.广东海洋大学水产学院, 湛江 524088; 2.广东海洋大学南海渔业资源监测与评估中心, 湛江 524088; 3.广东省南海深远海渔业管理与捕捞工程技术研究中心, 湛江 524088)
2015年春季在南海开展灯光罩网渔船鸢乌贼(Sthenoteuthis oualaniensis)采样, 样品胴长为10.22—199.01 mm, 体质量为1.3—328.8 g。研究根据鸢乌贼耳石微结构对其日龄、生长和种群结构进行研究, 结果显示:南海存在2个鸢乌贼群体, 中型群体(有发光器, 占59.38%)和微型群体(无发光器, 占40.62%), 无胴长大于500 mm的大型群体; 样品日龄范围为38—126d, 优势日龄组为51—80d, 占其总数的81.33%。分别计算不同海域和群体间5种生长模型的AIC权重值, 发现南海北部海域和南沙附近海域都以指数生长模型最适合描述鸢乌贼的生长关系, 中型群体以指数模型最为适合, 微型群体则以幂函数生长模型最适合; 南海北部海域的鸢乌贼个体生长速率略低于南沙附近海域鸢乌贼个体; 微型群体呈现急剧生长到逐渐缓慢生长的趋势, 中型群体的生长趋势与微型群体正好相反。同类相食现象表明微型群体的生长受到中型群体的影响而被抑制。
鸢乌贼; 耳石; 日龄与生长; 种群结构; 南海
鸢乌贼(Sthenoteuthis oualaniensis)隶属头足纲、枪形目、柔鱼科、鸢乌贼属, 为暖水性较强的大洋性种类[1]。主要分布于印度洋、太平洋的赤道和亚热带等海域, 其中在南海和印度洋西北部海域分布数量较大。近年来, 鸢乌贼成为南海三省区灯光罩网船的主要捕捞对象, 颜云榕等[2]通过对南海三省区的渔业进行抽样调查统计, 2011年鸢乌贼产量约为4×107kg, 为南海外海最大规模种类, 在海洋生态中占据重要的地位。
国外开展了鸢乌贼年龄相关研究, 例如Bizikov[3]基于内壳的研究发现阿拉伯海鸢乌贼生长周期大约在1年, 且雌性比雄性生长得快。Uozumid等[4]通过耳石的研究发现, 水温会影响鸢乌贼的生长。国内学者对鸢乌贼种群结构的研究有, 刘金立等[5]利用RAPD分析表明不同种群之间遗传背景上存在较大差异; 陈新军等[6]从形态学上发现鸢乌贼存在多个产卵群体; 李朋等[7]通过对基因分型提出南海海域存在两个鸢乌贼种群。张宇美等[8]从繁殖生物学方面进行研究, 发现鸢乌贼有两次性腺发育高峰期。鸢乌贼的多种结构对年龄进行的研究中, 耳石的轮纹对印度洋海域鸢乌贼的年龄估算是一种可靠而有效的方法[9]; 江艳娥等[10]根据耳石形态的差异将南海中部海域鸢乌贼分为3个不同群体; 刘必林等[11,12]发现内壳并不适合用作年龄鉴定的材料和角质额能成为年龄鉴定的材料。目前对于南海鸢乌贼年龄与生长研究的报道较少, 特别是对南海鸢乌贼年龄与生长建立适合的生长模型的研究尚未见报道。
本次实验样品采集于2015年春季, 来自于灯光罩网渔船在南海海域的鸢乌贼捕捞生产。实验通过分析采自南海海域鸢乌贼耳石轮纹来估算其生长周期、分析其生长状况, 并比较南海北部海域和南沙附近海域鸢乌贼种群的生长状况, 选择南海鸢乌贼适合的生长模型, 以期为南海鸢乌贼的种群结构分析及其资源可持续开发和管理提供基础数据。
1 材料与方法
1.1 样品采集
采样时间为2015年, 通过灯光罩网渔船在E110°—E117°, N9°—N22°海域进行鸢乌贼采样(图1), 详细记录采样时间和经纬度。共采集鸢乌贼样品943尾, 于–25℃冷冻库中保存, 在实验室进行生物学参数测定和耳石提取。样品信息见表 1。
1.2 生物学参数的测量和耳石的研磨
实验前, 对种类进行鉴定, 微型群体和中型群体根据有无发光器来确定, 测量生物学参数。

图 1 鸢乌贼采样位置Fig.1 Locations of sampling Sthenoteuthis oualaniensis Sampling locations of Sthenoteuthis oualaniensis
头足类的耳石位于头部后方平衡囊(Statocyst)内, 提取耳石后保存到75%浓度的酒精里, 待耳石磨片。耳石的研磨步骤包括: (1)包埋, (2)磨片, (3)抛光, (4)拍照并研读, (5)轮纹计数并测距[14]。
耳石切片在中国科学院水生生物研究所鱼类环境生物学实验室进行拍照与研读。耳石薄片放置在附带有ARTCAM-130MI (Artray Co., Ltd.)拍照装置的BX51光学显微镜下进行观察和拍照存档,采用SPOT.4.0.2软件对耳石的轮纹数和长度进行测量(图 2)。每个耳石样品都由两个人分别读取轮纹数, 两人的日轮读数差值少于10%方可接受, 否则重新再读[13]。
1.3 数据分析
孵化日期的推算参考Uozumi等[4]的头足类一日一轮的结论推测鸢乌贼的孵化日期, 根据日轮数和采样日期推算得出鸢乌贼的孵化日期。
生长模型的建立胴长、体质量和日龄数据分别采用以下的生长模型[14]来拟合不同种群的生长方程:
式中: Yt为t时刻的胴长(mm)或体质量(g); t为年龄(d); Y0为最大胴长或体质量; a、b、G、g和 t0为估算参数。
AIC权重值的计算AIC信息量准则, 也称为赤池信息量准则(Akaike’s information criterion, AIC), 是衡量统计模型拟合优良性的一种标准[15]。
首先对各个参数进行的预测, 计算出预测值p,根据预测值p算出最大似然估计值, 从而得到模型的AIC权重值。
预测值:p=(x)
其中标准差σ为观测值的15%, 最大似然估计值采用excel中的规划求解求得。

表 1 2015年春季鸢乌贼样品信息表Tab.1 The information of the squid in spring 2015 (mean±SE)
AIC值的大小受样本容量的影响较大[16], 对于同一组数据, AIC值越小, 表明该模型拟合度更高。
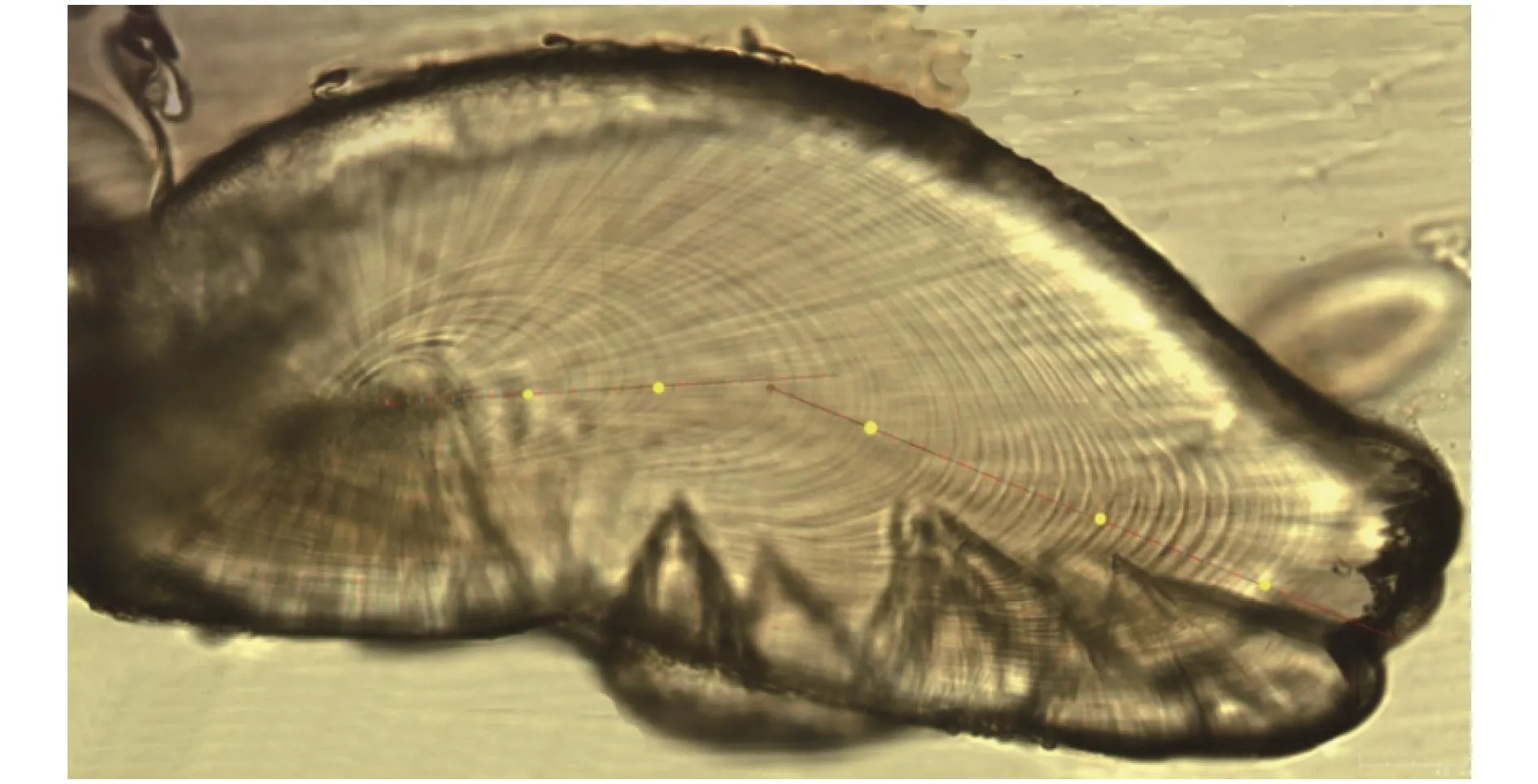
图 2 耳石核心至边缘的拼合图(雌性, 胴长为87 mm, 体质量为23.1 g, 日龄为58d)Fig.2 The mosaic picture of the statolith from core to edge (female, mantle length 87 mm, body mass 23.1 g, and age 58d)
生长速度的估算采用瞬时相对生长率(Instantaneous relative growth rate)和绝对生长率(Absolute growth rate)[17]来分析鸢乌贼的生长, 时间间隔为10d.其计算公式分别为:

式中: S1、S2分别为t1、t2年龄时的胴长(mm)或体质量(g)。
差异显著性分析方法利用Microsoft Excel 2010和SPSS 19软件对数据进行正态分布检验, 方差齐性检验和独立样本t检验, 将鸢乌贼年龄、性别、有无发光器、胴长和体质量等数据进行分析,检验其是否存在差异。
2 结果
2.1 种群结构
2015年春季采集的鸢乌贼样本共943尾, 将该样本的鸢乌贼胴体长度与其性别、海域及所属群体进行独立样本t检验(图 3)。结果表明, 春季样品中, 性别间的胴体长度差异并不显著, P值为0.822>0.05, 地域间和不同群体间的胴体长度有显著性差异, P值均小于0.001。
南海北部海域鸢乌贼胴长为10.22—128.67, 优势胴长组为70.01—130.00 mm, 占总体的82.38%;南沙附近海域鸢乌贼胴长范围为68.66—199.01 mm,优势胴长组为70.01—160.00 mm, 占总体的97.13%。
中型群体鸢乌贼胴长范围为96.45—199.01 mm,优势胴长组为100.01—160.00 mm, 占总体的95.89%;微型群体鸢乌贼胴长范围为10.22—128.67 mm, 优势胴长组为70.01—130.00 mm, 占总体的89.55%。
2.2 年龄结构
本研究共研磨了75尾鸢乌贼的耳石, 有发光器个体44尾, 无发光器个体31尾。南海春季鸢乌贼的日龄范围为38—126d, 日龄最小个体的胴长为118.75 mm, 体质量为50.00 g, 日龄最大个体胴长为199.01 mm, 体质量为328.80 g。平均日龄为64d, 日龄在51—60d这个范围内的个体占的比例最大(38.66%), 其次为61—70d和71—80d日龄组, 分别占29.33%和13.33%。
南海北部海域个体日龄范围为49—88d, 平均日龄为60d, 优势日龄组为51—70d, 占84.61%; 南沙附近海域个体日龄范围为38—126d, 平均日龄为66d, 优势日龄组为51—80d, 占79.51% (图 4)。
中型群体个体日龄范围为38—126d, 平均日龄为68d, 优势日龄组为51—70d, 占90.90%; 微型群体个体日龄范围为42—71d, 平均日龄为58d, 优势日龄组为51—80d, 占68.18% (图 4)。
根据日龄和捕捞日期, 推算孵化日期(图 5), 春季捕获的鸢乌贼个体孵化月份主要集中在同年的1—3月份。南海北部海域个体孵化月份集中在3月份, 占88.46%; 南沙附近海域个体孵化月份集中在1—2月份, 占73.91%。中型群体孵化月份与整体分布相似, 主要集中在1—3月份, 占96.22%; 微型群体孵化月份主要集中在3月份, 占50%。
2.3 生长模型

图 3 鸢乌贼胴长组频数分布图Fig.3 The frequency distribution of squid mantle length
构建线性、指数、幂函数、Gompertz和VonBertalanffy5个生长模型对鸢乌贼生长参数分别进行估计, 并计算AIC权重值(表 2)。对不同海域和群体的鸢乌贼进行5种生长模型的建立(图 6), 在整体、南海北部海域、南沙附近海域和鸢乌贼中型群体的生长方程中, 指数模型的AIC权重值最小, 为最适方程; 鸢乌贼微型群体的生长方程中幂函数模型的AIC权重值最小, 为最适方程。

2.4 生长率
南海北部海域鸢乌贼胴长相对生长率为0.831%/d, 南沙附近海域鸢乌贼胴长相对生长率为0.849%/d。鸢乌贼中型群体的胴长相对生长率为0.703%/d, 微型群体的胴长相对生长率呈逐渐下降趋势(图 7)。
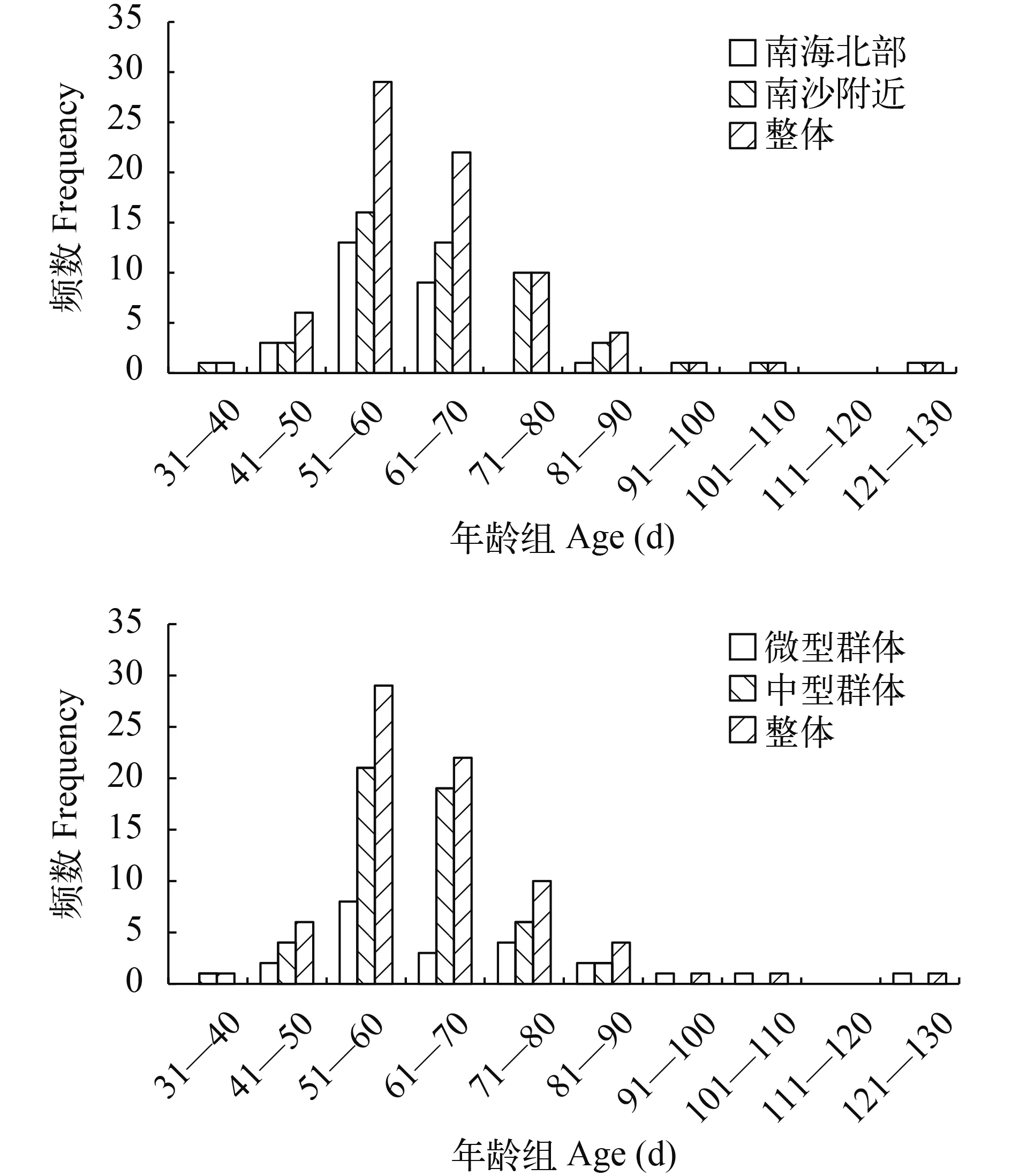
图 4 鸢乌贼年龄组频数分布图Fig.4 The frequency distribution of the squid age
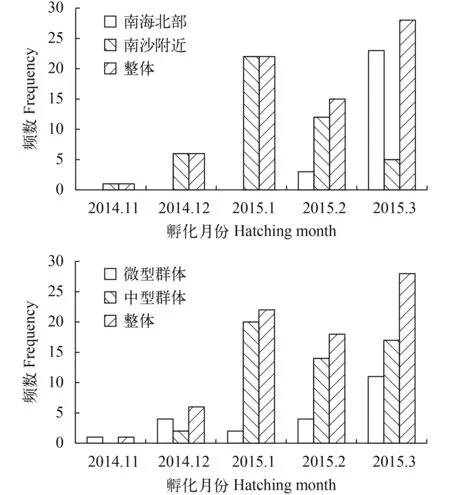
图 5 鸢乌贼孵化月份频数分布图Fig.5 The frequency distribution of squid hatching date
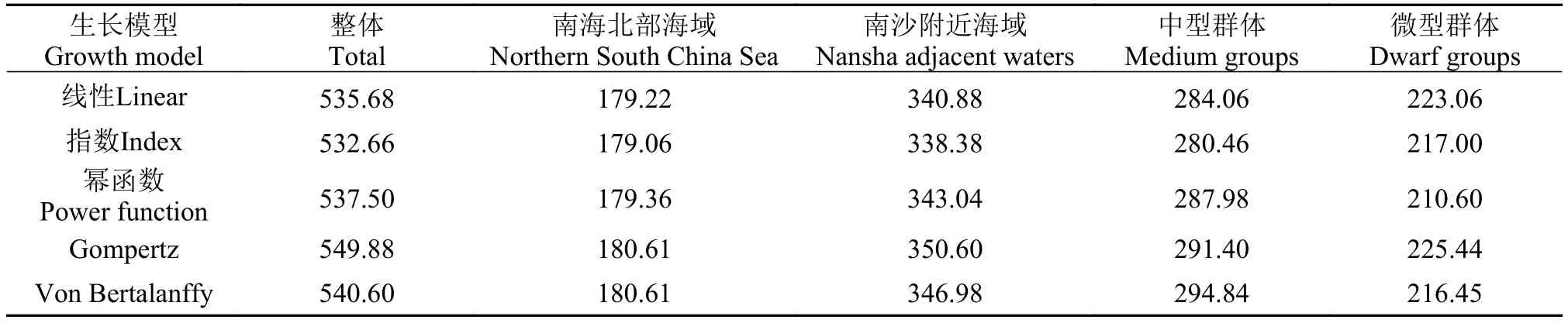
表 2 鸢乌贼生长模型的AIC权重值Tab.2 The AIC weight value of the squid growth model
南沙附近海域的鸢乌贼绝对生长率在生长的各个阶段均略高于南海北部海域的鸢乌贼; 鸢乌贼中型群体的绝对生长率呈不断上升确实, 微型群体绝对生长率则出现急剧下降至平稳状态。
3 讨论
3.1 南海春季鸢乌贼中型与微型种群的结构差异
鸢乌贼可根据胴长划分为大型群体、中型群体和微型群体三大宏观群体, 其中中型群体又可根据内壳叶柄侧轴划分为单内壳叶柄侧轴群体和双内壳叶柄侧轴群体[18]。Yatsu[19]通过胴长对阿拉伯海的鸢乌贼进行了群体划分, 胴长大于500 mm的为大型群体; 中型群体与微型群体通过胴体背部有无发光器进行区分, 无发光器为微型群体, 有发光器为中型群体。在本次实验品中, 鸢乌贼胴长范围在10.22—281.00 mm, 无大型群体。Nesis[20]对鸢乌贼种群的研究发现, 大型种群仅分布在印度洋北部的红海、亚丁湾和阿拉伯海, 中型种群存在于整个鸢乌贼种群分布区域内, 微型种群仅分布在赤道附近, 与本研究结果相吻合。

图6 南海春季鸢乌贼生长模型Fig.6 The growth model of the squid in the South China Sea in spring
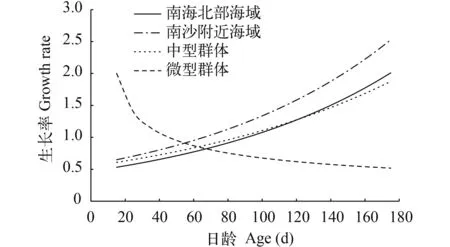
图 7 南海春季鸢乌贼胴长绝对生长率(AGR)Fig.7 The absolute growth rate of squid mantle length in the South China Sea in spring
在实验中发现, 胴长大于128.67 mm的个体, 胴背皮肤下具有明显的黄色椭圆形斑块, 胴长小于96.45 mm的个体, 胴背皮肤与其他部位皮肤无异,并无黄色椭圆形斑块。胴长在96.45—128.67 mm的个体, 存在有发光器和无发光器个体, 部分个体甚至存在不明显的黄色椭圆形斑块, 仔细观察仍可发现胴背位置存在密布的淡色小斑点, 容易误判为微型种群。鸢乌贼自身吐墨染黑胴背部皮肤和样品保存在低温中肌肉呈现软化状态, 造成对这些个体有无发光器的误判。通过鸢乌贼胴背皮肤的卵形光斑来划分鸢乌贼的微型群体和中型群体, 有少量个体处于从微型种群向中型种群过渡的阶段。
目前南海海域尚未发现胴长超过350 mm的个体, 这与谷津明彦[21]研究结果一致, 在赤道附近,小型个体的分布较为集中。刘必林等[22]对印度洋西北海域鸢乌贼的研究发现其胴长范围为142—575 mm, 体质量范围为99—5564 g; 样品包含了微型, 中型和大型个体; 优势胴长组为200—500 mm,占总体的88.8%; 优势体质量组为500—2000 g, 占总体的55.1%。在本次实验品中, 优势胴长组为70.01—160.00 mm, 占总体的93.84%。其中中型群体优势胴长组为100.01—160.00 mm, 微型群体优势胴长组为70.01—130.00 mm。本研究优势胴长组与刘必林等研究差距较大, 主要原因是捕捞方式的不同, 鱿鱼钓的捕捞方式主要捕捉较大个体的头足类, 灯光罩网的渔获分布比较均衡, 且南海尚未发现大型群体分布。
3.2 南海春季鸢乌贼年龄结构
本研究通过估算耳石日龄数和样品的捕捞日期推算出南海春季鸢乌贼的孵化时间集中在同年冬季, 可推断出鸢乌贼为全年产卵, 这可能与其栖息海域的水温常年高于20℃有关[23]。优势日龄组为51—80d, 占81.33%, 可估计鸢乌贼个体生命周期短, 全年分布, 并且1年产卵高峰期可能达到2—3次。由于本次研究在南海北部海域以及南沙附近海域通过灯光罩网渔船只能抓到微型群体和中型群体, 并未捕获大型群体, 只能反映微型群体与中型群体的年龄结构。
Mohamed等[24]对阿拉伯海域鸢乌贼的研究发现, 微型群体生命周期为6个月, 而中型群体和大型群体的生命周期为1年的说法相差甚远, 可能与生长环境等因素不同有关系。刘必林等[22]对印度洋西北海域鸢乌贼的年龄研究发现其日龄范围为88—363d。本次实验样品仅为微型个体和中型个体, 年龄结构差异属正常情况。捕捞方式的不同,捕获的群体也有所差异, 颜云榕等[2]对自动鱿钓和灯光罩网作业的调查发现, 灯光罩网的网囊网目较小, 选择性低, 而鱿钓的作业方式对个体较大的鸢乌贼有较高的选择性。
3.3 南海春季鸢乌贼的生长模型和生长率
外界生物因素和非生物因素时刻影响着头足类的生长发育[25], 因此在不同海域, 不同种群耳石受到不同环境影响, 而造成耳石轮纹的生长模型也有所不同。为了更准确地描述某一生长阶段的生长, 通常采用多种生长模型相结合的方法。
在线性、指数、幂函数、Gompertz和Von Bertalanffy五种生长模型中。整体、不同海域和中型群体的最适生长模型都为指数生长模型, 该部分鸢乌贼个体呈现随年龄增大, 生长速率逐渐增大的趋势。微型群体个体以幂函数生长模型最为适合, 微型群体随年龄增大, 胴体达到一定范围, 生长速率趋于平稳。生长模型的选择符合鸢乌贼个体的实际生长情况。
南海北部海域的鸢乌贼个体绝对生长速率略低于南沙附近海域的鸢乌贼个体。春季南海北部海域海表水温基本低于28℃, 南沙附近海域接近于29℃[26]。海表水温在25—28℃适合于鸢乌贼个体的集群和栖息[24], 南海北部海域的海表水温比南沙附近海域更适于群体的集群。在相同饵料情况下,生物量较大的群体的个体生长较为缓慢。因此南海北部海域的鸢乌贼个体生长速率较慢于南沙附近海域的鸢乌贼个体。
微型群体呈现急剧生长到逐渐缓慢生长的趋势, 中型群体则与微型群体正好相反, 中型群体低龄时生长较为缓慢, 绝对生长速率逐渐增大。当日龄达62d时, 微型群体和中型群体的绝对生长率相似, 此时微型群体胴长约为90 mm, 中型群体达到120 mm。该年龄段的中型群体个体较大于微型群体, 鸢乌贼个体存在同类相食现象[8], 鸢乌贼大个体会以小个体为食, 导致中型群体逐渐增大而微型群体的生长被抑制。
[1]Zhang X, Qi Z Y, Dong Z Z, et al.Decapodes (Cephalopodes) in China coast [J].Oceanologia et Limnologia Sinica, 1960, 3(3): 188—204 [张玺, 齐钟彦, 董正之, 等.中国沿岸的十腕目(头足纲).海洋与湖沼, 1960, 3(3): 188—204]
[2]Yan Y R, Feng B, Lu H S, et al.Fishery biology of purpleback flying squid Sthenoteuthis oualaniensis in northern sea areas around Nansha islands in su mmer [J].Oceanologia et Limnologia Sinica, 2012, 6(43): 1177—1186 [颜云榕, 冯波, 卢伙胜, 等.南沙群岛北部海域鸢乌贼(Sthenoteuthis oualaniensis)夏季渔业生物学研究.海洋与湖沼, 2012, 6(43): 1177—1186]
[3]Bizikov V A.Growth of Sthenoteuthis oualaniensis, using a new method based on gladius microstructure [C].ICES Marine Science Symposia.1995(1999): 445—458
[4]Uozumi Y, Ohara H.Age and growth of Nototodarus sloanlii (cephalopods: Oegopsida) based on daily increment counts in statolith [J].Nippon Suisan Gakkaishi, 1993, 9(59): 1469—1477
[5]Liu J L, Chen X J, Xu H Q.Population genetic structure of Sthenoteuthis oualaniensis in the Northwestern Indian Ocean by RAPD analysis [J].South China Fisheries Science, 2008, 2(4): 43—49 [刘金立, 陈新军, 许华强.印度洋西北部海域鸢乌贼种群遗传结构的RAPD分析.南方水产, 2008, 2(4): 43—49]
[6]Chen X J, Liu J L.Morphological analysis on population structure of Sthenoteuthis oualaniensis in the northwestern Indian Ocean [J].Journal of Shanghai Fisheries University, 2007, 2(16): 174—179 [陈新军, 刘金立.利用形态学方法分析印度洋西北部海域鸢乌贼种群结构.上海水产大学学报, 2007, 2(16): 174—179]
[7]Li P.Population genetic structure of Sthenoteuthis oualaniensis in South China Sea [D].Thesis for Master of Science.Shanghai Ocean University, Shanghai.2014 [李朋.南海鸢乌贼的种群遗传结构.硕士学位论文.上海海洋大学, 上海.2014]
[8]Zhang Y M, Yan Y R, Lu H S, et al.Study on feeding and reproduction biology of purple flying squid, Sthenoteuthis oualaniensis in the Western South China Sea [J].Journal of Guangdong Ocean University, 2013, 33(3): 56—64 [张宇美, 颜云榕, 卢伙胜.西沙群岛海域鸢乌贼摄食与繁殖生物学初步研究.广东海洋大学学报, 2013, 33(3): 56—64]
[9]Ma J, Liu B L, Chen X J, et al.Advances on using statolith to study age and growth of cephalopod [J].Marine Fisheries, 2009, 31(3): 316—324 [马金, 刘必林, 陈新军,等.利用耳石鉴定头足类年龄与生长研究进展.海洋渔业, 2009, 31(3): 316—324]
[10]Jiang Y E, Chen Z Z, Lin Z J, et al.Statolith morphology of purpleback flying squid Sthenoeuthis oualaniensis in the central South China Sea [J].South China Fisheries Science, 2014, 10(4): 85—90 [江艳娥, 陈作志, 林昭进,等.南海中部海域鸢乌贼耳石形态特征分析.南方水产科学, 2014, 10(4): 85—90]
[11]Liu B L, Chen X J, Li J H.Advances on age and growth of cephalopods by using gladius, stylets and cuttlebone [J].Marine Fisheries, 2015, 1(37): 68—76 [刘必林, 陈新军, 李建华.内壳在头足类年龄与生长研究中的应用进展.海洋渔业, 2015, 1(37): 68—76]
[12]Liu B L, Chen X J.Beak length analysis of the purpleback flying squid Sthenoeuthis oualaniensis in northwest Indian Ocean [J].Progress in Fishery Sciences, 2010, 1(31): 8—14 [刘必林, 陈新军.印度洋西北海域鸢乌贼角质颚长度分析.渔业科学进展, 2010, 1(31): 8—14]
[13]Liu B L.Studying age and growth of purple back flying squid (Sthenoeuthis oualaniensis) in northwest Indian Ocean based on statolith microstructure [D].Thesis for Master of Science.Shanghai Ocean University, Shanghai.2006 [刘必林.利用耳石微结构研究印度洋西北海域鸢乌贼的年龄和生长.硕士学位论文.上海海洋大学, 上海.2006]
[14]Bigelow K.Age and growth of three species of squid paralarvae from Hawaiian waters as determined by statolith microstructures [D].Thesis for Master of Science.USA University of Hawaii.1991
[15]Akaike H.Likelihood of a model and information criteria[J].Journal of Econometrics, 1981, 16(1): 3—14
[16]Hou G, Liu J D, Feng B, et al.Applying multi-model inference to estimate growth parameters of greater lizard fish Saurida tumbil in Beibu Gulf, South China Sea [J].Chinese Journal of Applied Ecology, 2014, 25(3): 843—849 [侯刚, 刘金殿, 冯波, 等.应用多模型推论估算北部湾多齿蛇鲻的生长参数.应用生态学报, 2014, 25(3): 843—849]
[17]Argüelles J, Rodhouse P G, Villegas P, et al.Age, growth and population structure of the jumbo flying squid Dosidicus gigas in Peruvian waters [J].Fisheries Research, 2001, 54(1): 51—61
[18]Zhang P, Yan L, Yang B Z, et al.Population structure of purpleback flying squid (Sthenoteuthis oualaniensis) in Nansha area in spring [J].South China Fisheries Science, 2015, 5(11): 11—19 [张鹏, 晏磊, 杨炳忠, 等.春季南沙海域鸢乌贼种群结构特征的研究.南方水产科学, 2015, 5(11): 11—19]
[19]Yatsu A.The biology of Sthenoteuthis oualaniensis and exploitation of the new squid resources [J].Bull of Far-Sea Fishery, 1991, 101: 6—9
[20]Nesis K N.Cephalopods of the World: Squids, Cuttlefishes, Octopuses, and Allies [M].New Jersey: TFH Publications.1987, 351—352
[21][谷津明彦.インド洋におけるトビイカの生物学並びに新資源としての可能性.遠洋, 1997, 101: 6—9]
[22]Liu B L, Chen X J, Zhong J S.Age, growth and popuation structure of squid Sthenoteuthis oualaniensis in northwest Indian Ocean by statolith microstructure [J].Journal of Dalian Fisheries University, 2009, 3(24): 206—212 [刘必林, 陈新军, 钟俊生.采用耳石研究印度洋西北海域鸢乌贼的年龄、生长和种群结构.大连水产学院学报, 2009, 3(24): 206—212]
[23]Basir S.Biological Feature of an Oceanic Squid, Sthenoteuthis oualaniensis in the South China Sea, Area Ⅲ: Western philippines [A].Proceedings of the SEAFDEC Seminar on Fishery Resources in the South China Sea, Area III: Western Philippines [C].Southeast Asian Fisheries Development Center.2000, 135—147
[24]Mohamed K S, Sasikumar G, Koya K P S, et al.Know the Master of the Arabian Sea-purple-back Flying Squid Sthenoteuthis oualaniensis [M].NAIP Booklet, Central Marine Fisheries Research Institute.2011, 5—10
[25]Siriraksophon S, Nakamura Y, Pradit S, et al.Ecological aspects of oceanic squid, Sthenoteuthis oualaniensis in the South China Sea, area Ⅲ: Western Philippines [A].Proceedings of the SEAFDEC Seminar on Fishery Resourcesin the South China Sea, Area III: Western Philippines [C].Southeast Asian Fisheries Development Center.1999, 101—117
[26]Lin L R, Zhao H.Analysis on the relations between sea surface temperature and phytoplankton Chlorophyll-a in the South China Sea [J].Journal of Marine Sciences, 2012, 4(30): 46—54 [林丽茹, 赵辉.南海海域浮游植物叶绿素与海表温度季节变化特征分析.海洋学研究, 2012, 4(30): 46—54]
AGE, GROWTH AND POPULATION STRUCTURE OF PURPLE BACK FLYING SQUID, STHENOTEUTHIS OUALANIENSIS IN THE SOUTH CHINA SEA IN SPRING BASED ON STATOLITH MICROSTRUCTURE
ZHAO Chun-Xu1, CHEN Zhao-Peng1, HE Xiong-Bo1, DENG Yu-Shu1, FENG Bo1,2,3and YAN Yun-Rong1,2,3
(1.College of Fisheries, Guangdong Ocean University, Zhanjiang 524088, China; 2.Center of South China Sea Fisheries Resources Monitoring and Assessment, Guangdong Ocean University, Zhanjiang 524088, China; 3.Guangdong Provincial Engineering and Technology Research Center of Far Sea Fisheries Management and Fishing of South China Sea, Zhanjiang 524088, China)
The current study investigated the purple back flying squid (Sthenoteuthis oualaniensis) collected by light attracting falling-net fishing vessels in the spring of 2015 in the South China Sea.The mantle lengths of S.oualaniensis were ranged from 10.22 to 199.01 mm while the body mass were ranged from 1.3 to 328.8 g.The Age, growth and population structures of S.oualaniensis were studied based on the microstructure of statolith.The result showed that there were two groups of S.oualaniensis in the South China Sea: the medium-sized (with photophore on the dorsal mantle) and dwarf groups (without photophore on the dorsal mantle) accounting for 59.38% and 40.62%, respectively, and that there was no large group whose mantle lengths were more than 500 mm.The ages of the sample were ranged from 38 to 126d among which the dominant age was from 51 to 80d, accounting for 81.33%.According to the AIC values of these five growth models in different waters and groups, the exponential growth model appropriately reflect the growth of S.oualaniensis in northern South China Sea and Nansha adjacent waters; the exponential growth model suitably explain the middle-size group and the power function growth model was best for dwarf groups.Moreover, the individual growth rate of squid in the northern South China Sea was slightly slower than that in Nansha adjacent waters.The growth rate of the dwarf groups first increased rapidly and then gradually decreased while the medium-sized groups were totally on the contrary.
Sthenoteuthis oualaniensis; Statolith; Age and growth; Population structure; The South China Sea
S917.4
A
1000-3207(2017)04-0884-07
10.7541/2017.110
2016-08-31;
2016-12-26
国家自然科学基金(41376158); 广东海洋大学创新强校工程项目(GDOU2013050305, GDOU2013050212); 大学生创新创业训练计划项目(CXXL2014002)资助 [Supported by the National Natural Science Foundation of China (41376158), Project of Enhancing School With Innovation of Guangdong Ocean University (GDOU2013050305, GDOU2013050212), Students’ Platform for Innovation and Entrepreneurship Training Program (CXXL2014002)]
招春旭(1991—), 男, 广东湛江人; 硕士研究生; 主要研究方向为渔业资源和捕捞学。E-mail: 294276856@qq.com
颜云榕(1975—), 男, 博士, 教授; 主要研究方向为南海渔业资源保护与开发。E-mail: yanyr@gdou.edu.cn
