中华沙鳅和宽体沙鳅形态差异及其物种有效性分析
2017-08-16徐丹丹王志坚王永明彭敏锐谢碧文何学福李
徐丹丹王志坚王永明彭敏锐谢碧文何学福李 斌
(1.内江师范学院生命科学学院, 内江 641100; 2.西南大学淡水鱼类资源与生殖发育教育部重点实验室, 重庆 400715)
中华沙鳅和宽体沙鳅形态差异及其物种有效性分析
徐丹丹1王志坚2王永明1彭敏锐1谢碧文1何学福2李 斌1
(1.内江师范学院生命科学学院, 内江 641100; 2.西南大学淡水鱼类资源与生殖发育教育部重点实验室, 重庆 400715)
通过主成分分析法并结合物种的生物学特征对长江上游支流沱江资中段69尾宽体沙鳅Sinibotia reevesae和长江干流宜宾段、南溪段及沱江资中段141尾中华沙鳅Sinibotia superciliaris的10个常规可量性状与20个框架性状进行了比较研究。结果显示: 主成分分析并不能区分宽体沙鳅和中华沙鳅之间的形态差异;依据二者在尾柄高、吻须长、D1-3(胸鳍基部起点至腹鳍基部起点之间的距离)、D7-9(臀鳍基部后末端至尾鳍基部腹部起点之间的距离)和D9-10(鳍基部腹部起点至尾鳍基部背部起点之间的距离) 等形态特征的显著性差异(P<0.05), 并结合其繁殖生物学特征, 认为中华沙鳅和宽体沙鳅应为2个有效种。建议在使用框架性状度量评价物种有效性时需选择恰当的形态指标, 数据分析应结合物种的生物学特征。
宽体沙鳅; 中华沙鳅; 框架性状度量方法; 形态差异
中华沙鳅(Sinibotia superciliaris)和宽体沙鳅(Sinibotia reevesae)隶属于鲤形目(Cypriniformes),鳅科(Cobitidae), 沙鳅亚科(Botiinae), 中华沙鳅属(Sinibotia), 是我国长江上游特有鱼类, 栖息于流水底层环境中, 具有重要的科学研究价值和经济价值[1]。作为长江上游鳅科鱼类典型代表的中华沙鳅和宽体沙鳅外部形态特征极为相似, 以传统分类方法对二者进行区分尚无统一标准[2,3]。随着长江上游鱼类资源保护和开发需求的不断加强, 对鳅科鱼类的准确鉴定、原种资源的优化选择的需求也越来越高。
长期以来, 表型性状快速检测遗传变异是一种古老而又简便易行的方法, 且广泛应用于动植物分类学中。现行的鱼类分类学形态度量方法主要基于Hubs和Lagler[4]提出的形态测量标准, 该方法主要从二维空间角度上衡量鱼类的外部形态特征[5]。近年来, 框架性状度量评价方法已经成功应用于鱼类物种鉴定中[6]。为准确区分长江上游不同地理单元的中华沙鳅和宽体沙鳅的形态差异, 本研究应用框架性状度量评价方法并结合物种的生物学特征对二者形态差异和物种有效性进行了探究, 以期为其野生资源的种质资源鉴定与评价及合理保护提供理论依据。
1 材料与方法
1.1 实验材料
所检视和测量的标本均来自长江上游鱼类资源保护与利用四川省重点实验室淡水鱼类标本馆。实验材料为福尔马林浸泡标本, 共计210尾, 在进行种类鉴定时, 以吻长是否大于眼前缘至头后端长作为二者区别的主要依据[2]。依据上述分类方法鉴定后获得宽体沙鳅69尾, 中华沙鳅141尾。中华沙鳅和宽体沙鳅不同地理种群的标本编号、采集地等基本信息见表 1。
1.2 形态参数测定
应用游标卡尺测定了常规形态性状和框架性状的30个参数, 共得到6930个原始数据, 其中: 头长(HL)、吻长(SnL)、尾柄长(CPL)、眼径(ED)、体高(BD)、尾柄高(CPD)、眼后头长(EHL)、眼间距(IW)、颌须长(MBL)、吻须长(RBL)10个参数为传统形态性状参数(图 1a)。框架数据测量参照于红霞等[7]的方法, 设置了10个坐标点, 分别是1.胸鳍基部起点、2.吻前端、3.腹鳍基部起点、4.枕骨后末端、5.臀鳍基部起点、6.背鳍基部起点、7.臀鳍基部后末端、8.背鳍基部后末端、9.尾鳍基部腹部起点、10.尾鳍基部背部起点。它们之间的距离用D1-2、D1-3……D9-10表示, 共产生 20个参数(图 1b)。
1.3 数据处理与分析
应用SPSS 16.0软件对可量性状进行主成分分析: 分析前所有数据进行自然对数转换(ln)以除去异速生长(Allometries)的影响, 再依据其方差-协方差矩阵(Variance-covariancematrix)提取主成分。同时应用Statistic 6.0软件对中华沙鳅和宽体沙鳅可量性状参数进行了方差分析(Breakdown and One-way ANOVA, P<0.05)。常规数据统计在Excel 2003软件中完成。
2 结果
2.1 方差分析
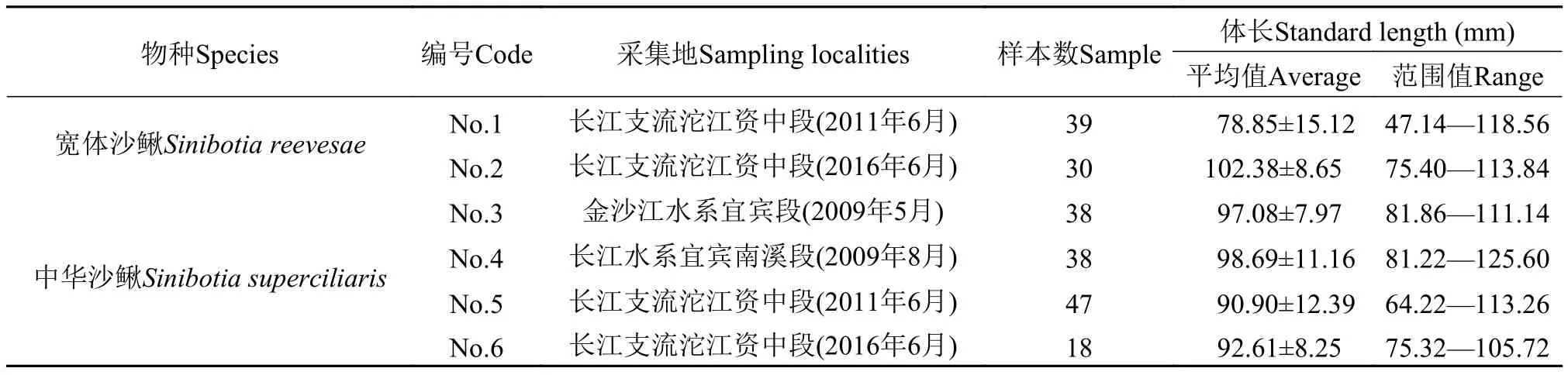
表 1 宽体沙鳅和中华沙鳅的采集地、数目与体长Tab.1 Sampling sities, numbers and standard length of Sinibotia reevesae and S.superciliaris
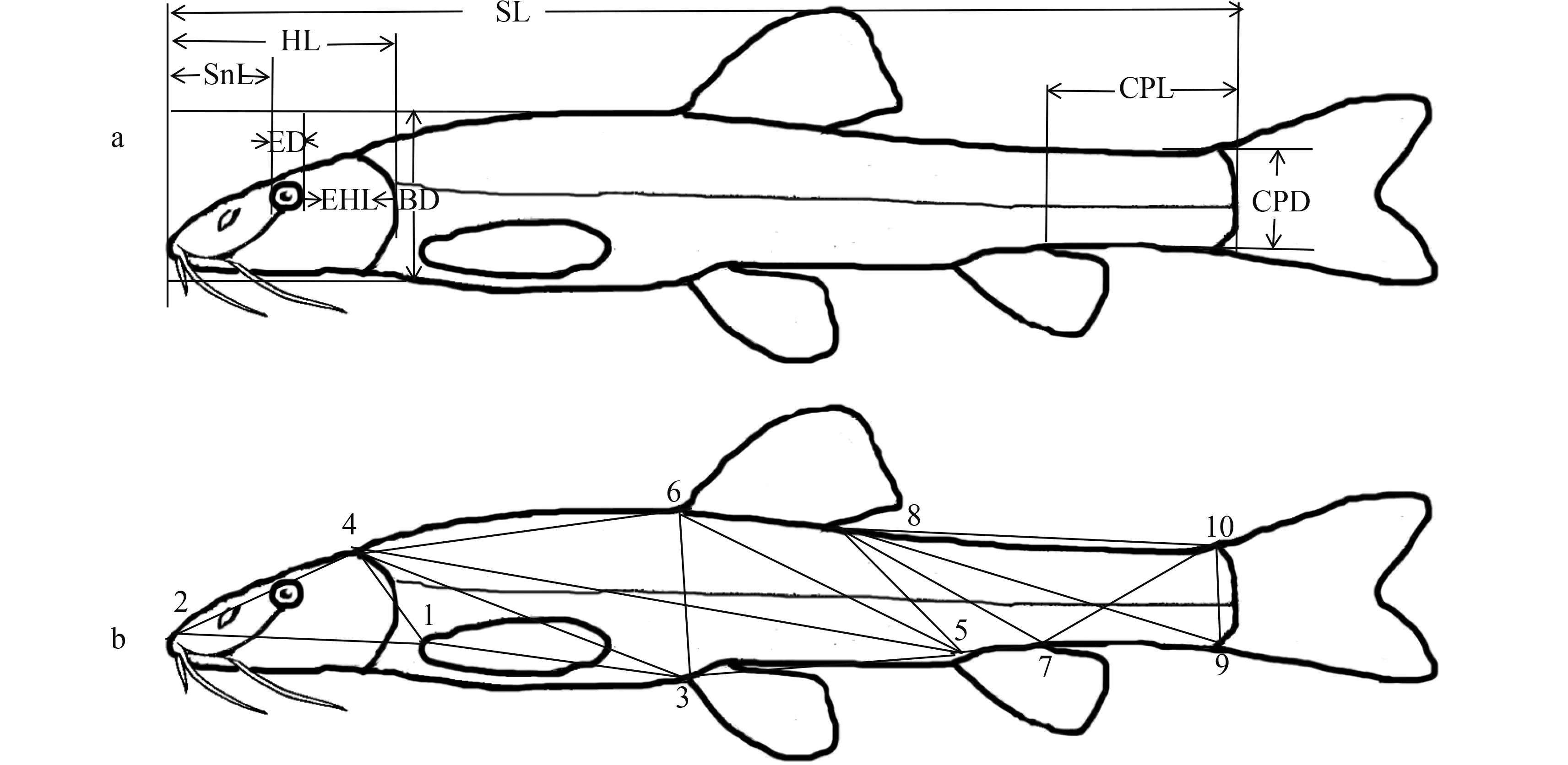
图 1 样本可量性状测量示意图Fig.1 Morphometric measurements of the samples
表 2显示了宽体沙鳅和中华沙鳅30组参数的比例性状。方差分析(ANOVA)显示, 2种鱼在吻须长、尾柄高、D1-3、D7-9和D9-10等形态参数方面差异显著(P<0.05), 其余25项差异均不显著(P>0.05)(表 3)。
2.2 主成分分析
对宽体沙鳅和中华沙鳅的30组性状参数进行相关性分析, KMO (Kaiser-Meyer-Olkin measure)统计量(KMO=0.668)和Bartlett球型检验结果(P<0.01)显示, 变量之间相关性强, 适合进行因子分析降维。
采用主成分分析得到前11个主成分的因子载荷(表 4), 其方差的累积贡献率达到63.74%, 前3个主成分贡献率达到29.06%。在PC1(第一主成分)轴上具有较大因子载荷的分别是D3-5、D5-6和D4-5;在PC2(第二主成分)轴上具有较大因子载荷的是颌须长、体高和D7-8; 在PC3(第三主成分)轴上具有较大因子载荷的是眼径和D2-4。为了深入探讨上述性状的分类学意义, 对PC1轴上具有较大因子载荷的3个性状进行了三维分布图分析(图 2), 结果显示宽体沙鳅和中华沙鳅并未形成两个独立的群体,同样的情况也存在于PC2和PC3轴上具有较大因子载荷的几个性状之中。
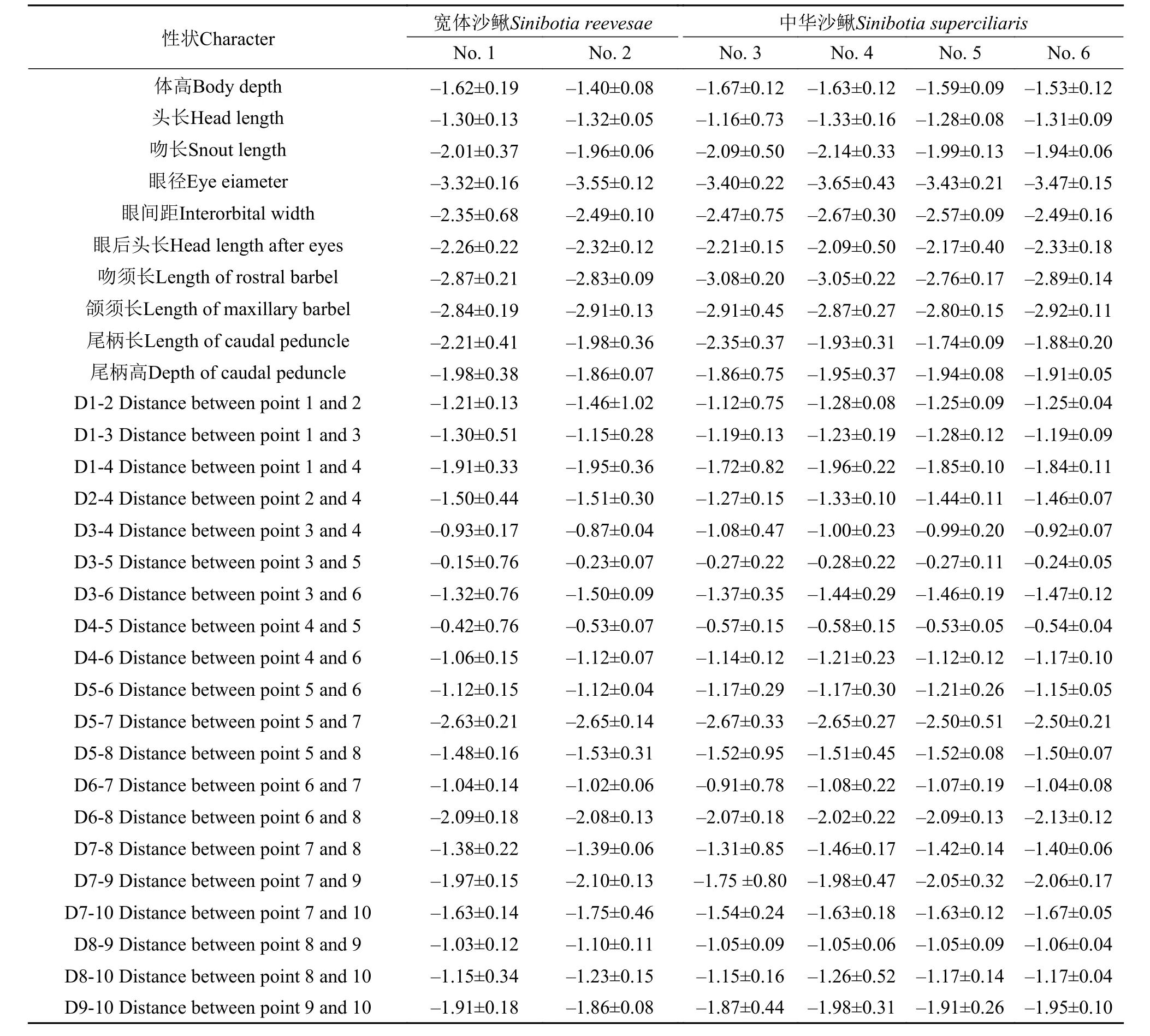
表 2 宽体沙鳅与中华沙鳅比例性状数据(平均值±标准差)Tab.2 Proportional characters for Sinibotia reevesae and S.superciliaris (Means±SD)
图 3和图 4分别是PC1与PC2和PC2与PC3的二维散布图。从图 3中可以看出, 无论是在PC1轴还是PC2轴上, 宽体沙鳅与中华沙鳅标本的重叠度非常大, 二者均不能互相分离; 同样图 4显示的结果与图 3一致。由此可见, 分布于我国长江上游的宽体沙鳅和中华沙鳅外部形态特征差异不显著。
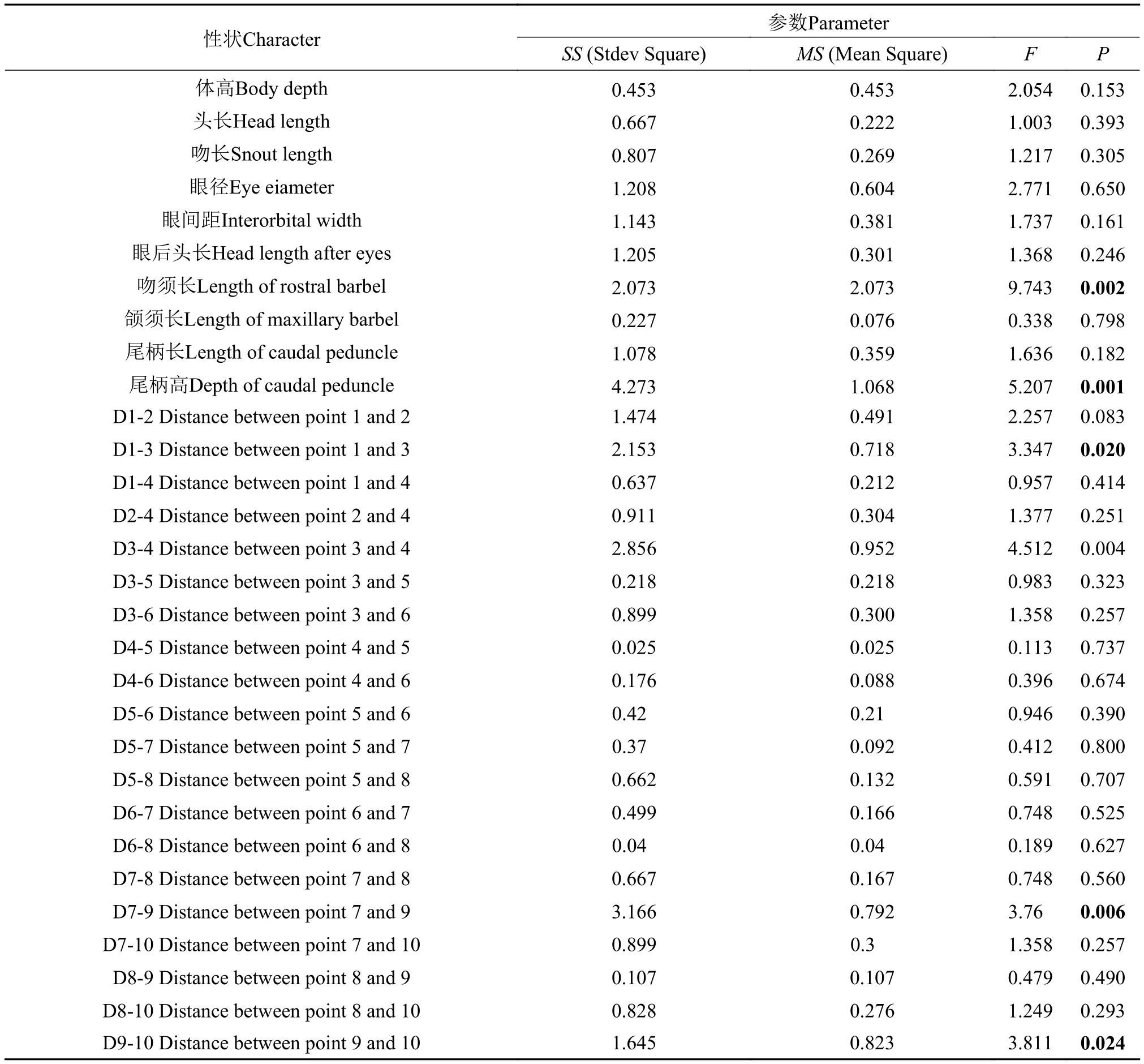
表 3 中华沙鳅与宽体沙鳅性状参数的方差分析Tab.3 The variance analysis of characteristic parameters for Sinibotia reevesae and S.superciliaris
2.3 聚类分析
由聚类分析结果(图 5)可知, 长江上游不同地理单元(No.1、No.2、No.3、No.4、No.5和No.6)的宽体沙鳅和中华沙鳅未形成预想中的两个独立的分支, 表明聚类分析并不能依据二者外部形态特征参数值差异将其区分开来。
3 讨论
3.1 中华沙鳅和宽体沙鳅的分类阶元问题
沙鳅亚科隶属鲤形目, 含7属, 约60种[8]。依据眼下刺是否分叉及颊部有无鳞片, 沙鳅亚科分为薄鳅属、沙鳅属和副沙鳅属[9]。传统形态学研究认为中华沙鳅和宽体沙鳅隶属于沙鳅属[2,3]。随着分子生物学的发展, 毛云涛等[10]通过线粒体基因对沙鳅亚科鱼类系统发育进行的研究, 认为中华沙鳅独立于沙鳅属之外, 单独聚为一支即中华沙鳅属, 隶属于沙鳅亚科; 黄燕[4]通过对长江上游特有鱼类DNA条形码研究认为中华沙鳅和宽体沙鳅均隶属于中华沙鳅属, 这些研究表明中华沙鳅和宽体沙鳅的分类地位存在较大争议。
3.2 两种沙鳅鱼类的有效性
通过对宽体沙鳅和中华沙鳅30个可量性状的主成分分析发现, 二者未形成2个明显的独立单元(图 3、图 4); 同样聚类分析也不能将二者很好的区分开来(图 5)。故本研究中框架性状度量评价的多元分析方法并未达到区分中华沙鳅和宽体沙鳅形态特征差异的目的, 但是从传统的形态学和生殖生态特征可以明确的对它们加以区别。

表 4 主成分分析中的因子符合矩阵及主成分对总变异的贡献率Tab.4 Loadings of components matrix and contributions on principal components to total variance of principal component analysis
形态差异根据方差分析显示中华沙鳅与宽体沙鳅在尾柄高、吻须长和D7-9等局部形态参数差异极为显著(P<0.01)(表 3); 同时笔者在研究中发现二者尾鳍末端形状差异十分明显, 中华沙鳅尾鳍末端呈“Λ”形状, 而宽体沙鳅尾鳍末端呈“Ω”形状。上述形态差异可作为区分中华沙鳅和宽体沙鳅的重要传统形态特征, 可以明显区别两种沙鳅。
生殖生态特征鱼类在长期自然演化过程中, 为适应各种类型的水体环境和生活方式, 进化出多样化的生殖生态类型, 这也是区分物种的一个重要依据[11,12]。尽管中华沙鳅和宽体沙鳅在食性类型、栖息类型和产卵方式(一次性产卵, 无黏性)等方面较为相似(表 5), 但二者的繁殖生物学特征差异十分明显, 具体如下: ①中华沙鳅的绝对繁殖力(平均值为6780粒)2倍于宽体沙鳅(平均值为3230粒), 因此本次研究中宽体沙鳅样品量明显少于中华沙鳅; ②同等温度条件下中华沙鳅受精卵吸水膨胀后的卵膜外直径2倍于宽体沙鳅, 但孵化时间明显多于后者(表 5); ③卵的类型与孵化条件有区别, 宽体沙鳅为半漂流性卵, 而中华沙鳅为漂流性卵。孵化时, 尤其是人工繁殖采用的孵化方式明显不同。依据以上特征, 中华沙鳅和宽体沙鳅应为二独立物种。
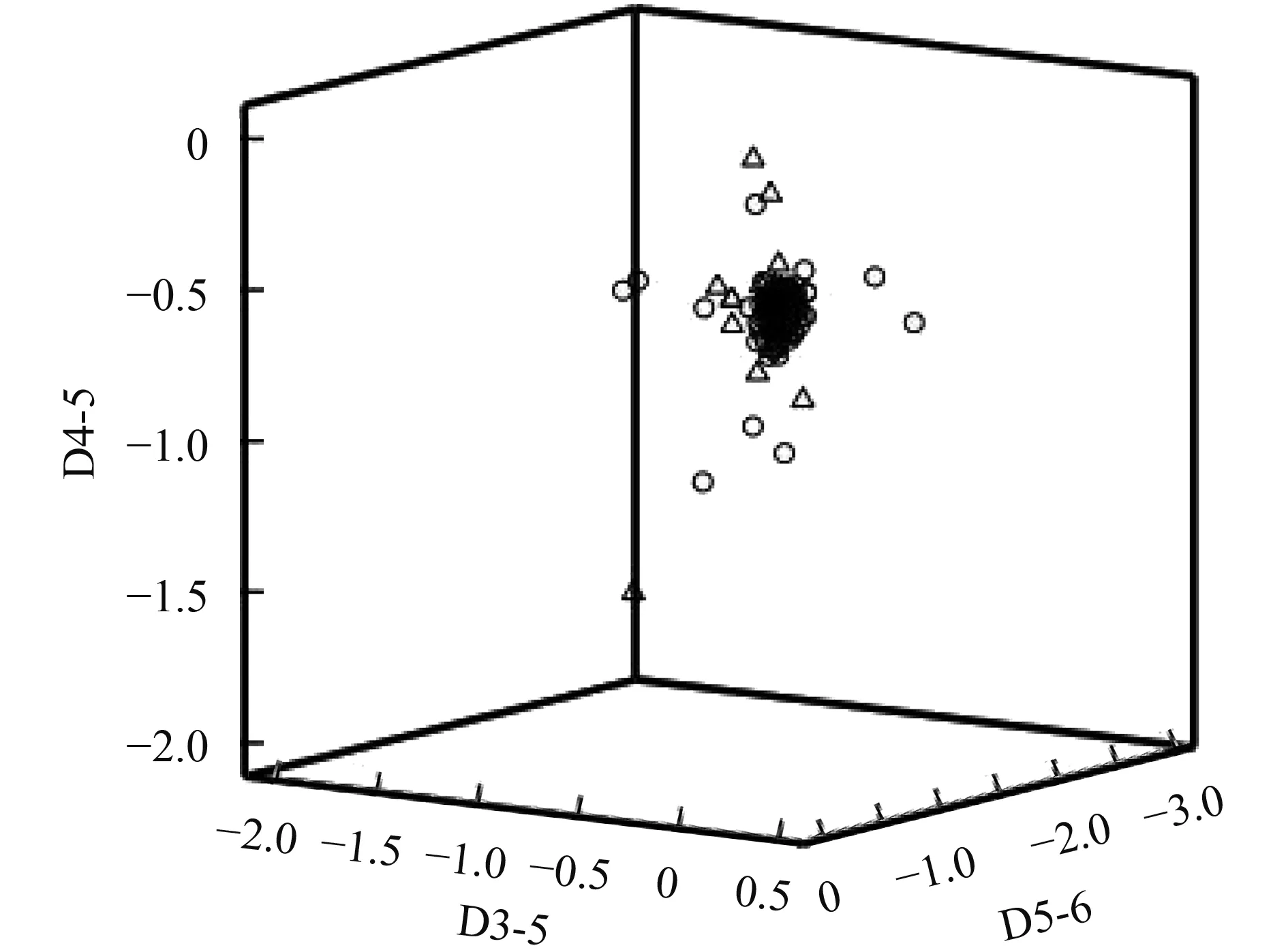
图 2 宽体沙鳅和中华沙鳅的D3-5、D4-5和D5-6的三维分布图Fig.2 Three dimensional distribution of D3-5, D4-5 and D5-6 for Sinibotia reevesae and S.superciliaris
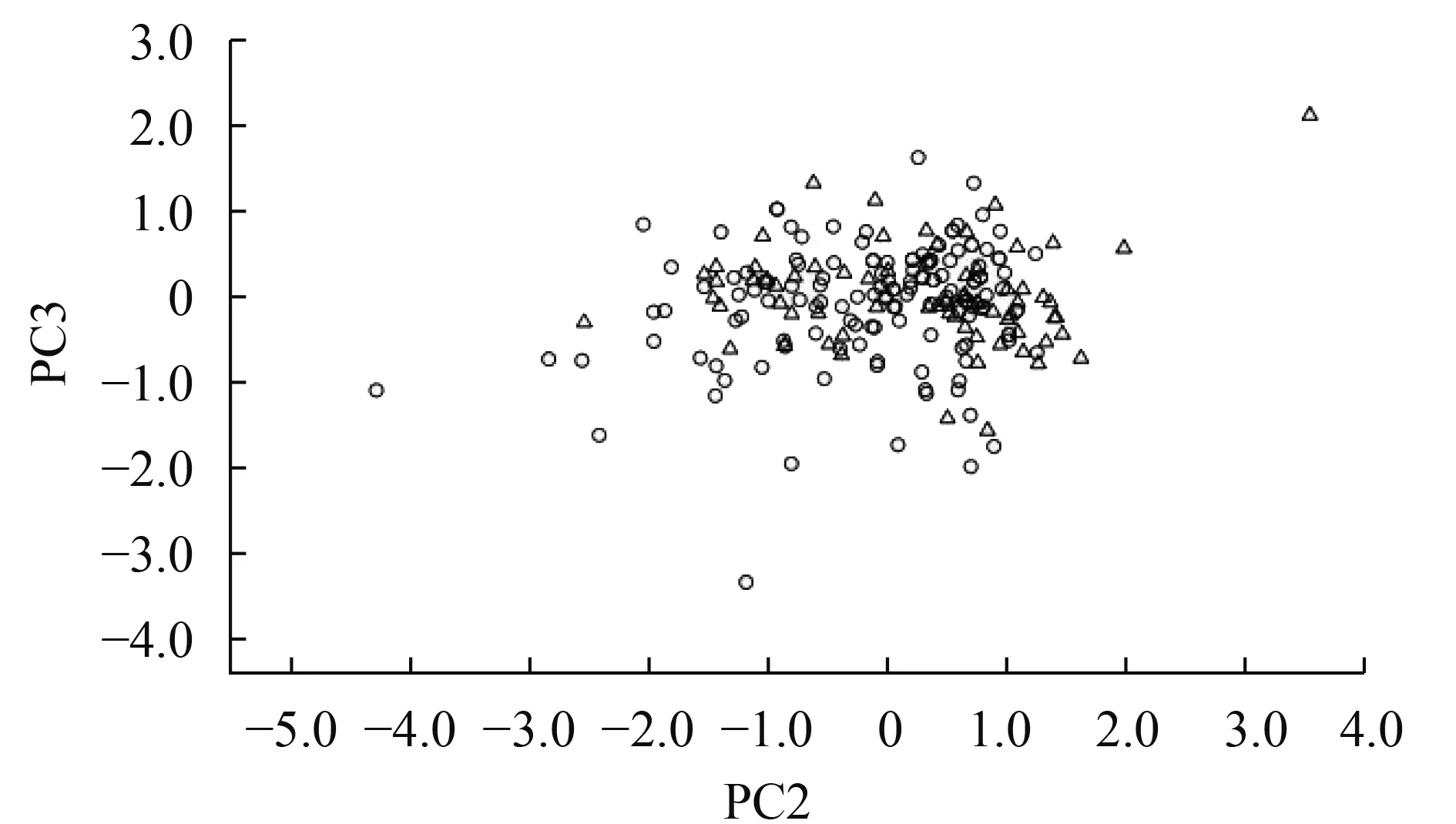
图 3 宽体沙鳅和中华沙鳅性状第一、二主成分散布图(Δ代表宽体沙鳅, Ο代表中华沙鳅)Fig.3 Scatterplot on the first two principle components extracted from characters of Sinibotia reevesae and S.superciliaris
3.3 基于框架性状度量评价效果分析
合理恰当分析方法的选择对鱼类种类区分至关重要。框架性状参数评价方法是通过选取鱼体外部形态特征坐标点的方式将鱼体分成若干个网格单元进行多维空间度量, 结合多变量分析方法进行量化分析, 其分类结果较为准确可靠[19]。目前采用传统可量性状和框架性状相结合的多变量形态度量学方法已被广泛应用于裂腹鱼Schizothoracinae[20]、鲂属Megalorama[21]、中华纹胸鲦Glyptothorax sinense[22]、团头鲂Megalobrama amblycephala[23]等鱼类的种间分类中, 并获得了较好的研究结果。
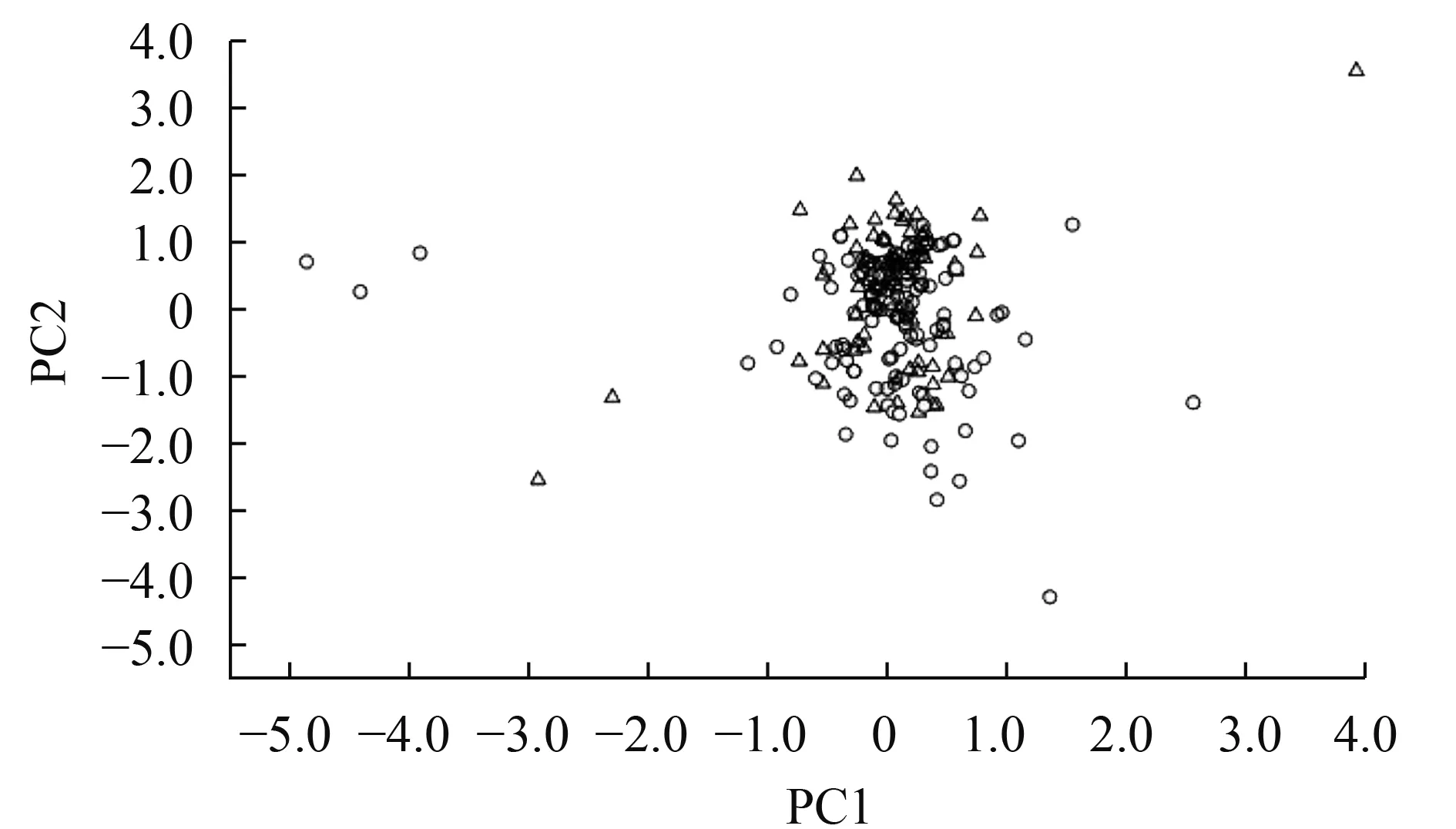
图 4 宽体沙鳅和中华沙鳅性状第二、三主成分散布图(Δ代表宽体沙鳅, Ο代表中华沙鳅)Fig.4 Scatterplot on the second and third principle components extracted from characters of Sinibotia reevesae and S.superciliaris

图 5 六个样本的聚类分析图Fig.5 Cluster dendrogram of six samples

表 5 宽体沙鳅和中华沙鳅生殖生态特征比较Tab.5 The ecology and reproductive characteristics of Sinibotia reevesae and S.superciliaris
宽体沙鳅和中华沙鳅是长江上游特有鱼类, 也是产区重要的经济鱼类, 市场价值高, 且供不应求。近年来, 过度捕捞、水体环境污染和水利工程建设等因素导致中华沙鳅和宽体沙鳅的资源量急剧下降[15], 资源量锐减可能会诱发其杂交现象增加;同时中华沙鳅和宽体沙鳅两种沙鳅在种间杂交具有较好的亲和力(数据显示二者在杂交后代受精卵和孵化率分别达到46.8%和72.8%)[24], 是其杂交现象发生的生物学基础。本研究应用了框架性状度量评价方法却未能清楚区分中华沙鳅和宽体沙鳅区之间的形态差异, 其原因是否与二者杂交产生的中间性状干扰有关?仍需进一步研究。
综上所述, 尽管框架性状度量评价方法已经得到广泛应用, 但本研究显示该方法并不适用于区分宽体沙鳅和中华沙鳅之间的形态特征。因此使用框架性状度量参数分析时需要注意: 针对不同的研究对象选取恰当框架结构测量和正确对数据的进行处理分析是十分重要的环节; 数据解释应结合研究对象的生物学特征和及其生态环境特征。
致谢:
数据测量过程中得到了内江师范学院生命科学学院学生向霞、邱德林等同学的大力帮助, 在此一并致谢。
[1]Li Q, Yao M Y, Zhou B, et al.Technique of artificial reproduction of Botia superciliaris Günther [J].Journal of Freshwater Fisheries of China, 2011, 41(5): 92—95 [李强, 姚明予, 周波, 等.中华沙鳅人工繁殖技术初探.淡水渔业, 2011, 41(5): 92—95]
[2]Ding R H.The Fishes of Sichuan [M].Chengdu: Sichuan Science and Technology Press.1994, 96—104 [丁瑞华.四川鱼类志.成都: 四川科学技术出版社.1994, 96—104]
[3]Chu X L, Chen Y R.The Fishes of Yunnan 2 [M].Beijing: Science Press.1990, 68—70 [褚新洛, 陈银瑞.云南鱼类志下卷.北京: 科学出版社.1990, 68—70]
[4]Huang Y.DNA barcoding of the endemic fishes from the upper Yangtze River basin [D].Master Dissertation Southwest University, Chongqing.2014 [黄燕.长江上游特有鱼类DNA条形码研究, 西南大学硕士生研究论文,重庆.2014]
[5]Hubbs C L, Lagler K F.Fishes of the Great Lakes Region [M].Ann Arbor: University of Michigan Press.1964, 276—279
[6]Zhang E, Xie Z G, Xie C X.Morphological variation between Sinibrama macrops and S.wui, with notes on their validities [J].Acta Hydrobiologica Sinica, 2004, 28(5): 511—517 [张鄂, 谢仲桂, 谢从新.大眼华鳊和伍氏华鳊的形态差异及其物种有效性.水生生物学报, 2004, 28(5): 511—517]
[7]Yu H X, Tang W Q, Li S F.Ontogenetic changes in meristic measurements of silver carp and bighead carp [J].Zoological Research, 2010, 31(2): 169—176
[8]Tang Q Y, Yu D, Liu H Z.Leptobotia zebra should be revised as Sinibotia zebra (Cypriniformes: Botiidae) [J].Zoological Research, 2008, 29(1): 1—9 [唐琼英, 俞丹,刘焕章.斑纹薄鳅(Leptobotia zebra) 应该为斑纹沙鳅(Sinibotia zebra).动物学研究, 2008, 29(1): 1—9]
[9]Zhu S Q.Synopsis of Fresh Water Fishes of China [M].Nanjing: Jiangsu Science and Technology Publishing House.1995, 1—549 [朱松泉.中国淡水鱼类检索.南京: 江苏科学技术出版社.1995, 1—549]
[10]Mao Y T, Gan X N, Wang X Z.DNA barcodes and molecular phylogeny of botiinae fishes based on the mitochondrial COI gene [J].Acta Hydrobiologica Sinica, 2014, 38(4): 737—744 [毛云涛, 甘小妮, 王绪祯.基于线粒体COI基因的沙鳅亚科鱼类DNA条形码及其分子系统发育研究.水生生物学报, 2014, 38(4): 737—744]
[11]Yin M C.Fishery Ecology [M].Beijing: Chinese Agricultural Press.1993, 105—132 [殷名称.鱼类生态学.北京:中国农业出版社.1993, 105—132]
[12]Xie C X.Ichthyology [M].Beijing: Chinese Agricultural Press.2010, 331—360 [谢从新.鱼类学.北京: 中国农业出版社.2010, 331—360]
[13]Wang F, Yue X J, Xie B W, et al.Anatomical and histological structure of digestive system of Botia reevesae [J].Sichuan Journal of Zoology, 2011, 30(4): 569—572 [王芳, 岳兴建, 谢碧文, 等.宽体沙鳅消化系统的结构.四川动物, 2011, 30(4): 569—572]
[14]Li B, Tao M, Xu D D, et al.Diet of Sinibotia superciliaris by stable isotope signature from the upper reaches of the Yangtze River [J].Acta Hydrobiologica Sinica, 2016, 40(5): 978—984 [李斌, 陶敏, 徐丹丹, 等.长江上游中华沙鳅食性特征的C、N同位素分析.水生生物学报, 2016, 40(5): 978—984]
[15]Yue X J, Wang F, Xie B W, et al.Embryonic development of Botia reevesae in Tuojiang River [J].Sichuan Journal of Zoology, 2011, 30(3): 390—397 [岳兴建, 王芳, 谢碧文, 等.沱江流域宽体沙鳅的胚胎发育.四川动物, 2011, 30(3): 390—397]
[16]Yang M S, Ding X.Study on the reproductive biology of Botia superciliaris [J].Journal of Hydroecology, 2010, 3(2): 38—41 [杨明生, 丁夏.中华沙鳅的繁殖生物学研究.水生态杂志, 2010, 3(2): 38—41]
[17]Huang Y, Wang F, Yue X J, et al.Studies on the individual fecundity of Botia revesae in the Tuojiang River, a tributary river of the Yangtze River [J].Sichuan Journal of Zoology, 2011, 30(6): 916—920 [黄燕, 王芳, 岳兴建,等.沱江宽体沙鳅个体繁殖力的研究.四川动物, 2011, 30(6): 916—920]
[18]He B, Chen X Y, Wen T, et al.Embryonic development of Botia superciliaris [J].Southwest China Journal of Agricultural Sciences, 2014, 27(3): 1332—1337 [何斌, 陈先均, 温涛, 等.中华沙鳅的胚胎发育.西南农业学报, 2014, 27(3): 1332—1337]
[19]Cavalcanti M J, Monteiro L E, Lopes P R D.Landmarkbased morphometric analysis in selected species of Serranid Fishes (Perciformes; Teleostei) [J].Zoological Studies, 1999, 38(3): 287—294
[20]Li Z L, Chen Y X, Hu S Y, et al.Multivariate analysis on the Morphological differentiation of Kozlov’s Schizothoracin (Schizothorax kozlovi) and David’s Schizothoracin (S.davidi) [J].Chinese Journal of Zoology, 2015, 50(4): 547—554 [李忠利, 陈永祥, 胡思玉, 等.四川裂腹鱼和重口裂腹鱼形态差异的多元分析.动物学杂志, 2015, 50(4): 547—554]
[21]Cai M J, Zhang M Y, Zeng Q L.A study on morphometrics of the Genus Megalobrama [J].Acta Hydrobiologica Sinica, 2001, 25(6): 631—635 [蔡鸣俊, 张敏莹, 曾青兰.鲂属鱼类形态度量学研究.水生生物学报, 2001, 25(6): 631—635]
[22]Xie Z G, Zhang E, He S P.Study on species validation for Glyptothorax sinense (Regan) and G.fukiensis (Rendahl) with the method of Morphometrics [J].Journal of Huazhong Agricultural University, 2001, 20(2): 169—172 [谢仲桂, 张鹗, 何舜平.应用形态度量学方法对中华纹胸和福建纹胸物种有效性的研究.华中农业大学学报, 2001, 20(2): 169—172]
[23]Li S F, Cai W Q, Zhou B Y.Morphological and biochemical genetic variations among populations of blunt snout bream (Megalobrama amblycephala) [J].Journal of Fisheries of China, 1991, 15(3): 264—210 [李思发, 蔡完其,周碧云.团头鲂种群间的形态差异和生化遗传差异.水产学报, 1991, 15(3): 264—210]
[24]Wang Y M, Xie B W, Yue X J, et al.Embryonic development of hybrid of Sinibotia superciliaris (♀) and Botia reevesae (♂) [J].Sichuan Journal of Zoology, 2014, 33(1): 90—98 [王永明, 谢碧文, 岳兴建, 等.中华沙鳅(♀)与宽体沙鳅(♂)杂交的胚胎发育.四川动物, 2014, 33(1): 90—98]
MORPHOLOGICAL VARIATION BETWEEN SINIBOTIA SUPERCILIARIS AND SINIBOTIA REEVESAE WITH NOTES ON THEIR VALIDITIES
XU Dan-Dan1, WANG Zhi-Jian2, WANG Yong-Ming1, PENG Min-Rui1, XIE Bi-Wen1, HE Xue-Fu2and LI Bin1
(1.Key Laboratory of Sichuan Province for Fishes Conservation and Utilization in the Upper Reaches of the Yangtze River, School of Life Sciences, Neijiang Normal University, Neijiang 641100, China; 2.Key Laboratory of Freshwater Fish Reproduction and Development (Ministry of Education), Southwest University, Chongqing 400715, China)
Sinibotia reevesae and Sinibotia superciliaris are endemic fish with similar morphology that live only in the upper reaches of the Yangtze River.The current study assessed 10 traditional morphological parameters and 20 truss network features of 210 specimens from these two species using traditional taxonomy method and biological characteristics.69 specimens of S.reevesae were collected from Zizhong section of the Tuojiang River, a tributary river of the Yangtze River and 141 specimens of S.superciliaris were collected from Zizhong section of the Tuojiang River, Yibin and Nanxi section of the Yangtze River.PCA analysis could not distinguish morphological difference of these two species, although there were significant differences in other aspects, such as the height of caudal peduncle, the length of rostral barbel, the distance of D1-3 (from the pectoral fin base to the origin of pelvic fin base), D7-9 (from the end of anal fin base to the ventral origin of caudal fin base) and D9-10 (from the ventral origin of caudal fin base to the dorsal origin of caudal fin base).The S.reevesae and S.superciliaris should be considered as separate species based on characteristics of the reproductive biology.Appropriate evaluation criteria should be chosen to evaluate the species validities while use truss network features, and the data analysis should combine with the biological characteristics of species.
Sinibotia reevesae; Sinibotia superciliaris; Framework character method; Morphological variations
Q958.8
A
1000-3207(2017)04-0827-08
10.7541/2017.103
2016-07-12;
2016-11-14
内江师范学院校级项目(15JC12); 四川省教育厅重点项目(14ZA0251); 四川省科技厅支撑项目(2011NZ0075)资助 [Supported by the Project of Neijiang Normal University (15JC12); the Key Fund Project of Sichuan Province (14ZA0251); Science and Technology Support Project of Sichuan Province (2011NZ0075)]
徐丹丹(1986—), 女, 新疆昌吉人; 硕士; 研究方向为鱼类遗传与进化。E-mail: xudandan5201@163.com
李斌, E-mail: libin2004xinyang@126.com
