不同浓度外源乙酸钠对耐乙酸大肠埃希菌DA19代谢和关键酶酶活的影响
2017-01-03张艳军张萍华董曼茗
张艳军, 张萍华, 周 灿, 董曼茗
(浙江师范大学 化学与生命科学学院, 浙江 金华 321004)
不同浓度外源乙酸钠对耐乙酸大肠埃希菌DA19代谢和关键酶酶活的影响
张艳军, 张萍华, 周 灿, 董曼茗
(浙江师范大学 化学与生命科学学院, 浙江 金华 321004)
为研究外源乙酸钠对大肠埃希菌DA19生长代谢的影响,将该菌株在氮源限制基本培养基及添加不同浓度乙酸钠的氮源限制基本培养基中连续培养,测定稳态时生长代谢参数和胞内关键酶酶活。与MN培养基相比,葡萄糖比消耗速率和延胡索酸比生成速率随外源乙酸钠质量浓度增加而逐渐下降,丙酮酸比生成速率则随外源乙酸钠质量浓度增加而明显增加,而乙酸比生成速率则明显降低(除9 g/L乙酸钠外)。磷酸果糖激酶、异柠檬酸脱氢酶、异柠檬酸裂解酶、苹果酸脱氢酶、磷酸烯醇式丙酮酸羧化酶和乙酸激酶酶活随外源乙酸钠质量浓度增加而呈先下降后上升的趋势,而6-磷酸葡萄糖脱氢酶则随着外源乙酸钠质量浓度增加而逐渐降低。为了应对外源乙酸钠压力,大肠埃希菌DA19的生长代谢和中心代谢途径酶活都发生了明显改变。
大肠埃希菌;乙酸;酶活
大肠埃希菌是最常用的表达宿主之一,被广泛用于各种重组蛋白和小分子物质的生产[1]。但其在培养过程中易产生乙酸,不但降低了碳源利用效率,而且积累到一定浓度时会抑制菌体生长,降低外源基因表达效率[2]。例如,大肠埃希菌BL21(DE3)生产栖热菌属麦芽糖淀粉酶(ThMA)时,在培养基中添加50 mmol/L乙酸可以显著抑制菌体生长,同时使ThMA表达水平降低90%[3]。通过优化大肠埃希菌培养工艺和菌株,可以在一定程度上减少乙酸产生。但这些方法大多具有一定局限性,而且无法完全避免乙酸生成,尤其是在大规模工业化生产中[4],即使限制性补加葡萄糖,也会因局部葡萄糖浓度过高或溶氧限制而造成乙酸积累。朱才庆等[5]以提高大肠埃希菌乙酸耐受性为目标,采用60Co诱变和乙酸为选择性压力连续培养相结合的方法,成功选育了耐乙酸突变株DA19,该菌株在耐乙酸能力增强的同时还减少了乙酸生成。在以该突变株作为表达宿主时,培养基中乙酸质量浓度达到12 g/L时仍可表达外源产物人表皮生长因子[6],说明利用耐乙酸突变株可以改善外源蛋白表达时乙酸抑制情况。虽然传统的诱变育种方法并不是改造菌种、提高菌株乙酸耐受力的最佳选择,但是对所得到耐乙酸突变株的研究有助于理解乙酸耐受机理,为设计新的耐乙酸途径提供信息[7]。本研究对耐乙酸大肠埃希菌DA19在添加不同浓度乙酸钠的氮源限制基本培养基中进行了连续培养,比较该菌株在不同外源乙酸压力下生长代谢和关键酶酶活差异,从而解析外源乙酸钠对大肠埃希菌DA19生长代谢的影响,为下一步研究该菌株对乙酸压力的应答机理提供参考。
1 材料与方法
1.1 材料
1.1.1 菌种 大肠埃希菌DA19是由大肠埃希菌DH5α筛选得到的耐乙酸突变株[5]。
1.1.2 培养基(g/L) LB培养基:蛋白胨10,酵母抽提物5,NaCl 10,pH 7.2;M培养基:Na2HPO4·12H2O 15.12,KH2PO43,NaCl 0.5,MgSO4·7H2O 0.5,CaCl20.011,葡萄糖8,氯化铵1,1%(质量分数)维生素B10.2 mL,微量元素混合液[8]0.2 mL,pH 7.0;MN培养基(氮源限制的M培养基):葡萄糖8,氯化铵0.3,其他成分与M培养基相同;MNA培养基(添加乙酸钠的MN培养基):其中乙酸钠质量浓度分别为3.6、4.8、7.0、9.0 g/L。所有培养基均使用去离子水配制。
1.1.3 仪器与试剂 5 L发酵罐(国强生化FMG-5L);SBA-40C谷氨酸葡萄糖分析仪(山东省科学院生物研究所);液相色谱(安捷伦1260);酶活检测试剂盒(苏州科铭生物技术有限公司)。
1.2 方法
1.2.1 种子培养 将1 mL冷冻保存的大肠埃希菌DA19菌种接入装有30 mL LB培养基的250 mL锥形瓶,37 ℃、200 r/min培养12 h为一级种子。转接3 mL一级种子至装有100 mL M培养基的500 mL锥形瓶中,37 ℃、200 r/min培养10 h为二级种子。
1.2.2 发酵罐中连续培养 在5 L发酵罐中进行连续培养,初始装入2.3 L MN或MNA培养基,接入200 mL二级种子,搅拌转速500 r/min,通气量4 L/min,37 ℃下分批培养至静止期,通过控制培养基流加速率使得菌体生长速率在0.16/h左右。每隔4 h取样测定菌体浓度、葡萄糖浓度和pH,连续3次不变即认为到达稳态。离心收集上清液用于产物检测,所得菌体用于胞内关键酶酶活测定。
1.2.3 分析方法 菌体浓度采用浊度法,将培养液适当稀释后测OD600,按标准曲线计算菌体干重。采用SBA-40C谷氨酸葡萄糖分析仪测定葡萄糖浓度。采用Berthelot法[9]比色法测定铵离子浓度。采用液相色谱检测样品中丙酮酸、乙酸和延胡索酸浓度,其色谱条件:COSMOSIL C18-MS-II柱(4.6 mm×250 mm,5 μm);以0.01 mol/L磷酸溶液为流动相A,以甲醇为流动相B,两者比例为95∶5,流速为0.5 mL/min;进样量20 μL,检测波长为215 nm,柱温为25 ℃。采用酶活检测试剂盒测定胞内关键酶酶活,包括:磷酸果糖激酶(PFK)、6-磷酸葡萄糖脱氢酶(G6PDH)、异柠檬酸脱氢酶(ICDH)、异柠檬酸裂解酶(ICL)、苹果酸脱氢酶(MDH)、磷酸烯醇式丙酮酸羧化酶(PEPC)和乙酸激酶(ACK)。其中1个单位的酶活定义为在测定条件下每分钟形成或消耗1 nmol NADH(或NADPH)所需要的酶量。本研究采用比酶活(U/mg DCW),即单位干菌体的酶活,所列数据为3次测定平均值。
2 结果与分析
2.1 大肠埃希菌DA19在MN和MNA培养基中连续培养
大肠埃希菌DA19在氮源限制的MN和MNA培养基中进行连续培养,达到稳态时测定各参数,其结果见表1。所有连续培养达到稳态时铵离子浓度均为0,因此未在表1中列出。
与MN培养基相比,大肠埃希菌DA19的葡萄糖比消耗速率(QGlc)随着外源乙酸钠质量浓度增加而呈逐渐下降趋势,在9 g/L时降低了约10%。同时,添加外源乙酸钠使菌体关于葡萄糖的得率系数(YX/G)增加了约10%。
与MN培养基相比,丙酮酸关于葡萄糖的得率系数(YPyr/G)和比生成速率(QPyr)随着外源乙酸钠质量浓度增加而明显增加,并在7 g/L乙酸钠时达到最高,分别为MN培养基中的214%和202%。此后两者在9 g/L乙酸钠时无明显增加。与丙酮酸不同,添加3.6、4.8和7.0 g/L乙酸钠使乙酸关于葡萄糖的得率系数(YAC/G)比MN培养中分别降低了约34%、28%和29%,而乙酸比生成速率(QAC)比MN培养中分别降低了约37%、29%和30%,但外源乙酸钠增加至9.0 g/L时,YAC/G和QAC则分别为MN时的186%和172%。
与MN培养基相比,延胡索酸关于葡萄糖的得率系数(YFUM/G)和比生成速率(QFUM)随着外源乙酸钠质量浓度增加而明显降低,在7.0 g/L乙酸钠时分别为MN培养基中的36%和38%,此后在9.0 g/L乙酸钠时两者略有降低。
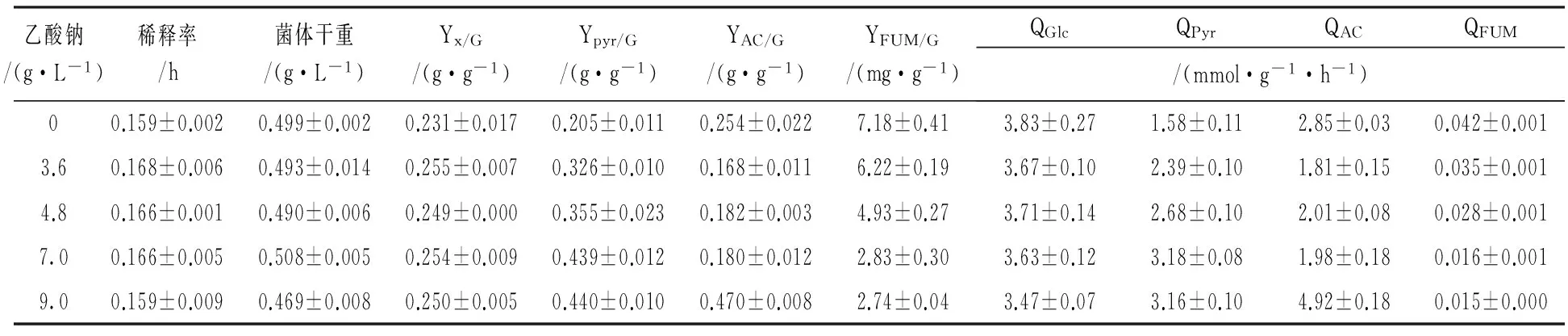
表1 大肠埃希菌 DA19在MN和MNA培养基中连续培养时稳态参数Table 1 Process parameter of DA19 continuously cultured in the MN and MNA medium
2.2 不同浓度乙酸钠对大肠埃希菌DA19胞内关键酶酶活的影响
在MN基本培养基中添加乙酸钠使大肠埃希菌DA19生长代谢发生了明显改变,因此对其胞内参与糖代谢相关途径的关键酶酶活进行了测定。
2.2.1 糖酵解途径(EMP) 磷酸果糖激酶(PFK)催化果糖-6-磷酸 (fructose 6-phosphate)生成1,6-二磷酸果糖,它是糖酵解作用的限速酶。如图1所示,在MN培养基中添加乙酸钠使得磷酸果糖激酶酶活逐步降低,并在4.8 g/L时达到最低,为未添加乙酸钠时酶活的82%。当添加乙酸钠质量浓度进一步增加时,磷酸果糖激酶酶活反而逐步升高,并在9.0 g/L时达到未添加乙酸钠时酶活的94%。因此磷酸果糖激酶酶活随添加乙酸钠质量浓度的增加呈先下降后上升的趋势。

图1 不同外源乙酸钠质量浓度对磷酸 果糖激酶酶活的影响Fig.1 Effect of the different concentrations of exogenous sodium acetate on the activity of phosphofructokinase
2.2.2 磷酸戊糖途径(PPP) 磷酸戊糖途径提供菌体合成所必需的前体物质和还原力(NADPH),对于菌体合成和抵抗氧化压力具有非常重要的作用。6-磷酸葡萄糖脱氢酶(G6PDH)催化的6-磷酸葡萄糖脱氢反应为不可逆反应,在生理条件下属于限速反应,是磷酸戊糖途径的重要调控点[10]。如图2所示,在MN培养基中添加乙酸钠后,6-磷酸葡萄糖脱氢酶酶活变化与磷酸果糖激酶明显不同,它随添加乙酸钠质量浓度的增加而逐渐降低,在7.0 g/L和9.0 g/L时酶活仅为未添加乙酸钠时酶活的65%左右。

图2 不同外源乙酸钠质量浓度对6-磷酸葡萄 糖脱氢酶酶活的影响Fig.2 Effect of the different concentrations of exogenous sodium acetate on the activity of glucose-6-phosphate dehydrogenase
2.2.3 三羧酸(TCA)循环和乙醛酸支路(GS) 进入三羧酸循环的碳流在形成异柠檬酸后有两个流向:继续三羧酸循环或者进入乙醛酸支路。异柠檬酸脱氢酶是三羧酸循环与乙醛酸支路分支后的第一个酶,也是三羧酸循环的一个限速酶,而异柠檬酸裂解酶是乙醛酸支路的一个重要酶。苹果酸脱氢酶是三羧酸循环中一个酶,催化苹果酸脱氢生成草酰乙酸。如图3所示,在MN培养基中添加乙酸钠后,异柠檬酸脱氢酶和苹果酸脱氢酶酶活变化趋势与磷酸果糖激酶酶活基本一致,即随添加乙酸钠浓度增加呈先下降后上升的趋势,均在4.8 g/L时酶活最低,分别为未添加乙酸钠时酶活的79%和61%,而在9.0 g/L乙酸钠时酶活分别为未添加乙酸钠时酶活的117%和89%。大肠埃希菌DA19在MN培养基中连续培养时,其胞内乙醛酸支路的异柠檬酸裂解酶酶活仅为异柠檬酸脱氢酶的1/20左右,远低于以乙酸为碳源时的活性[11]。添加乙酸钠后,异柠檬酸裂解酶酶活随着乙酸钠质量浓度增加也呈先下降后上升的趋势,但其酶活仍远低于相同条件下异柠檬酸脱氢酶酶活。

图3 不同外源乙酸钠质量浓度对异柠檬酸脱氢酶、 异柠檬酸裂解酶和苹果酸脱氢酶酶活的影响Fig.3 Effect of the different concentrations of exogenous sodium acetate on the activity of isocitrate dehydrogenase, isocitrate lyase and malate dehydrogenase
2.2.4 磷酸烯醇式丙酮酸羧化酶(PEPC)回补途径 三羧酸循环不仅能为菌体生长提供能量,而且还是生物合成所必需中间体的重要来源。当TCA途径这些中间产物因参与氨基酸合成而减少时,需要通过回补途径进行补充,才能保证TCA循环顺利运转。磷酸烯醇式丙酮酸羧化酶催化磷酸烯醇式丙酮酸与二氧化碳反应生成草酰乙酸,是大肠埃希菌胞内主要回补途径。如图4所示,在MN培养基中添加乙酸钠后,磷酸烯醇式丙酮酸羧化酶酶活略有降低,在4.8 g/L时酶活为未添加乙酸钠时酶活的83%。此后,该酶酶活随着添加乙酸钠质量浓度增加反而逐步升高,在9 g/L时酶活达到未添加乙酸钠时酶活的136%。
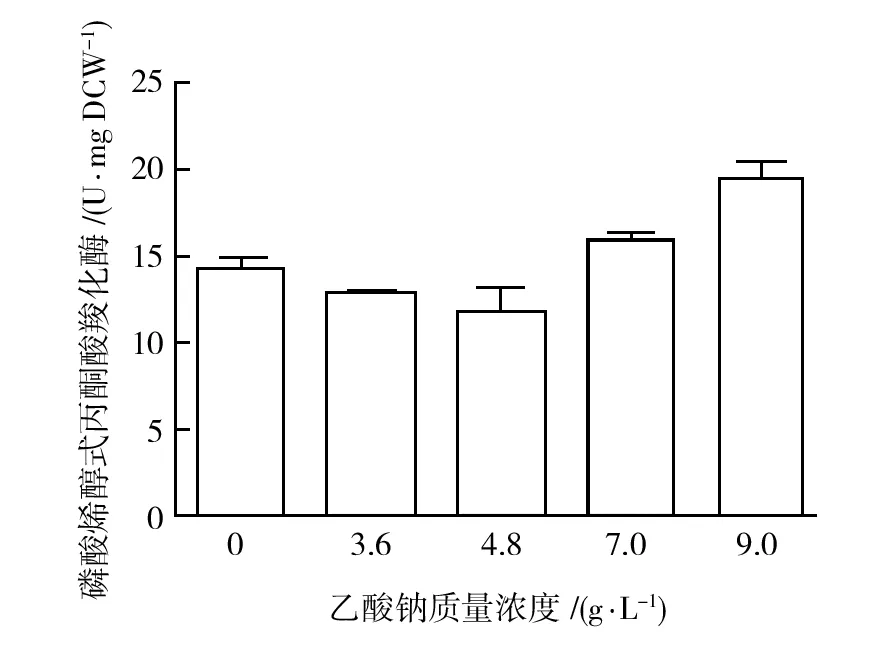
图4 不同外源乙酸钠质量浓度对磷酸烯醇 式丙酮酸羧化酶酶活的影响Fig.4 Effect of the different concentrations of exogenous sodium acetate on the activity of phosphoenolpyruvate carboxylase
2.2.5 乙酸生成途径 在大肠埃希菌体内产生乙酸的主要途径是由磷酸转乙酰酶(PTA)和乙酸激酶(ACK)催化乙酰CoA生成乙酸。如图5所示,在MN培养基中添加3.6 g/L乙酸钠后,乙酸激酶酶活略有降低,该酶在4.8 g/L乙酸钠时酶活进一步降低,为未添加乙酸钠时酶活的66%。随着乙酸钠浓度进一步增加,该酶在7和9 g/L乙酸钠时酶活与3.6 g/L乙酸钠时相当,略低于未添加乙酸钠时酶活。

图5 不同外源乙酸钠质量浓度 对乙酸激酶酶活的影响Fig.5 Effect of the different concentrations of exogenous sodium acetate on the activity of acetate kinase
3 讨 论
大肠埃希菌DA19在MN培养基中进行连续培养时,菌体因过量吸收葡萄糖而生成了乙酸和丙酮酸。与MN培养基相比,添加3.6、4.8和7.0 g/L乙酸钠使得大肠埃希菌DA19自身产生的乙酸明显减少,这与胞内乙酸产生途径的乙酸激酶(ACK)酶活降低结果基本相符,同时与Polen等[12]实验结果一致,Polen认为大肠埃希菌在外源乙酸存在情况下倾向于避免自身乙酸的生成;与乙酸不同,添加3.6、4.8和7.0 g/L乙酸钠使得丙酮酸生成量逐渐增加,这也与Polen等实验结果一致。研究表明,由葡萄糖生成的丙酮酸可以诱导表达小非编码RNA Spot42表达,进而激活主要耐酸性调节因子 RpoS表达,从而在酸性压力条件下保护大肠埃希菌[13]。由此推测,大肠埃希菌DA19增加丙酮酸生成既减少了乙酸生成,又增强了本身乙酸耐受性。与7.0 g/L乙酸钠相比,添加9.0 g/L乙酸钠质量浓度使得乙酸生成量显著提高,而丙酮酸生成量无明显增加,这表明在添加9 g/L乙酸钠情况下,大肠埃希菌DA19可能需要通过乙酸生成途径来提供菌体代谢所需ATP。此外,大肠埃希菌DA19在MN培养基中进行连续培养时,还生成了极少量的延胡索酸,说明在葡萄糖过量情况下,一部分碳源通过PEPC回补途径进入TCA循环,并以延胡索酸形式分泌到胞外,而添加不同浓度乙酸钠明显降低了延胡索酸生成量。
在大肠埃希菌胞内的糖代谢途径主要包括糖酵解途径、磷酸戊糖途径、三羧酸循环、乙醛酸支路、磷酸烯醇式丙酮酸羧化酶回补途径以及乙酸生成途径等。与MN培养基相比,添加3.6和4.8 g/L乙酸钠使得大肠埃希菌DA19胞内PFK、G6PDH、ICDH、ICL、MDH、PEPC和ACK酶活逐渐降低。这可能是因为培养基中乙酸通过扩散作用进入胞内,并迅速发生解离,释放出质子,使得细胞液酸化,从而降低胞内酶活[7]。为了应对胞液酸化问题,大肠埃希菌可以通过谷氨酸、精氨酸和赖氨酸脱羧基反应来减少胞内质子,从而提高胞内pH[14-16]。α-酮戊二酸和草酰乙酸是合成谷氨酸、精氨酸和赖氨酸的前体物质,因此这两个物质会因参与氨基酸脱羧基反应而减少,需要通过回补途径进行补充,这可能是PEPC酶活在7.0和9.0 g/L乙酸钠下增加的原因。在添加7.0和9.0 g/L乙酸钠时,大肠埃希菌DA19胞内ICDH酶活分别为4.8 g/L乙酸钠时的1.17和1.49倍,同时MDH酶活分别为4.8 g/L乙酸钠时的1.19和1.47倍,表明大肠埃希菌DA19胞内的TCA循环在添加7.0和9.0 g/L乙酸钠时明显增强。乙醛酸支路的ICL酶活在添加7.0和9.0 g/L乙酸钠时也有所提高,但远低于相同条件下的异柠檬酸脱氢酶酶活,而且异柠檬酸裂解酶对异柠檬酸的亲和力低于异柠檬酸脱氢酶[17],因此添加乙酸钠并未使大肠埃希菌DA19启动乙醛酸支路,这与文献报道的葡萄糖对异柠檬酸裂解酶存在阻遏作用结果一致[18]。由此推测,大肠埃希菌DA19可能采用了与耐乙酸醋酸菌类似机制来解决胞液酸化问题,即将乙酸流向TCA循环[7],而不是流向乙醛酸支路。
与ICDH、ICL、MDH、PEPC不同,当乙酸钠质量浓度增加至7.0和9.0 g/L时,G6PDH酶活进一步降低,表明磷酸戊糖途径可能与大肠埃希菌DA19的耐乙酸能力增强无关。此外,ACK酶活在7.0和9.0 g/L乙酸钠质量浓度时有所增加,表明此时大肠埃希菌DA19可能需要通过乙酸生成途径来产生ATP。
综上所述,添加不同浓度外源乙酸钠明显改变了大肠埃希菌DA19的生长代谢和糖代谢途径酶活,这些结果有助于了解该菌株对乙酸压力的应答机理。在此基础上,利用双向电泳、表达谱芯片和real-time PCR等技术在蛋白质水平和转录水平上进一步研究该菌株对乙酸压力的应答机理,为应用代谢工程方法构建耐受高浓度乙酸大肠埃希菌提供信息。
[1] Wang H, Wang F, Wang W, et al. Improving the expression of recombinant proteins in BL21 (DE3) under acetate stress: an alkaline pH shift approach[J] . PLoS One, 2014, 9(11):e112777.
[2] Kleman GL, Strohl WR. Acetate metabolism byEscherichiacoliin high-cell-density fermentation[J]. Applied and Envi-ronmental Microbiology, 1994, 60: 3952-3958.
[3] Kim TS, Jung HM, Kim SY, et al. Reduction of acetate and lactate contributed to enhancement of a recombinant protein production inE.coliBL21[J]. Journal of Microbiology and Biotechnology, 2015, 25(7):1093-1100.
[4] Oh MK, Rohlin L, Kao KC, et al. Global expression profiling of acetate-grownEscherichiacoli[J]. Journal of Biological Chemistry, 2002, 277(15):13175-13183.
[5] 朱才庆, 叶勤. 大肠埃希菌DH5α耐乙酸突变株的选育及其代谢特性研究[J]. 微生物学报, 2003, 43(4): 460-465.
[6] Han Y, Li ZM, Du P, et al. Production of human epidermal growth factor in fed-batch culture of acetate-tolerantEscherichiacoli[J]. Chinese Journal of Chemical Engineering, 2007, 15(5): 760-764.
[8] Pan JG, Rhee JS, Lebeault JM. Physiological constraints in increasing biomass concentration ofEscherichiacoliB in fed-batch culture[J]. Biotechnology Letters, 1987, 9:89-94.
[9] Shahinian AH, Reinhold JG. Application of the phenol-hypochlorite reaction to measurement of ammonia concentrations in kjeldahl digests of serum and various tissues[J]. Clinical Chemistry, 1971, 17:1077-1080.
[10]王镜岩, 朱胜庚, 徐长法. 生物化学[M]. 北京:高等教育出版社, 2002:151.
[11]Peng L, Shimizu K. Global metabolic regulation analysis forEscherichiacoliK12 based on protein expression by 2-dimensional electrophoresis and enzyme activity measurement[J]. Applied and Environmental Micorbiology, 2003, 61(2):163-178.
[12]Polen T, Rittmann D, Wendisch VF, et al. DNA microarray analyses of the long-term adaptive response ofEsherichiacolito acetate and propionate[J].Applied and Environmental Micorbiology, 2003, 69:1759-1774.
[13]Wu J, Li Y, Cai Z, et al. Pyruvate-associated acid resistance in bacteria[J].Applied and Environmental Micorbiology, 2014, 80(14):4108-4113.
[14]Richard H, Foster JW.Escherichiacoliglutamate-and arginine-dependent acid resistance systems increase internal pH and reverse transmembrane potential[J]. Journal of Bacteriology, 2004, 186(18):6032-6041.
[15]Iyer R, Williams C, Miller C. Arginine-agmatine antiporter in extreme acid resistance inEscherichiacoli[J]. Journal of Bacteriology, 2003, 185:6556-6561.
[16]Moreau PL.The lysine decarboxylase CadA protectsEscherichiacolistarved of phosphate against fermentation acids[J].Journal of Bacteriology, 2007,189:2249-2261.
[17]Noronha SB, Yeh HJ, Spande TF, et al. Investigation of the TCA cycle and the glyoxylate shunt inEscherichiacoliBL21 and JM109 Using13C-NMR/MS[J]. Biotechnology and Bioengineering, 2000, 68(3):316-327.
[18]Walsh K,Koshland DE Jr.Branch point control by the phosphorylation state of isocitrate dehydrogenase[J]. Journal of Biological Chemistry, 1985, 260(14): 8430-8437.
Effect of Different Concentration Exogenous Sodium Acetate on Metabolism and the Key Enzyme Activity in the Acetate-TolerantEscherichiacoliDA19
ZHANG Yan-jun, ZHANG Ping-hua, ZHOU Can, DONG Man-ming
(Coll.ofChem. &LifeSci.,ZhejiangNormalUni.,Jinhua321004)
In order to study the effect of exogenous sodium acetate on the metabolism ofE.coliDA19, the strain were continuously cultured in nitrogen-limited defined media without as well as with different concentration of sodium acetate. The metabolic parameters and the activities of key enzymes at steady growth state were tested and determined. As compared with nitrogen-limited defined medium, specific glucose consumption rate and specific fumarate production rate ofE.coliDA19 decreased following with the increase of exogenous sodium acetate concentration, and specific pyruvate production rate increased following with the increase of exogenous sodium acetate concentration, while the specific production rate of acetic acid significantly decreased (except 9 g/L sodium acetate). The enzyme activities of phosphofructokinase, isocitrate dehydrogenase, isocitrate lyase, malate dehydrogenase, phosphoenolpyruvate carboxylase and acetate kinase increased and then decreased following with the increase of exogenous sodium acetate concentration. However, the enzyme activities of glucose-6-phosphate dehydrogenase decreased following with the increase of exogenous sodium acetate concentration. In response to the stress of exogenous sodium acetate, distinct changes in enzyme activities happened both in growth metabolism and center metabolic pathways ofE.coliDA19.
Escherichiacoli; acetic acid; enzyme activity
浙江省自然科学基金项目(LQ12C01001);金华市科学技术研究计划项目(2014-2-042)
张艳军 男,助理研究员,博士。研究方向为发酵工程。Tel:0579-82282187,E-mail:yjzhang@zjnu.cn
2015-11-25;
2016-01-24
Q935
A
1005-7021(2016)05-0015-06
10.3969/j.issn.1005-7021.2016.05.003
