肌纤维募集定律的理论溯源及其对力量训练实践的影响
2016-12-14于洪军李成伟
于洪军,李成伟,于 淼
肌纤维募集定律的理论溯源及其对力量训练实践的影响
于洪军1,李成伟1,于 淼2
肌纤维的募集定律是力量训练中的基础理论,从该理论提出的背景入手,在参考原始文献的基础上理解和认识该理论,意在提供更好运用此理论来指导力量运动实践的方法。采用HistCite文献计量学学术史的研究方法,将该理论对竞技体育学术史中的发展影响情况进行分析和讨论。研究提出,在力量训练的实践中,需要遵循大负荷原则、力竭性训练负荷原则,以募集更多肌纤维参与运动,更好地发展力量能力。同时,对募集理论指导下的传统力量和核心力量训练问题展开讨论;研究认为,核心力量训练是一种协调稳定能力的训练,是传统抗阻力量训练的一种有益补充,刺激肌纤维的增长和募集,必须要通过抗阻力量来实现,传统抗阻力量训练对力量能力的提高发挥着不可替代的根本性作用。
肌纤维募集定律;力量训练;核心力量;力竭性训练
引言
当今,世界竞技力量训练理论发展已经进入了一个活跃时期,各种新的力量训练理论、训练方法不断出现,如单组力量负荷训练法、金字塔力量训练法、核心力量训练法、振动力量训练法等。在力量的训练中,任何训练方法和手段,都应该以力量训练过程中肌肉收缩的生物学基础为基石。正如陈小平[1]指出,“运动生物学知识是训练理论的重要基础,它从自然科学的方面对训练理论形成支撑,是制定训练目标和任务、选择训练方法和手段以及控制训练过程和检验训练效果的依据”。因此,我们要站在运动训练生物学基础的视角上,冷静的去分析和鉴别运动训练理论发展过程中的学术观点和争论,对训练基础理论形成和发展的过程有一个全面、客观的认识与把握,以甄别各种训练方法的科学性,实现更加科学地指导训练实践。
肌纤维募集定律(size principle),即肌肉在收缩过程中,运动单位对快肌纤维和慢肌纤维在募集过程中的先后顺序的规律,是人体运动、尤其是力量训练中刺激快肌纤维和慢肌纤维产生适应性变化的基础理论。我国的教练员和运动员在力量训练的过程中,往往着眼于运动的形式,并未深入分析和理解力量训练的生理学基础,出现了诸如强调肌肉的肥大而忽视神经支配、不清楚力量的增长和消退的时间等问题[1,2]。
本文通过对肌纤维募集定律提出的背景、其生理学理论溯源和理论本身发展对相关学术领域的影响及变迁视角重新梳理了该理论,旨在让研究者和读者能够从运动训练的生理学基础上,去认识和指导力量训练实践。
1 肌纤维募集定律的溯源
1.1 理论提出的背景
任何科学研究的发现,都是以前人研究为基础取得新的进步和发现的,肌纤维募集定律的提出也不例外。从运动单位被发现到该理论的提出,大概经历了40年左右的时间(表1)。

表1 肌纤维募集定律提出背景的相关研究历程
1925年,英国牛津大学生理学教授Charles Sherrington等人首次在研究中提出了运动单位的概念[42],即一个脊髓α-运动神经元或脑干运动神经元,和受其支配的全部肌纤维所组成的肌肉收缩的最基本的单位,称为一个运动单位。
1927年,Eggleton等人[27]对青蛙运动研究的实验中,首次发现了不同运动强度下的动物下肢肌纤维募集差异问题。该研究报道,中等强度下青蛙跑步时神经主要募集慢肌纤维(I型)参与运动,最大强度间歇运动(跳跃活动)中主要募集快肌纤维(IIb型)参与运动。虽然Eggleton等人对肌纤维募集的差异有所观察,但当时并未对肌纤维的不同特点进行深入分析和研究。
1929年,作为Charles Sherrington在牛津大学退休前的最后一批学生[60],来自新西兰的Denny-Brown在牛津大学对猫体的下肢生理肌肉实验中,用照相机技术首次发现和区分了肌纤维中的红肌(慢肌)和白肌(快肌)的不同生理特点[24]。
1938年,Denny-Brown和Pennybacker在《横纹肌收缩和运动》(Fibrillationandfasciculationinvoluntarymuscle)一文中,阐述了随着外力的不断增大,肌纤维的收缩按照一定的顺序不断增大,即小的慢肌纤维的运动单位首先动员,随后大的快肌纤维得到募集的现象。此后,Denny-Brown等人并未就此问题进行继续深入研究,他将研究兴趣转到了中枢神经系统上,后来成为了一名著名神经科医生。虽然,该研究仅仅是提出了一个想法和发现了这种现象(与Eggleton等人1927年发现的现象类似),并未在实验中进行量化,也未从生理机制上对该现象进行深入研究。但是,他们的研究报道,极有可能为后来Henneman发现和证实肌纤维的募集定律提供了很大的启发。令人遗憾的是,Henneman在发现和证实肌纤维募集定律的文章中并未引用和提及这个研究[60],这成为了Henneman文章为后来学者所诟病的一大原因。
1941年,Gasser[34]研究报道,神经突触的大小对应着神经冲动频率的高低,即不同大小的神经突触会在神经冲动和募集过程中存在一定的顺序。这个发现为随后电刺激下观察不同负荷刺激中神经冲动的传导奠定了基础。
1957年,Henneman在科学杂志(Science)上报道了电刺激下的神经募集不断增加的现象[37]。在该研究中,作者发现,在电刺激的作用下,随着刺激强度的增加,同侧坐骨神经刺激信号以不断上升顺序被激活和募集,该现象以单个曲肌运动突触(single flexor motor axons)的形式在细胞外神经前单肌纤维(ventral root filament)上被记录下来。当电刺激停止后,突触兴奋冲动信号的释放是以与募集相反的顺序恢复,即首先兴奋的神经突触去募集化最晚,而后兴奋的神经突触去募集化最早。他认为,这种不同大小神经突触兴奋具有一定顺序的现象会适用于所有神经系统。
进入20世纪60年代,生理学领域中的一些关键生物学技术的发明,为Henneman通过技术手段定量研究肌纤维的募集情况提供了可能。1963年,Engel和他的同事们在美国国家健康研究所(National Institues of Health)研发了有关肌肉组织的化学研究技术,为定量研究不同快慢肌纤维特点提供了技术支持[28]。1965年,哈佛大学生理学教授Henneman利用肌肉组织化学技术等手段,在对猫体的肌纤维募集特点的研究中提出了肌纤维的募集定律。
1.2 肌纤维募集定律的提出和核心内涵
Henneman Elwood(1915—1996年),美国哈佛大学医学院运动生理学教授。他最初想成为一名外科医生,主要研究神经生理学,后受洛克菲勒研究所(Rock-efeller institute)David Lloyd的影响,开始对肌肉的神经研究产生兴趣。1957年,Henneman开始对神经募集中的单一运动单位进行深入研究,同年即发现了肌纤维在神经刺激下,随着电刺激强度不断增大,肌纤维募集逐渐增多的现象[37]。1965年,他提出了肌纤维的募集定律[39],该理论认为,在肌肉收缩克服外部阻力的过程中,肌纤维遵循着一定的顺序被募集。运动单位的募集遵循着从小到大的原则,即慢肌纤维首先被募集,然后快肌纤维逐步得到募集。
1965年,Henneman和他的同事们在JournalofNeurophysiology杂志上连续发表了5篇文献介绍神经元的特性、神经肌肉募集特点以及神经肌肉募集过程中的规律,正式提出了肌纤维动员过程中的肌纤维募集定律概念。上述5篇文章发表于同一年,发表时间仅差几个月(表2)。

表2 1965年提出肌纤维募集定律的5篇系列文献
第1篇文章中[43],Henneman 等人首先注意到了红肌收缩速度慢,白肌收缩速度快的不同生理特性。他们对猫体实验研究中发现,相同比目鱼肌和腓肠肌电刺激下的运动单位募集具有“小-大”募集顺序特点。另外,研究还发现,相对于比目鱼肌而言,腓肠肌在高强度的电刺激下更容易产生疲劳。
第2篇文章中[40],Henneman等人在猫体的实验研究中发现了肌纤维的募集具有随着阻力增大,募集数量逐步增多的现象。
第3篇文章中[39],Henneman等人首次系统阐述了肌纤维的募集定律。他们发现,神经突触较小的运动单位首先得到动员和募集,而神经突触较大的运动单位则募集较晚。该文中,他们首先报道了在实验中,切除大脑的猫体其下肢肌纤维在神经的募集中,支配慢肌纤维轴突的神经运动单位首先得到动员和募集,随着外力的不断增大,支配快肌纤维的神经运动单位逐步得到动员。作者在这篇文章的讨论部分,首次正式提出了运动神经元的募集定律概念。
第4篇文献中[38],Henneman等人提出了运动单位惯性(motor unit usage)的概念。他们发现,大的运动神经元支配着白肌纤维,其线粒体和毛细化较少,而小的运动神经元支配着红肌纤维,其线粒体和毛细化较多。他们认为,因为慢肌纤维具有丰富的线粒体和较多的毛细化程度,其抗疲劳能力较强。因此,在运动单位的募集过程中,小的运动单位对神经冲动更加敏感和活跃,更容易被首先得到募集。
第5篇文献中[40],Henneman等人探讨了大的运动单位首先去募集化的问题。他们认为,因为大的运动单位对神经兴奋的敏感程度更高,可能导致兴奋释放后首先得到恢复,由此产生去募集化。在文献中,研究者还讨论了募集定律的意义和价值,并认为,募集定律是神经肌肉系统的独立运行定律,不受其他神经控制的干扰,一旦达到刺激阈值,小的运动单位马上就会得到动员和募集。
Henneman等人1965年有关肌纤维募集定律的系列文章引起了美国国家健康研究所Robert Burke的极大关注。他使用肌糖原耗竭技术,进一步对肌纤维募集过程中的肌纤维募集情况进行了证实和详述,并对不同类型的肌纤维的神经募集顺序给予了更加清晰的展示和证实。更进一步的是,其实验研究发现了大的运动单位支配快肌纤维、小的神经单位支配慢肌纤维的现象[14,15,17]。在此后的25年时间里,Henneman用18篇相关的研究文章在神经生理学杂志上描述了肌纤维的募集定律理论。
对于募集定律的最早提出者方面,学术界有一些小的争议和讨论。虽然,肌纤维的募集定律是1965年由Henneman等人在实验研究首次正式报道,但早在1938年,Denny-Brown等人的研究报道中,就提及了在肌束震颤和脊髓萎缩病人的肱二头肌肌纤维收缩过程中,肌纤维可能存在从小的运动单位到大的运动单位募集的顺序现象[25]。另外,在其1929年的报道中,也提及到了在猫体实验过程中发现红肌(慢肌)纤维早于白肌(快肌)纤维被动员的现象,但并未提及这其中存在着一个不同类型肌纤维募集顺序的原理[24]。鉴于1938年Denny-Brown等人的研究并未在实验中进行定量化研究,以及未在生理实验机制上给予深入研究,而仅仅是提出了一个想法和阐明现象而已,所以,学术界将募集理论的系统提出归功于Henneman教授是恰当的[30]。但是,Henneman在参考文献中未引用Denny-Brown的研究成果,也确有不妥之处。
Henneman等人发表于1965年有关于肌纤维募集定律的5篇系列论文在生理学领域产生了重要的影响。该系列文献不仅提出了诸多神经生理学的重要概念,同时,对这些概念进行了定量化和可视化的科学记录与研究阐述。这些经典研究论文对整个运动生理学领域产生了深远影响,截止到2016年3月10日,以上5篇文献SCI他引达到3 117次。
1.3 肌纤维募集定律概念提出:原文导读
在以上的5篇文献中,1965年,Henneman在JournalofNeurophysiol杂志上以“Functionalsignificanceofcellsizeinspinalmotoneurons”一文,首次系统阐述了肌肉的募集定律[39]。研读21页的原文文献发现,作者通过对平均重量为3.2 kg,35个成年切除大脑的实验猫体的165对下肢肌纤维中不同类型运动单位(快肌和慢肌)的肌纤维募集实验进行研究发现,肌纤维的神经元在快肌和慢肌纤维之间存在着一定的募集模式,不同类型神经元支配下的肌纤维具有不同的募集阈值,即小的神经元募集阈值较低,首先得到募集;大的神经元阈值较高,后得到募集。其中,实验猫体的165对肌纤维中,142.5对遵循小大原则,22.5对(因数据统计原因,故有0.5对)募集顺序则相反。基于实验结果,作者提出了肌纤维募集定律。
在文中,作者对该研究结果的报道分为了引言、实验方法、结果、讨论和结论5个部分进行报道。引言部分探讨了他在之前几篇文献报道中所述的不同类型肌纤维具有不同兴奋阈值的现象,即1957年作者在Science发表的“Relationbetweensizeofneuronsandtheirsusceptibilitytodischarge”一文中提到的不同神经元之间不同兴奋阈值的问题[37]。以及1965年,作者与同事共同发表于JournalofNeurophysiol杂志的“Propertiesofmotorunitsinahomogeneousredmuscle(soleus)ofthecat”[43]和“Propertiesofmotorunitsinaheterogeneouspalemuscle(m.gastrocnemius)ofthecat”[61]两篇文献中分别就受试猫肌纤维中的红肌(慢肌)和白肌(快肌)不同募集顺序问题进行的说明。
探讨快慢肌纤维的发现是该研究的基础。一个值得注意的事件是,作者引用了Gasser 1941年对于快肌纤维和慢肌纤维支配神经的直径不同的研究结论,即快肌纤维由直径较大的神经支配,慢肌纤维则由直径较小的神经支配。但快、慢肌纤维的首次发现是1938年由Denny-Brown等人对有关肌纤维募集观察的研究报道[60],而且,Denny-Brown在该文中也提及到了不同肌纤维募集顺序的问题。但令人遗憾的是,Henneman在其研究中并未引用该研究文献[29,60]。
在实验结果部分,作者首先对比了在收缩和放松阶段2个不同大小运动神经元在收缩和放松阶段的激活顺序(图1)。在收缩阶段,小的运动单位神经支配下的肌纤维首先被激活,大的运动单位肌纤维随后被募集;在放松阶段,大的神经元首先停止激活,得到放松,小的运动单位肌纤维最后得到放松。
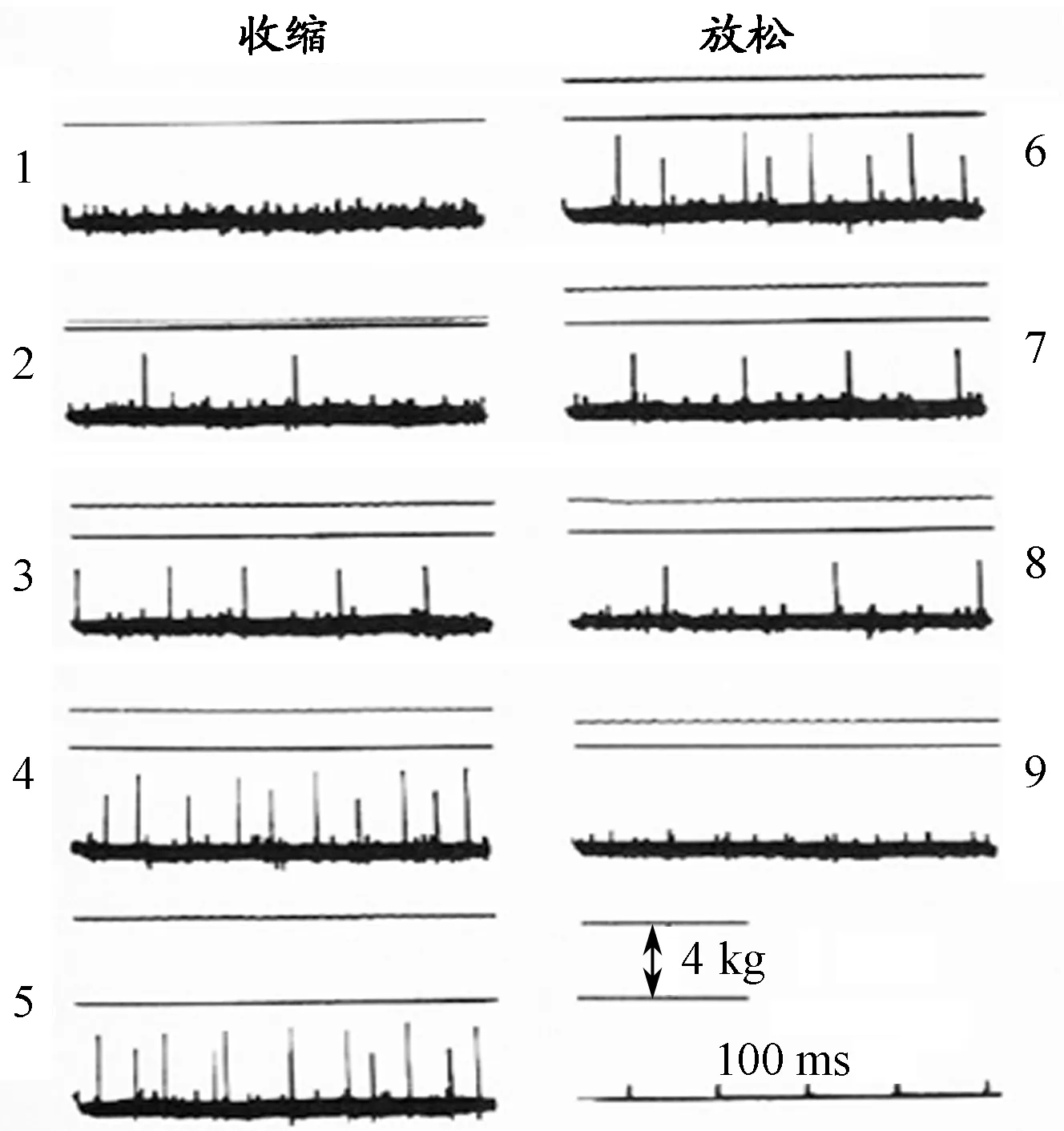
图1 不同大小运动单位(2对)
随后,作者在文中又展示了5对(不同大小)运动单位神经支配下的肌纤维募集的顺序情况(图2)。研究发现,收缩时,募集的顺序是由小到大(1,2,3,5,4);放松时,顺序是由大到小(5,4,3,2,1)。基于这个发现,作者提出了肌纤维神经募集定律,即肌纤维募集过程中,存在着小-大现象,即小的运动单位首先得到募集,而大的运动单位反之。
在下文中,作者以平均体重3.2 kg(1.8~5.0 kg)、切除大脑的35只成年受试猫为受试对象,研究其下肢165对不同类型的肌纤维进行神经肌肉募集情况(图3)。实验结果表明,在实验过程中,165对不同型号肌纤维的募集过程,其中,142.5对遵循着小-大原则,即神经元小的肌纤维首先得到募集,大的神经元支配下的肌纤维后得到募集,剩余的22.5对没有遵循这一小-大募集原则。如图4所示,不同大小运动单位的肌纤维募集阈值不同,小的运动单位募集阈值较小,大的运动单位募集阈值较大。

图2 不同大小运动单位(5对)
在讨论部分中,Henneman正式提出了肌纤维募集定律的概念:在肌肉收缩过程中,形态较小的运动单位肌纤维(慢肌)首先得到募集,形态大的运动单位支配下的肌纤维(快肌),最后才能够被募集。同时作者认为,这一定律可以应用到人体所有肌纤维的募集中。之后,作者也探讨了肌纤维型号与其兴奋阈值之间的关系问题,提出了肌纤维的兴奋阈值也遵循着小-大的原理,即慢肌纤维兴奋阈值较低,快肌纤维兴奋阈值较高。最后,作者探讨了不同神经冲动1~20 u下的肌纤维兴奋和激活问题,认为慢肌纤维神经元的兴奋阈值在1~8 u之间,快肌纤维的兴奋阈值在9~20 u之间。如果需要让快肌纤维得到兴奋激活,其外部负荷和肌纤维的兴奋阈值需要达到9 u以上。在最后的总结部分,作者对该实验中的肌纤维募集原理,总结为小-大原则——慢肌纤维首先得到动员和兴奋,快肌纤维最后才能够被动员起来。
2 肌纤维募集理论的影响和发展
在肌纤维募集理论的理论发展脉络和理论影响上,本文以1965年Henneman等人1965年正式提出该理论的“Functi-onalsignificanceofcellsizeinspinalmotoneurons”一文为基础,对该理论在学术史中的影响和发展脉络进行文献分析[39]。在文献分析中,本研究采用了HistCite软件,文献分析的数据来源于SCI-E数据库。数据库文献截至到2016年1月1日,该文已被他引1 307次。本文在数据库中下载了引用该文献的1 307篇文献的标题、作者、摘要、参考文献等信息,以进行原始数据分析。

图3 不同大小肌纤维的募集顺序之间的关系示意图

图4 3次实验中不同阈值运动单位的肌纤维募集示意图
2.1 学术影响
2.1.1 他引次数
截至到2016年1月1日,基于SCI数据库中引用该文的1 307篇文献分析可以看出,Henneman自1965年提出肌肉的募集定律以来,该理论就逐步得到学界的重视。如图5所示,从1965年提出至2015年,肌纤维募集理论一直得到学术界的引用和关注。

图5 肌纤维募集定律理论论文他引数量变化趋势图
2.1.2 肌纤维募集定律所影响的学术领域
在通过对他引文献的前10个研究领域的分析中可以看出,肌纤维募集定律理论的影响范围主要在神经科学、生理学和体育科学3个领域。其中,体育科学领域中引用该理论的文献数为254篇,占全部他引文献的19.4%,位于该理论影响领域的第3位(表3)。

表3 肌纤维募集定律影响排名前10名研究领域分布
2.1.3 体育科学领域学术影响
直到1976年,Henneman于1965年所发现的肌纤维募集定律理论才被体育科学领域关注和引用。如图6所示,对254篇体育科学领域中引用该文献的数据进行分析表明,该理论自20世纪90年代之后,一直在体育科学领域得到关注,2014年,体育科学领域引用该文献的论文数达到17篇。
2.2 理论发展影响脉络:基于HistCite文献计量学可视化分析
本文采用HisCite的文献计量学分析方法,对体育科学领域中,该理论的影响脉络进行了深入分析。

图6 肌纤维募集定律理论在体育
HistCite的研究方法简介如下:HistCite=history of cite,意味引文历史,或引文图谱分析软件。该软件系SCI的发明人Garfield开发[33],能够用图示的方式展示某一领域不同文献之间的关系。HistCite可以快速帮助研究者绘制出一个领域的发展历史图示,定位出该领域的重要文献以及最新的核心文献。在文献数据中,GCS(global citation score),即引用次数,也即SCI-E数据库存的引用次数。LCS是local citation score的简写,即本地引用次数;与GCS相对应,GCS是总被引次数。LCS是某篇文章在当前数据库中被应用的次数,故LCS一定是小于或等于GCS。若一篇文章GCS很高,说明该文章被全球科学家关注较多,但如果一篇GCS很高,而LCS很小,说明这种关注可能主要来自于同一领域的科学家。
自肌纤维的募集定律于1965年被提出后,该理论在体育科学领域中的影响学术史可视化图谱如图7所示。1984年,Citterio等[21];1987年,Moritani等[46]和1987年,Sale[54]的3篇文章在体育科学领域分别从人体自行车功率、自行车的肌纤维募集、人体肌电测试和力量训练中的肌纤维募集视角对募集定律进行了测试和实践运用。进入20世纪90年代后, Moritani等[48]于1992年对人体在运动中肌肉募集的氧运输等方面进行了实验验证,属于20世纪90年代引用该理论比较有影响的研究文献。进入21世纪以来,2000年,Takarada等[59]对力量训练中肌纤维的募集原则进行了实验分析,该论文在本学科内被引用17次(截止于2016年1月1日)。2004年,Moore等[45]报道了低强度下力量训练中肌纤维募集和血流的影响,这对该理论的进一步发展产生了重要影响。2007年,Fujita等[31]报道了低强度力量训练中血流和肌肉蛋白影响有关肌纤维募集的文章,再次对该理论的发展起到了重要作用。2013年,Schoenfeld等[56]就力量训练中代谢压力下的高原适应问题进行论述,再次提及肌纤维的募集经典理论。

图7 基于HistCite的肌纤维募集定律
3 肌纤维募集定律对力量训练的影响和发展
Thomas Delorme是首位将力量训练带入到科学研究的轨道上来的学者[23]。1945年,他在二战期间负责伤病人员的恢复训练,对伤员进行了不超过10 次(7~10次/天)的最大负荷力量训练刺激后发现,伤员的受训练肌肉的力量迅速提高,之后,力量训练成为了一个全新的研究领域,越来越多的基础科学尤其是生理学基础理论逐步被应用到力量训练中。1962年,Bergstrom[13]发明了肌肉活检技术,为定量研究人体肌纤维收缩前、中和后期肌纤维的募集变化提供了可能。随着该技术的发明和应用,科学家对于人体运动过程中神经肌纤维募集顺序原理的发现和研究逐步深入。1973年,Stein等[44]首次在人体试验中证实了肌纤维的募集定律。Philip Gollnick等人报道了人体不同强度下肌纤维募集的研究结果,在其研究报道中,在次最大运动强度下,慢肌(I型)肌纤维主要得到募集,在超最大运动强度(即超过最大摄氧量的强度)下,快肌纤维(II型)逐步得到募集[35,36]。1981年,Burke[16]提出了基于肌纤维的神经和肌纤维组织化学特性,包括肌纤维收缩时间和疲劳特性,提出了4种肌纤维类型的另一种分类方法。在这样的分类方法下,他认为,Henneman提出的慢肌纤维首先被募集的肌纤维募集定律仍然适用。Kreamer等[41]、Ratamess等[51],都将肌纤维的募集定律视为力量训练的重要生理基础理论。以上研究成果,为肌纤维的募集定律成为指导力量训练的重要基础理论提供了条件。基于募集定律,在力量训练实践中,需要遵循以下3个训练原则。

表4 基于Histcite的肌纤维募集定律在体育科学领域不同年代的代表性学术论文
3.1 大负荷原则
肌纤维的募集定律对力量训练实践具有极其重要的意义。据该理论,可以推断出力量训练负荷(强度)在由低到高的增长过程中对不同运动单位的影响和作用。低负荷和慢速度的训练只能动员慢运动单位(I型),而快肌单位(II型)的动员则需要通过大强度的负荷刺激,如欲让所有运动单位都得到募集和训练,就必须运用大负荷、高强度的训练方法[49]。有研究表明,在力量训练中,小的负荷(≤40%最大力量)只能够动员慢收缩氧化肌纤维(ST)参与运动,随着负荷的增加,快收缩氧化-糖酵解肌纤维(FTO)和快收缩糖酵解肌纤维(FTG)也开始被动员起来,直到约95%的运动单位都被募集[3]。根据力量的分类,力量素质分为最大力量、快速力量、力量耐力和反应力量[4],其中,力量的负荷强度是进行不同类型力量素质训练和决定力量训练效果的关键指标。根据不同的训练目标,应进行合理的力量训练安排,以保证具体训练目标的实现。例如,爆发力训练目标、最大力量训练目标和肌肉耐力的训练目标是不同的,疲劳的肌纤维在爆发力和最大力量的训练中是无效的训练刺激。
有研究报道[12],在高重复性力量(>125次的重复)和耐力训练中,除非慢肌纤维中的肌糖原等能量耗竭,快肌纤维不会被募集,其力量的提高幅度较小。低负荷和高重复次数的力量训练(<20% 1RM)对快肌纤维的刺激很小,力量提高也非常有限。Campos等[19],Schuenke等[58]的研究报道指出,25~30 RM的低负荷力量训练不会刺激II型肌纤维的发展和肌纤维的肥大适应。只有8~11 RM,3~5 RM或者超过90% 1RM的力量训练才能使得肥大性肌纤维神经冲动得以产生,达到神经冲动的最低阈值,募集快肌纤维的参与,而超过20 RM的力量训练不能够达到这种训练刺激的效果。Schoenfeld等[57]在研究综述提到,一般而言,60% 1RM以上的训练负荷刺激强度能够促进肌纤维肥大适应性变化,30% 1RM的训练负荷重量是保证快肌纤维募集的最低负荷强度。
陈小平研究认为[5],基于肌纤维募集定律,在力量训练中需要充分重视“神经”支配能力的训练,训练负荷是训练方法的一个重要组成部分,在训练方式确定之后采用什么样的练习量(次数和组数等)和练习强度(重量、速度和间歇时间等),对所训练肌肉力量的发展方向将产生决定性的作用。在力量训练中,如果分别考虑神经与肌肉系统对力量的功能和作用,根据专项和运动员个体情况有针对性地改善力量水平,则必须对传统的力量训练负荷原则做进一步细化。从负荷重量的角度,以最大力量的90%为界,将发展最大力量的负重(75%~100% 1RM)进一步划分为两个不同的功能区域:优先发展运动神经募集能力的最大负荷(90%~100% 1RM)和优先发展肌肉横断面积的次最大负荷(75%~90% 1RM)。只有负荷强度达到一定的阈值,即90% 1RM负荷刺激,才能够动员到快肌纤维参与运动,以提高运动员的最大力量。另外,大负荷的快速力量训练能够更好的募集快肌纤维参与运动。
Cope等[22]人的研究综述显示,在有关募集理论的研究报道中,有研究结果遵循Henneman提出的肌纤维募集定律,即募集顺序遵循从小到大(先募集慢肌后募集快肌)的原则(图8A);但也有研究报道指出,在人体多关节参与的试验中[18],由于神经支配因素的影响[32,48],肌纤维发生选择性募集,即神经支配下肌纤维募集中出现部分小的慢肌纤维始终没有得到募集,而大的快肌选择性被募集的现象,但此募集顺序仍旧遵循从小到大的原则(图8B)。Cope等[25]认为,关于募集定律的研究虽有一些不同研究报道结果,但肌纤维的募集定律是肌肉收缩过程中的基础定律,该定律在解释力量训练和肌肉收缩过程中肌纤维募集原理仍然普遍适用。同时,他认为,肌纤维的募集现象在大负荷刺激或多关节运动中,由于神经因素的参与,在神经支配下,可能会发生选择性肌纤维募集现象。

图8 不同肌肉收缩速度下的肌纤维募集情况示意图
基于以上的研究,我们认为,Henneman提出的“小-大”原则在解释递增负荷下肌纤维募集仍然被认可,但在神经参与支配和高强度特殊运动情况下的肌纤维募集,该理论得到最新一些研究的挑战,该经典理论并不能完全解释所有募集现象,即存在图8B表示的快肌纤维选择性优先募集的情况。需要注意的是,大负荷训练原则在青少年阶段要谨慎使用[11]。
3.2 力竭性训练负荷刺激原则
肌纤维在募集过程中,负荷阻力的时间也是决定肌纤维募集多少的一个重要因素。随着人体克服外部阻力不断增大需要募集越来越多的肌纤维参与运动。每一个运动单元都支配着不同数量和不同类型的肌纤维,其募集原理均遵循着慢肌纤维首先得到募集的原则。Carpinelli认为[20],在力量训练中,瞬时肌肉疲劳刺激能够更好的募集高阈值的快肌纤维运动单元参与,对力量的增加和肌纤维肥大适应效果最佳。Rooney等[53]人研究报道,单组大负荷肌纤维疲劳性动态性(力竭性)力量训练刺激的训练效果要高于多组次最大负荷刺激的训练效果,其两组之间的力量增长分别是41.2%和19.7%。Drinkwater等[26]人报道,力竭性动态性力量训练比非力竭性力量训练,其训练效果更好,其中,两组力量分别提高9.5%和5%。在这些力量训练中,包括了单组6次最大重复的力量训练刺激[53];4组,每组6次最大重复的训练刺激[26]和每组30 s,4组静力性训练刺激。其中,每组练习的次数是力量训练中的重要因素,在肌纤维的募集定律中,只有每组练习的次数达到力竭才能够募集更多的肌纤维参与训练和提高[52]。以上的研究均表明,力竭性训练刺激的训练效果高于非力竭性训练刺激训练。另外,为了达到力竭性力量训练刺激原则,需要在力量训练中进行多次数和多组数刺激。根据肌肉的募集定律,快肌纤维的动员,除需要足够的负荷刺激之外,还需要足够的时间和足够的组数才能得到募集和动员。陈小平[3]认为,当参与运动的运动单位被动员起来后,还需要进一步提高已经被动员起来的运动单位的兴奋性。Sale等[55,63]人研究报道,肌肉收缩速度的改
变首先取决于运动单位的募集数量,当肌肉以20%的最大力量收缩时,可以动员约40%参与运动肌肉的运动单位,当肌肉力量达到最大力量的50%时,大约所有应该参与运动的运动单位都被动员起来。此后,肌肉收缩力量的进一步提高主要是通过提高运动单位的兴奋冲动频率,而不能或很少通过运动单位的募集。此外,不同训练的组合能够发挥肌纤维的募集定律参与工作。采用上肢-下肢或下肢-上肢力量练习的顺序,能够使得更多的肌纤维得到募集和训练刺激。例如,在爆发力的竖直跳训练之前,先进行5 RM的半蹲训练,可使训练的效果最好。
基于以上的神经肌肉募集的特性,在力量训练中,如果需要让快肌纤维得到充分的募集,需要进行多次数和多组数的刺激负荷训练。在具体的训练方法上,力量训练中的单组重复力竭训练方法就是利用了神经肌肉募集这一原理而设计的训练方法。
3.3 抗阻力量和核心力量的关系
核心力量的出现,带来了对传统抗阻力量的冲击和质疑,传统抗阻力量和核心力量之间的关系问题是目前运动训练中一个具有争议的话题。
核心力量的概念来源于康复训练,由Pope等[50]于1985年首次提出。“核心力量是指人体核心部位(腰椎-骨盆-髋关节)的肌肉,以稳定人体核心部位、控制重心运动、传递上下肢力量为主要目产生的力量能力,它主要受神经支配能力、核心部位肌肉的支撑力量能力和呼吸与运动之间的配合等3方面因素的影响”[9]。核心力量的英文概念是“core stability”,即核心稳定性,准确来讲是属于一种稳定性能力[8],这种稳定性能力,是腰腹小肌群之间传递力量的一种“协调、稳定和传递力”的能力,不属于传统的抗阻力量能力,也不是人体力量增长的全部,它是大肌群肌肉力量发力和力量协调传递的保障和补充,因此,核心力量和传统抗阻力量二者之间不存在矛盾关系。有国内学者对核心力量的肌纤维募集情况进行了实证研究。黎涌明等[7]人通过对13名男性大学生运动员在非稳定支撑面上自由负重练习的肌电分析研究表明,在30% 1RM、60% 1RM负重下非稳定的肌分肌电值并不显著。该研究认为,非稳定性训练不能代替负重抗阻训练对发展肌肉横断面、最大力量和爆发力的作用。刘瑞东等[10]人通过对14名足球项目大学生运动员在稳定与多级非稳定徒手和30% 1RM负重深蹲测试研究结果发现,无论在稳定和非稳定的状态下,“徒手”训练基本不造成参与活动肌肉力量的变化,而负重可以显著改变肌纤维的募集,提高其力量能力,该研究也提出,核心稳定力量不能代替传统力量抗阻训练。洪扬等[6]人对19名无训练经历的大学生徒手和30% 1RM稳定和非稳定平面负重深蹲实验研究结果表明,大部分肌肉在30% 1RM负重情况下,稳定和不同级别非稳定状态中肌电之间没有显著性差异。本研究同样认为,非稳定条件不是造成肌肉力量和用力模式改变的原因,负重是提高参与工作肌肉活性的重要因素。
在力量训练中,负荷强度是提高力量能力的最主要因素。有研究报道,60% 1RM以上的训练负荷刺激强度能够促进肌纤维肥大适应性变化,30% 1RM的训练负荷重量是保证快肌纤维募集的最低负荷强度[57]。根据肌纤维的募集定律,负荷是导致大的快肌纤维逐步得到募集的主要因素。根据以上的研究报道可以看出,非稳定性的不平衡并不能提高肌纤维的募集,在其他条件一致的情况下,负重抗阻训练的肌纤维募集效果要优于徒手非稳定性练习。从训练的效果来看,核心力量训练的主要贡献是通过非稳定性条件的刺激能够通过动员深层小肌肉(群)的募集和协调,弥补传统抗阻力量训练中对深层小肌肉(群)的“短板”效应,提高肌肉链的衔接和传递,以促使力量能力得到最大程度的发挥。根据肌纤维的募集定律,抗阻力量训练是提高肌纤维募集的主要因素,而非稳定的核心力量训练不能动员大肌群的快肌纤维得到募集,长期进行核心力量训练而不进行抗阻力量训练会导致大肌群力量下降。综上所述,核心力量训练是一种协调稳定能力的训练,是传统抗阻力量训练的一种有益补充,刺激肌纤维的增长和募集,必须要通过抗阻力量来实现,传统抗阻力量训练对力量能力的提高发挥着不可替代的根本性作用。
4 结语
重塑运动训练学的生物学基础,深入研究和理解运动训练的经典理论是体育科学研究学者的责任和使命。对于各种层出不穷的训练方法和训练手段,唯有从基础理论去理解和甄别,从经典理论的源头去把握,才是训练学学术研究的基础路径。从肌纤维募集定律的提出至今,已经经历了半个多世纪。本文从该理论提出的背景、原始文献的视角出发,对该理论进行了梳理。在认识和理解该理论的基础上,对力量训练中需要遵循大负荷和力竭性力量训练的原则从该理论的基础上进行了阐释,并对传统抗阻和核心力量关系问题进行探讨,认为基于肌纤维的募集定律,核心力量不能代替传统抗阻力量训练。
[1]陈小平.重塑我国训练理论的运动生物学基础[J].体育科学,2010,30(11):17-23.
[2]陈小平.生物学和教育学理论基础的建设——我国运动训练理论亟待解决的问题[J].体育科研,2011,32(4):19-20.
[3]陈小平.当代运动训练热点问题研究[M].北京:北京体育大学出版社,2005.
[4]陈小平.力量训练的发展动向与趋势[J].体育科学,2004,24(9):36-40.
[5]陈小平.“神经支配能力”的训练——我国力量训练一个亟待解决的问题[J].中国体育教练员,2004,(4):25-26.
[6]洪扬,刘瑞东,陈小平.基于肌电均方根和包络线的稳定与多级非稳定条件下徒手与负重深蹲的肌电特征研究[J].体育科学,2016,36(3):67-73.
[7]黎涌明,曹春梅,陈小平.非稳定支撑面上自由负重练习的肌电分析[J].体育科学,2012,33(6):39-43.
[8]黎涌明,徐权,陈小平.“核心力量”和力量训练刍议[J].中国体育教练员,2011,(4):38-41.
[9]黎涌明,于洪军,资薇,等.论核心力量及其在竞技体育中的训练——起源·问题·发展[J].体育科学,2008,28(4):19-29.
[10]刘瑞东,洪扬,陈小平.稳定与多级非稳定条件下徒手与负重深蹲的肌电特征研究及其对当前力量训练的启示[J].体育科学,2015,35(8):45-51.
[11]于洪军,任越.青少年力量训练:机制、方法与趋向[J].体育科学,2015,35(8):76-85.
[12]ANDERSON T,KEARNEY J T.Effects of three resistance training programs on muscular strength and absolute and relative endurance[J].Res Q Exe Sport,1982,53(1):1-7.
[13]BERGSTROM J.Muscle electrolytes in man-determined by neutron activation analysis on needle biopsy specimens -study on normal subjects,kidney patients,and patients with chronic diarrhoea[J].Scand J Clin Lab Inv,1962,1468:1.
[14]BURKE R E.Motor unit types of cat triceps surae muscle[J].J Physiol,1967,193(1):141-160.
[15]BURKE R E.Group ia synaptic input to fast and slow twitch motor units of cat triceps surae[J].J Physiol,1968,196(3):605-630.
[16]BURKE R E.Motor units:Anatomy,physiology,and functional organization[J].Handbook of Physiol,1981,(1):345-422.
[17]BURKE R E,LEVINE D N,ZAJAC F,etal.Mammalian motor units:Physiological-histochemical correlation in three types in cat gastrocnemius[J].Sci,1971,174(4010):709-712.
[18]BUTLER T J,KILBREATH S L,GORMAN R B,etal.Selective recruitment of single motor units in human flexor digitorum superficialis muscle during flexion of individual fingers[J].J Physiol,2005,567(1):301-309.
[19]CAMPOS G E,LUECKE T J,WENDELN H K,etal.Muscular adaptations in response to three different resistance-training regimens:Specificity of repetition maximum training zones[J].Eur J Appl Physiol,2002,88(1-2):50-60.
[20]CARPINELLI R N.Challenging the american college of sports medicine 2009 position stand on resistance training[J].Med Sport,2009,13(2):131-137.
[21]CITTERIO G,AGOSTONI E.Selective activation of quadriceps muscle-fibers according to bicycling rate[J].J Appl Physiol,1984,57(2):371-379.
[22]COPE T C,PINTER M J.The size principle:Still working after all these years[J].News Physiol Sci,1995,10(6):280-286.
[23]DELORME T L.Restoration of muscle power by heavy-resistance exercises[J].J Bone Joint Surg Am,1945,27(4):645-667.
[24]DENNY-BROWN D E.The histological features of striped muscle in relation to its functional activity[J].Proc R Soc Lond B Biol Character,1929,104(731):371-411.
[25]DENNY-BROWN D,PENNYBACKER J B.Fibrillation and fasciculation in voluntary muscle[J].Brain,1938,61(3):311-312.
[26]DRINKWATER E J,LAWTON T W,LINDSELL R P,etal.Training leading to repetition failure enhances bench press strength gains in elite junior athletes[J].J Strength Condit Res,2005,19(2):382-388.
[27]EGGLETON P,EGGLETON G P.The inorganic phosphate and a labile form of organic phosphate in the gastrocnemius of the frog[J].Biochem J,1927,21(1):190-195.
[28]ENGEL W K.Adenosine triphosphatase of sarcoplasmic reticulum triads and sarcolemma identified histochemically[J].Nature,1963,200(4906):588-589.
[29]FUGLEVAND A J.Henneman's size principle:The right name[J].Sci,1998,281(5379):919.
[30]FUGLEVAND A J.Henneman's size principle:The right name[J].Sci,1998,281(5379):919.
[31]FUJITA S,ABE T,DRUMMOND M J,etal.Blood flow restriction during low-intensity resistance exercise increases S6K1 phosphorylation and mMuscle protein synthesis[J].J Appl Physiol,2007,103(3):903-910.
[32]GARNETT R,STEPHENS J A.Changes in the recruitment threshold of motor units produced by cutaneous stimulation in man[J].J Physiol,1981,311(1):463-473.
[33]GARFIELD E.From the science of science to scientometrics visualizing the history of science with histcite software[J].J Informetrics,2009,3(3):173-179.
[34]GASSER H S.The classification of nerve fibers[J].Ohio J Sci,1941,41(3):145-159.
[35]GOLLNICK P D,ARMSTRONG R B,SEMBROWICH W L,etal.Glycogen depletion pattern in human skeletal muscle fibers after heavy exercise.[J].J Appl Physiol,1973,34(5):615-618.
[36]GOLLNICK P D,PIEHL K,SALTIN B.Selective glycogen depletion pattern in human muscle fibres after exercise of varying intensity and at varying pedalling rates[J].J Physiol,1974,241(1):45-57.
[37]HENNEMAN E.Relation between size of neurons and their susceptibility to discharge[J].Sci,1957,126(3287):1345-1347.
[38]HENNEMAN E,OLSON C B.Relations between structure and function in the design of skeletal muscles[J].J Neurophysiol,1965,28(3):581-598.
[39]HENNEMAN E,SOMJEN G,CARPENTER D O.Functional significance of cell size in spinal motoneurons[J].J Neurophysiol,1965,28(3):560-580.
[40]HENNEMAN E,SOMJEN G,CARPENTER D O.Excitability and inhibitibility of motoneurons of different sizes[J].J Neurophysiol,1965,28(3):599-620.
[41]KRAEMER W J,ADAMS K,CAFARELLI E,etal.Joint posi-tion statement:Progression models in resistance training for healthy adults[J].Med Sci Sport Exer,2002,34(2):364-380.
[42]LIDDELL E,SHERRINGTON C S.Recruitment and some other features of reflex inhibition[J].Pro R Soc Lond B Biol Character,1925,97(686):488-518.
[43]MCPHEDRAN A M,WUERKER R B,HENNEMAN E.Properties of motor units in a homogeneous red muscle(Soleus) of the cat[J].J Neurophysiol,1965,28(1):71-84.
[44]MILNERBR.H S,STEIN R B,YEMM R.Orderly recruitment of human motor units during voluntary isometric contractions[J].J Physiol-London,1973,230(2):359-370.
[45]MOORE D R,BURGOMASTER K A,SCHOFIELD L M,etal.Neuromuscular adaptations in human muscle following low intensity resistance training with vascular occlusion[J].Eur J Appl Physiol,2004,92(4-5):399-406.
[46]MORITANI T,MURO M.Motor unit-activity and surface electromyogram power spectrum during increasing force of contraction[J].Eur J Appl Physi Occup Physi,1987,56(3):260-265.
[47]MORITANI T,SHERMAN W M,SHIBATA M,etal.Oxygen availability and motor unit-activity in humans[J].Eur J Appl Physi Occup Physi,1992,64(6):552-556.
[48]NARDONE A,ROMANC,SCHIEPPATI M.Selective recruitment of high-threshold human motor units during voluntary isotonic lengthening of active muscles[J].J Physiol,1989,409(1):451-471.
[49]PETERSON M D,RHEA M R,ALVAR B A.Maximizing strength development in athletes:A meta-analysis to determine the dose-response relationship[J].J Strength Condit Res,2004,18(2):377-382.
[50]POPE M H,PANJABI M.Biomechanical definitions of spinal instability[J].Spine(Phila Pa 1976),1985,10(3):255-256.
[51]RATAMESS N A,ALVAR B A,EVETOCH T E,etal.Progression models in resistance training for healthy adults[J].Med Sci Sport Exer,2009,41(3):687-708.
[52]RHEA M R,ALVAR B A,BURKETT L N.Single versus multiple sets for strength:A meta-analysis to address the controversy[J].Res Q Exe Sport,2002,73(4):485-488.
[53]ROONEY K J,HERBERT R D,BALNAVE R J.Fatigue contributes to the strength training stimulus[J].Med Sci Sport Exe,1994,26(9):1160-1164.
[54]SALE D G.Influence of exercise and training on motor unit activation[J].Exe Sport Sci Rev,1987,15(1):95-151.
[55]SALE D G.Neural adaptation to resistance training[J].Med Sci Sport Exe,1988,20(S5):135-145.
[56]SCHOENFELD B J.Potential mechanisms for a role of metabolic stress in hypertrophic adaptations to resistance training[J].Sports Med,2013,43(3):179-194.
[57]SCHOENFELD B J,ARAGON A A,KRIEGER J W.The effect of protein timing on muscle strength and hypertrophy:A meta-analysis[J].J Int Soc Sport Nutr,2013,10(1):1-13.
[58]SCHUENKE M D,HERMAN J R,GLIDERS R M,etal.Early-phase muscular adaptations in response to slow-speed versus traditional resistance-training regimens[J].Eur J Appl Physiol,2012,112(10):3585-3595.
[59]TAKARADA Y,TAKAZAWA H,SATO Y,etal.Effects of resistance exercise combined with moderate vascular occlusion on muscular function in humans[J].J Appl Physiol,2000,88(6):2097-2106.
[60]VILENSKY J A,GILMAN S.Renaming the henneman size principle[J].Sci,1998,280(5372):2027.
[61]WUERKER R B,MCPHEDRAN A M,HENNEMAN E.Properties of motor units in a heterogeneous pale muscle(M.Gastrocnemius) of the Cat[J].J Neurophysiol,1965,28(1):85-99.
[62]ZEHR E P,SALE D G.Ballistic movement-muscle activation and neuromuscular adaptation[J].Can J Appl Physiol,1994,19(4):363-378.
The Original of Size Principle and Its Influence on Strength Training
YU Hong-jun1,LI Cheng-wei1,YU Miao2
Size principle is very important basically theory for strength training.This paper tries to review the background and the original
related to the size principle,the main purpose is to deepen understand the theory of size principle and to inspire the strength training practice.Applying HistCite software,we analyzed the development and influence of the size principle in sport science field.This paper finally put forward that in strength training,it is necessary to follow the principle of heavy load,the principle of exhaustive training load,in order to recruit more muscle fibers to participate in sports,better develop strength ability.At the same time,traditional strength and core strength training are discussed under the guidance of recruitment theory.It suggests that core strength training is a kind of coordination and stability training,it is one of the beneficial supplements of traditional resistance strength training.Stimulating the growth of the muscle fibers and recruitment must be done by resistance force.Traditional resistance training plays a fundamental role to improve the strength power.
sizeprinciple;strengthtraining;corestrength;exhaustivetraining
1000-677X(2016)11-0056-10
10.16469/j.css.201611007
2016-04-05;
2016-10-31
国家社会科学基金一般项目(13BTY049);高等学校博士学科点专项科研基金(20130002120035);清华大学自主科研青年基金(20151080433)。
于洪军(1982-),男,河北沧州人,副教授,博士,主要研究方向为运动训练理论、身体活动与健康,Tel:(010)62795363,E-mail:yuhj12@mail.tsinghua.edu.cn;李成伟(1982-),男,福建宁德人,讲师,硕士,主要研究方向为运动训练学、跳水训练理论;E-mail:lichengwei@mail.tsinghua.edu.cn;于淼(1985-),女,吉林长春人,馆员,在读博士研究生,主要研究方向为信息组织,E-mail:yumiao@ruc.edu.cn。
1.清华大学 体育部,北京 100084;2.中国人民大学 图书馆,北京 100872 1.Tsinghua University,Beijing 100084,China;2.Renmin University,Beijing 100872,China.
G808
A
