白木香结香过程中阻隔层的形成及其特性研究
2016-11-10刘培卫张玉秀刘洋洋
刘培卫 杨 云 张玉秀 杨 勇 刘洋洋
(1.中国医学科学院北京协和医学院药用植物研究所海南分所,海口 570311; 2.海南省南药资源保护与开发重点实验室,海口 570311)
白木香结香过程中阻隔层的形成及其特性研究
刘培卫1,2杨 云1,2张玉秀1,2杨 勇1,2刘洋洋1,2
(1.中国医学科学院北京协和医学院药用植物研究所海南分所,海口 570311;2.海南省南药资源保护与开发重点实验室,海口 570311)
分别采用滑走切片、组织化学染色、石蜡切片技术、扫描电镜技术和分光光度法等,对白木香通体结香过程中阻隔层的形成过程、解剖结构等特性进行了研究。结果显示:木间韧皮部内的薄壁细胞在伤害刺激下脱分化、分裂产生阻隔层;阻隔层位于沉香层和过渡层之间,其厚度变化较大,由薄壁细胞群和阻隔层木质部组成;阻隔层中含有少量的淀粉和较多的可溶性总糖。上述研究结果为消除或延迟阻隔层的产生提供参考,为进一步优化通体结香技术提供帮助。
白木香;木间韧皮部;阻隔层;通体结香技术
沉香是瑞香科(Thymelaeaceae)沉香属(AquilariaLam)植物受到伤害后形成的含有树脂的木材[1],具行气止痛、温中止呕、纳气平喘的功效[2],是我国、日本、印度以及其他东南亚国家传统的名贵药材和天然香料[3],需求量巨大。目前全世界沉香及其直接加工产品的贸易额达200亿元以上[4]。受高效结香技术的困扰,沉香产量极低,使得沉香价格一直居高不下,优质沉香价格甚至比黄金还贵[5]。过度的伐树结香,使得沉香属野生资源日渐枯竭,2004年沉香属的所有种均被列入《濒危野生动植物种国际贸易公约》附录Ⅱ[6]。白木香(A.sinensis(Lour.) Spreng)是生产国产沉香的正品植物资源[2],也是我国特有的珍贵药用植物。
项目组前期发明了世界上领先的通体结香技术,该技术具有高效—高产—高质—稳定(三高一稳定)的优点,使沉香产业进入了规模化和规范化发展的道路[7]。在通体结香技术大面积推广和使用中,我们发现部分白木香树会在沉香层的外围形成一层宽约1~5 mm的白色带状的结构,它的出现限制了沉香层的增厚,大大降低了沉香的产量和品质[5]。我们将其命名为阻隔层,并首次对其结构和组织化学特性进行详细的研究报道,旨在为消除或延迟阻隔层的产生提供参考,为进一步优化通体结香技术提供帮助。
1 材料与方法
1.1 材料
海南演丰白木香种植基地,树龄8年。选取通体结香处理30 d,60 d,180 d,1年,2年的白木香树,将样木伐倒后,在每株样木的胸径处切取一个厚约5 cm的圆盘,用于后续实验。
1.2 方法
1.2.1 组织化学染色
采用滑走切片技术,对试样进行横切,分别采用水合氯醛透明,I2-KI染色和PAS染色等处理后,在光学显微镜下(Nikon,80i,日本)进行观察和拍照。
1.2.2铬酸—硝酸法(Jeffery法)观察阻隔层细胞的形态
将分离得到的阻隔层和白木层,分割成火柴棒粗细的小木条,加入20倍量的1∶1的10%硝酸和10%铬酸的离析液,在真空泵中常温处理12 h,至完全离析,放入蒸馏水中浸泡,直到水成无色为止,用50%的酒精保存。将样品用1%番红染色4 min,做成临时装片,在光学显微镜下(Nikon,80i,日本),观察和统计分子形态。
1.2.3 石蜡切片
将含有阻隔层的新鲜材料切成0.5 cm见方的小块,FAA固定,系列酒精脱水,二甲苯透明,石蜡包埋,然后用轮转切片机(Leica RM2155,德国)切片,厚10 μm,PAS染色或番红固绿对染。
1.2.4 扫描电镜样品的制备
将含有阻隔层的新鲜样品切割成大约0.5 cm见方的木块,将横切面修成光滑的平面,立即放入FAA固定液中固定4 h,在37℃恒温干燥箱中干燥7 d至完全干燥,用导电胶将样品粘贴在样品台上,用离子溅射仪(JFC-1600,日本)喷金,然后在扫描电子显微镜(JSM6510LV,日本)下进行观察拍照。
1.2.5 淀粉含量的测定
将采集到的样品于70℃下烘干后,粉碎,保存于-20℃冰箱中备用;准确称取0.2 g粉碎样品(精确至0.000 1 g),置于具塞50 mL试管中,加入25 mL 80%乙醇—饱和氯化钠溶液,室温下超声萃取25 min后过滤,向滤渣中加入40%高氯酸溶液14 mL,室温下超声萃取30 min,加入蒸馏水10 mL继续室温超声5 min,转入100 mL容量瓶中,蒸馏水洗涤2次,合并,定容、摇匀后即成待测液(含14%的40%高氯酸溶液),静置。然后取静置后的上清液10 mL,加入1 mL碘—碘化钾溶液,以含有14%的40%高氯酸溶液的蒸馏水溶液为空白(加入1 mL碘—碘化钾溶液),使用分光光度计于580 nm处进行测定,根据标准曲线计算样品中的淀粉含量。
1.2.6 可溶性总糖含量测定
取干燥粉碎样品0.2 g,溶于6 mL 80%乙醇中,于80℃水浴锅中加热40 min,然后于4 000 r·min-1离心机中离心10 min。收集上清液,沉淀部分用80%乙醇洗涤然后以相同条件进行离心,重复2次。将上清液合并,加入活性炭后过滤,用80%乙醇定容至50 mL容量瓶中,摇匀(如无色素,可不用加入活性炭)。吸取样品提取液0.5 mL于20 mL试管中,加蒸馏水1.5 mL,向试管中加入0.5 mL蒽酮乙酸乙酯试剂和5 mL浓硫酸,充分振荡,立即将试管放入沸水浴中,逐管均准确保温1 min,取出后自然冷却至室温,以空白作参比,在630 nm波长下测定,根据标准曲线计算样品中的可溶性总糖含量。
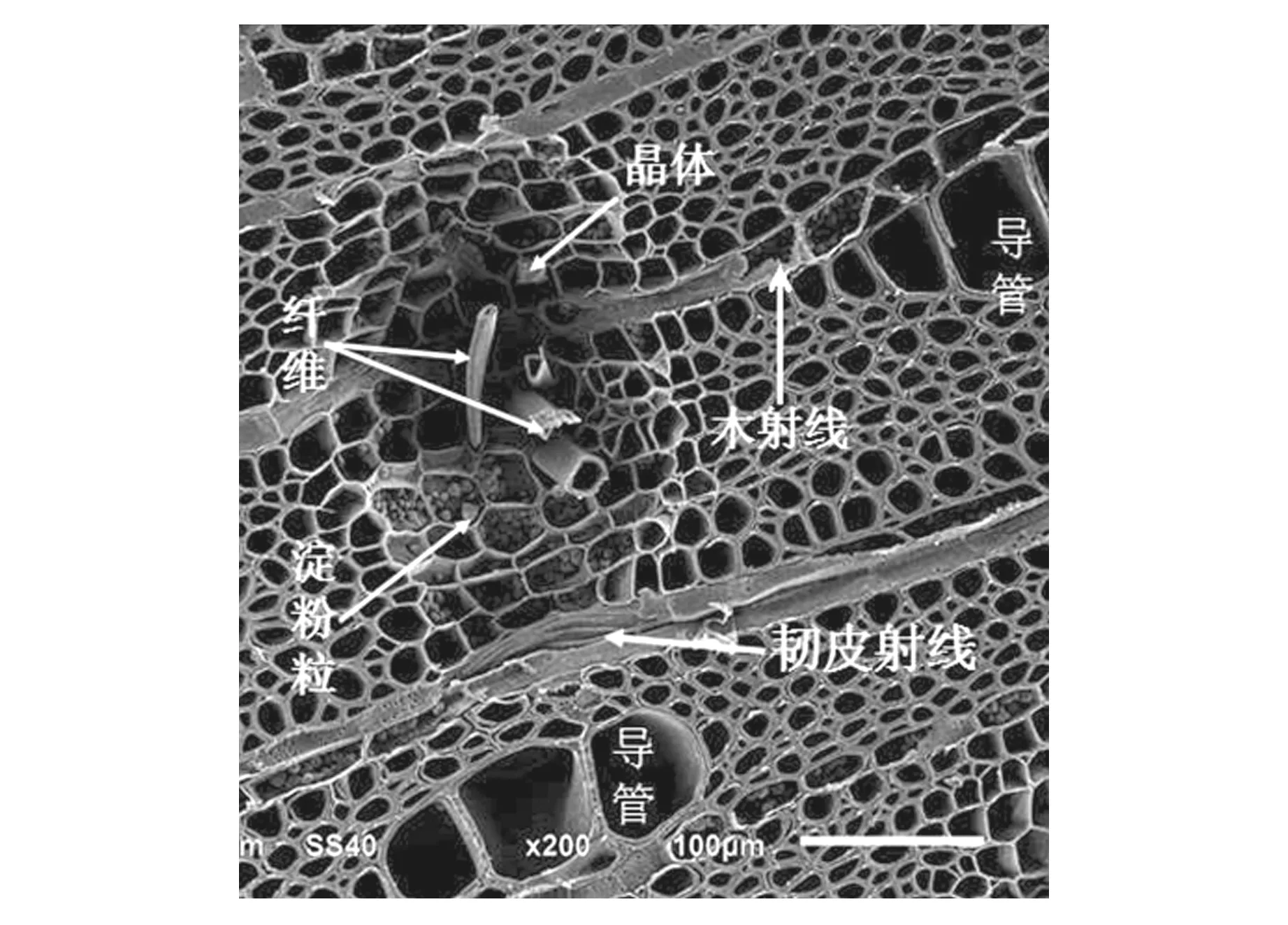
图1 成熟的木间韧皮部Fig.1 Anatomical structure of the mature interxylary phloem
2 结果
2.1 木间韧皮部的结构特征
木间韧皮部呈岛屿状,均匀散生在次生木质部中。成熟的木间韧皮部一般由薄壁细胞,筛管,韧皮射线和韧皮纤维等4部分组成(图1)。薄壁细胞位于木间韧皮部外围,通常有1~4层,近方形;韧皮射线比较扁平,与木射线相连;具有输导功能的筛管位于木间韧皮部的内部;木间韧皮部内还有一类厚壁细胞:韧皮纤维,它一般位于木间韧皮部的中下部(图1)。
2.2 阻隔层形成过程
通体结香后白木香茎在颜色和结构上会发生明显的变化,根据颜色和结构的不同,可以将其为4层:腐烂层,沉香层,过渡层和白木层(图2:a)。除了这4层外,还有一定比例的白木香树在沉香层和过渡层之间出现一层白色的带状组织:阻隔层(图2:b~e)。阻隔层的厚度不一,成连续或不连续的带状包围在沉香层周围(图2:e)。

图2 组织学水平阻隔层的形成及结构特征 a.阻隔层未出现;b.阻隔层开始出现;c~d.阻隔层逐渐增厚;e.疏松的薄璧细胞群和致密的木质部 D.腐烂层;A.沉香层;T.过渡层;B.阻隔层;H.白木层;Bp.阻隔层薄壁细胞群;Bx.阻隔层木质部 下同。Fig.2 Development and anatomical structure of barrier zone a.Barrier zone is absent; b.Barrier zone begins to form; c-d.Thickness of barrier zone increases; e.Loose Bp and compact Bx D. Decay zone;A. Agarwood zone;T. Transition zone;B. Barrier zone;H. Whiter-timber zone;Bp. Barrier zone parenchyma;Bx. Barrier zone xylem The same as below.
通过对阻隔层的跟踪观察发现,阻隔层是沉香层外围的部分木间韧皮部脱分化重新分裂形成的。在外界伤害信号的刺激下,沉香层外围的1~2层木间韧皮部薄壁细胞脱分化,进行平周分裂使木间韧皮部在径切方向上增宽(图3:b),同时在弦切方向上进行切向分裂,使木间韧皮部相互延伸,直至多个木间韧皮部连接起来,形成异常的条带状结构—阻隔层的雏形(图3:c)。随着时间的推移,在靠近沉香层的一侧,阻隔层中会形成一个比较稳定的异常形成层,它向内分裂形成薄壁纤维和薄壁细胞等,组成薄壁细胞群(图3:d),向外分裂形成异常次生木质部和异常木间韧皮部等结构,组成阻隔层木质部(图3:d~e)。阻隔层刚形成时,主要以薄壁细胞为主,但随着时间推移,阻隔层木质部所占的比例越来越大(图3:b~d)。
2.3 阻隔层的结构特征
阻隔层由阻隔层木质部和阻隔层薄壁细胞群构成,比较发达的阻隔层中,木质部所占的比例较大,其质地较致密,主要分布于靠近过渡层的一侧。阻隔层木质部与正常的木质部类似,都由木间韧皮部、木射线、导管和纤维管胞等4部分组成,但是阻隔层木质部中的木间韧皮部比较扁平,且变形比较严重,其内主要有薄壁细胞组成,没有观察到纤维(图4:c~d),阻隔层中的木射线比较臃肿,往往不与沉香层和过渡层中的木射线相连(图4:a,d),阻隔层木质部中的管孔成卵圆形或多角形,多数导管常由3~10个管孔径向连成链状结构(图4:a,d),导管分子长度约是正常导管的二分之一,宽度约为正常导管的三分之一(表1)。纤维管胞梭形(图4:a~b),长度虽然只有正常纤维管胞的三分之二,但其宽度却是正常纤维管胞的1.5倍(表1)。
阻隔层中的薄壁细胞群质地较疏松,主要位于靠近沉香层的一侧(图3:d,图4:b,d),但偶尔也有部分的薄壁细胞群散生在过渡层附近(图3:f,图4:a)。阻隔层薄壁细胞群主要由薄壁纤维和薄壁细胞组成,其中薄壁纤维的平均长度为1 574 μm(表1),其质地柔软,容易弯曲(图5:c)。阻隔层薄壁细胞群干燥后,往往收缩成蜡质的薄膜状物质,覆盖在相邻的阻隔层木质部或沉香层上,在扫描电镜观察时,此处往往成塌陷状(图4:b)。

图3 滑走切片图,示阻隔层的形成及结构特征 a.沉香层(A)外围木间韧皮部未发生变化;b.沉香层外围开始膨胀变形的木间韧皮部(B);c.膨胀的多个木间韧皮部连接起来逐渐形成阻隔层;d.阻隔层中异常形成层(V)分裂产生的薄壁细胞群(Bp)和木质部(Bx);e.阻隔层中的导管(箭头)和木间韧皮部(IP);f.阻隔层(B)与过渡层(T)的交接处的薄壁细胞群(五星) IP.木间韧皮部 下同。Fig.3 Development and anatomical structure of barrier zone by sliding a.Normal interxylary phloem beside agarwood zone(A); b.Swollen interxylary phloem(B); c.Swollen interxylary phloems begin to connect; d.Barrier zone parenchyma(Bp) and xylem(Bx) arised from abnormal cambium(V);e.Vessel(arrowhead) and interxylary phloem in barrier zone; f.Barrier zone parenchyma(asterisk) near transition zone IP. Interxylary phloem The same as below.
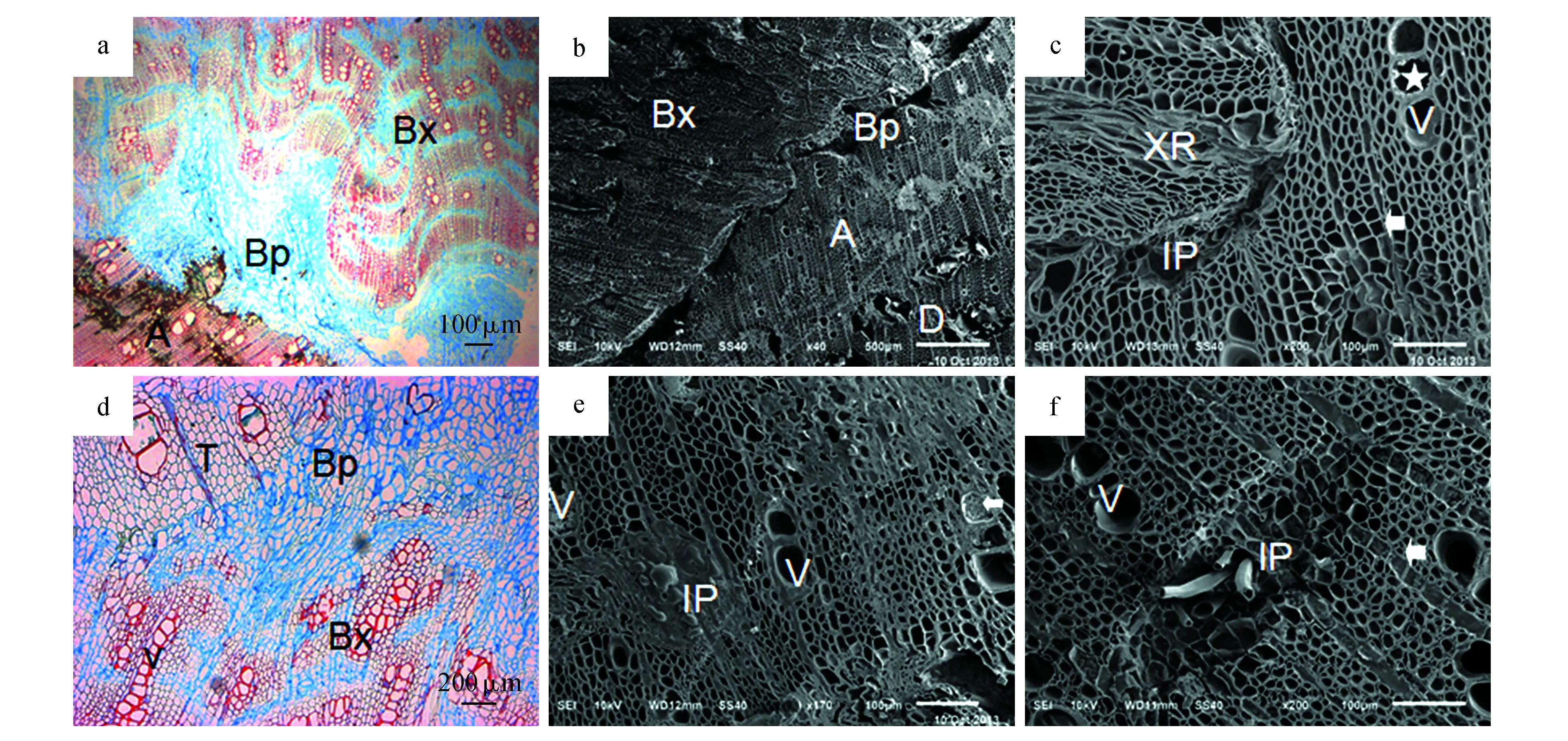
图4 石蜡切片及扫描电镜下阻隔层的显微结构特征 a.阻隔层薄壁细胞群(Bp)与木质部(Bx);b.阻隔层与沉香层交界处,塌陷状的薄壁细胞群(Bp);c.阻隔层木质部变形的木间韧皮部(IP)和异常多列射线(XR);d.阻隔层中特异的链状导管(v);e.沉香层,示充满沉香树脂的木间韧皮部(IP),含有侵填体的导管(箭头);f.白木层,示正常的木间韧皮部(IP),导管(V)和木射线(箭头)Fig.4 Microscopic picture of barrier zone by paraffin section and scanning electricity microscopy a.Abnormal parenchyma(Bp) and xylem(Bx) in barrier zone; b.Parenchyma collapses(Bp); c.Abnormal interxylary phloem(IP) and xylem ray(XR); d.Abnormal vessel(V); e.Interxylary phloem(IP) and vessel(arrowhead) containing agarwood resin in Agarwood zone; f.Normal interxylary phloem(IP), vessel and xlem ray(arrowhead) in whiter-timber zone

图5 阻隔层和白木层的离析图 a.白木层中纤维管胞;b.阻隔层木质部中纤维管胞和导管;c.阻隔层薄壁细胞群中的纤维Fig.5 Microscopic picture of segregation tissue of barrier zone and whiter-timber zone a.Fibre tracheid in whiter-timber zone; b.Fibre tracheid and vessel in Bx; c.Fiber in Bp
表1白木层和阻隔层中木材分子的数量特征(均值±标准差)
Table1Quantitycharactersofstructureoftimberaboutwhiter-timberandbarrierzone(Dataaremeans±SD)
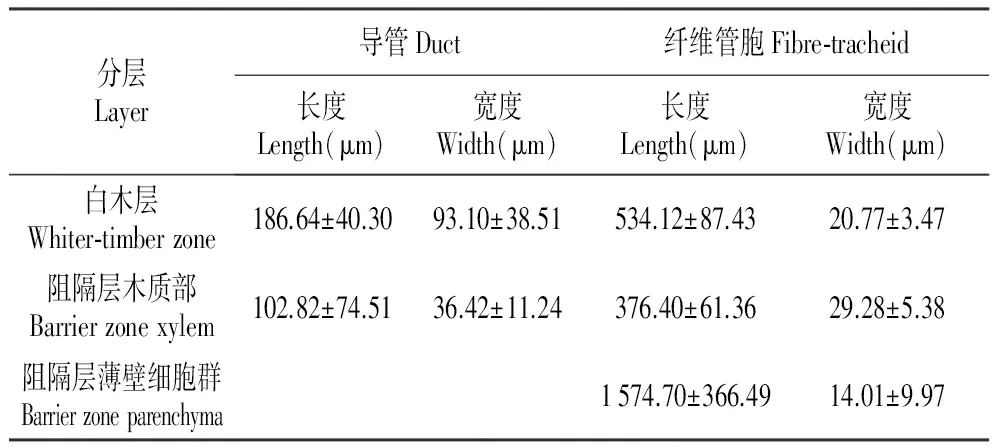
分层Layer导管Duct纤维管胞Fibre⁃tracheid长度Length(μm)宽度Width(μm)长度Length(μm)宽度Width(μm)白木层Whiter⁃timberzone186.64±40.3093.10±38.51534.12±87.4320.77±3.47阻隔层木质部Barrierzonexylem102.82±74.5136.42±11.24376.40±61.3629.28±5.38阻隔层薄壁细胞群Barrierzoneparenchyma1574.70±366.4914.01±9.97
2.4 淀粉和可溶性总糖的含量测定
碘—碘化钾(I2-KI)组织化学染色结果显示,沉香层中淀粉粒被完全消耗(图6:c),阻隔层中还存在少量的淀粉粒(图6:b),而白木层中依然存在大量的淀粉粒(图6:a)。希夫试剂染色(PAS反应)结果显示,被油脂完全填充的沉香层无红色反应(图6:f),阻隔层(图6:e)中和白木层(图6:d)中都有较深的红色反应(图6:e),说明沉香层中的还原糖已经被完全消耗,阻隔层和白木层中含有大量的还原糖。

图6 组织化学染色显微图 a~c. I2-KI染色:a.白木层,IP内含有大量的淀粉;b.阻隔层,IP含有少量的淀粉;c.沉香层,IP中不含有淀粉 d~f. PAS染色:d.白木层,IP中含有大量还原糖;e.阻隔层,IP含有大量的还原糖;f.沉香层,IP中充满大量棕色树脂Fig.6 Microscopic picture of histochemical staining a-c. I2-KI staining: a. Much starch in interxyalry phloem in whiter-timber zone; b. Less starch in barrier zone; c.No starch and reducing sugar in agarwood zone 4-6. PAs staining: d. Much reducing sugar in whiter-timber zone; e. Much reducing sugar in barrier zone; f. No starch and reducing sugar in agarwood zone
为了更精确的观察淀粉和可溶性总糖的含量,我们利用分光光度法,对白木香茎中不同层内的淀粉和可溶性总糖进行定量检测。结果发现,沉香层中可溶性总糖和淀粉含量均最低,说明沉香层中的能量物质大多转化为沉香树脂。可溶性总糖含量在阻隔层、过渡层和白木层中差别不大,但淀粉含量差异明显。各层中可溶性总糖和淀粉含量的比值从大到小依次为:阻隔层(1.45)>白木层5(0.87)>过渡层(0.60)>沉香层(0.29)>白木层4(0.32)。木间韧皮部分裂形成阻隔层,需要较大的能量代谢为旺盛的生理活动提供动力。而淀粉被转化为可溶性糖类物质才能被利用,因此可溶性总糖和淀粉的比值越高,说明代谢活动越旺盛。
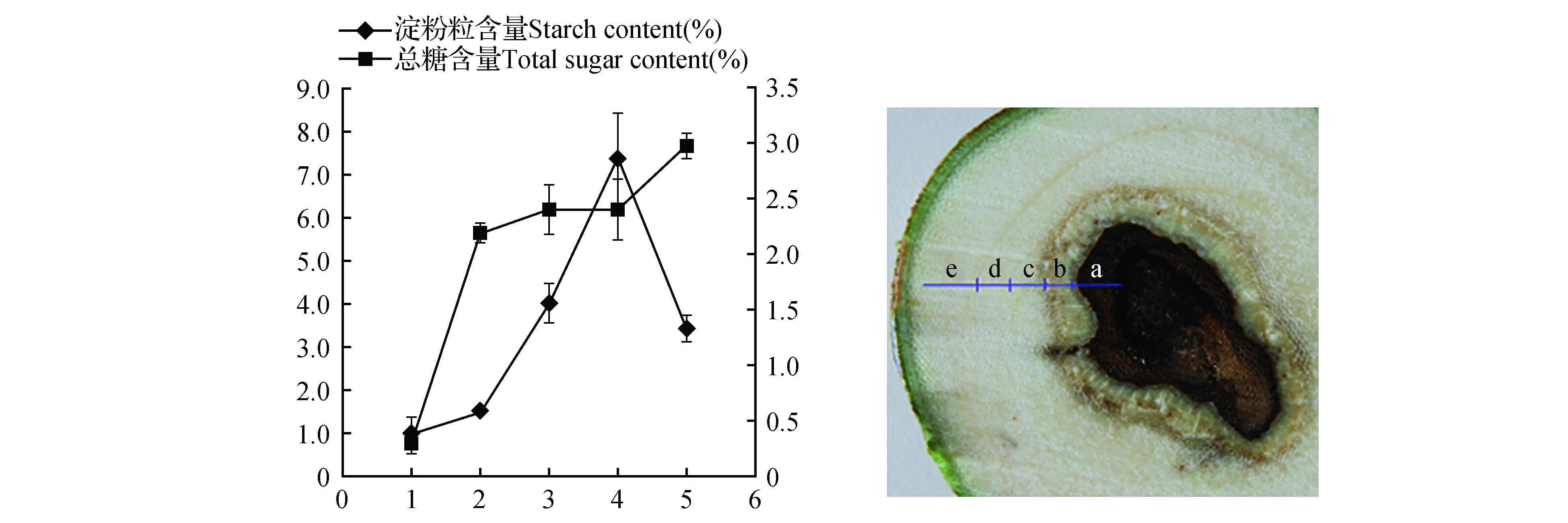
图7 不同分层中淀粉和可溶性总糖的含量 a.沉香层;b.阻隔层;c.过渡层;d~e.白木层Fig.7 Content of starch and total sugar in different zones a.Agarwood zone;b.Barrier zone;c.Transition zone;d-e.Whiter-timber zone
3 讨论
3.1 木间韧皮部的作用
笔者在前人研究的基础上,发现至少在17个科约54个属的部分或全部物种中具有木间韧皮部[8],具有木间韧皮部的植物往往具有巨大的果实或花朵[9]。我们根据木间韧皮部的解剖结构推测木间韧皮部在光合产物的运输和贮藏等方面具有重要的作用。人们发现剥除树皮后的白木香树仍旧可以存活,这有力的验证了我们的推断。
有学者早在1992年发现,沉香属植物木间韧皮部内部的薄壁细胞可能保持有或者在外界刺激的激发下重新获得细胞分裂的能力[10]。笔者在研究阻隔层形成的过程中,发现木间韧皮部内的1~2层薄壁细胞脱分化重新恢复分裂功能,使木间韧皮部在径切方向上增宽、弦切方向上延伸,多个木间韧皮部逐渐连接起来,在沉香层外围形成异常的条带状结构—阻隔层。我们的发现证明了木间韧皮部确实具有重新分裂的能力,但必须在外界刺激下才能发生,因此我们将沉香属木间韧皮部称为“休眠的形成层”。
3.2树木区室化在伤害诱导白木香形成沉香中的意义
众所周知,树木不能像动物那样来躲避伤害,但在亿万年的进化过程中,树木为了生存,掌握了一种独特的、高超的抵御伤害的能力—区室化[11]。区室化是指树木受到伤害后,通过形成多种屏障将受到伤害的组织隔离开来,以阻止病原物等伤害的扩散[12]。树木区室化主要分为2个阶段[12]:(1)在伤害或者病原物侵染的过程中,伤口周围的活细胞产生抗菌物质,从而形成化学边界。(2)形成层在伤害信号因子的刺激下,形成化学成分和结构都不同的特殊组织,称之为阻隔层。阻隔层将健康组织和受到伤害的组织隔离开,可有效的阻止伤害在树体内扩展。Blanchette等明确提出树木受到创伤后,木材的变色、死亡和腐烂,都属于区室化现象[13]。
沉香是沉香属植物受到伤害后形成的含有树脂的木材。项目组初步解析了“伤害诱导白木香防御反应形成沉香”理论:物理、化学伤害或真菌侵染均可作为伤害诱导白木香产生防御反应,产生具有抑菌活性的防御物质—沉香倍半萜和色酮类成分,这些防御物质填充在木间韧皮部、木射线、导管和纤维管胞内,与周围的木材结合在一起,形成了阻挡伤害进一步扩散的屏障[14]。该理论与树木区室化理论的第一阶段非常吻合,即形成了抵御外界伤害的化学边界—沉香层。根据区室化理论,阻隔层形成是树木区室化的第二阶段,是树木响应伤害信号后,产生的具有保护功能的重要屏障。
从位置上来看,阻隔层位于沉香层和过渡层之间,它的出现使健康组织远离了伤害;从结构上来看,阻隔层薄壁细胞中的纤维长度与树皮中的纤维长度相近,而且韧性好,所以我们通常把阻隔层称为“新树皮”。Rioux等认为阻隔层的细胞中往往含有抗菌活性的酚类物质,细胞壁往往栓质化,可以有效的防止水分流失[15]。总之,阻隔层在位置、结构和化学组成上,都成了阻挡伤害向树体正常组织扩展的重要屏障,对白木香抵御外界伤害意义重大,这也从理论上证明了“伤害诱导白木香防御反应形成沉香”理论的科学性。
3.3 本研究对于优化结香技术具有重要的意义
阻隔层的形成与树的品种,个体遗传学,伤害类型以及环境因素有关。但树木自身情况是影响阻隔层形成的关键因素[16]。我们发现3年生的白木香在通体结香后的30天,可以观察到阻隔层的出现,但在10年以上的老树上,一般1年以后才开始大量出现阻隔层。Rioux等认为幼树与老树相比,储存着较少的能量物质[15]。3年生的白木香幼树存储的营养物质偏低,当幼树受到伤害后,化学边界(沉香层)的形成很快将存储的物质消耗殆尽,其阻隔作用较弱;伤害信号通过沉香层进入正常组织中,激发临近木间韧皮部的薄壁细胞脱分化恢复分裂形成第二层屏障,即阻隔层,以有效的抵御外界伤害在树体内扩展。而10年以上的老树正当壮年,积累了大量的营养物质,受到伤害后可以形成较厚的沉香层,能够在一定时期内阻挡伤害的进一步扩展。通过可溶性总糖和淀粉含量的研究,我们发现沉香层中几乎不含有可溶性总糖和淀粉,说明储藏的能量物质几乎全部转化为沉香树脂;阻隔层由于需要建成新的细胞所以代谢活动较旺盛,其可溶性总糖含量和淀粉含量的比值最高;而过渡层介于两者中间。这更加验证了我们的推测。从这方面来看,老树才能结香的说法有一定的科学道理。
我们在通体结香的大面积推广中发现,随着结香时间延长,阻隔层出现的概率有增大的趋势。但这种趋势在不同的地域中有所差异,在贫瘠的条件下,材质比较坚硬的树木,阻隔层出现的时间较晚,且概率小。在肥沃的条件下,材质比较疏松,阻隔层出现的时间较早,概率较大。Rioux等将长喙壳属真菌(Ophiostomaulmi)侵染PrunuspensylvanicaL.发现,侵染4天时阻隔层首次出现,侵染15天后出现的概率几乎为100%[15]。因此伤害诱导结香的时间并不是越久越好,根据树木材质以及营养环境的不同,选择一个适宜的结香时间以及收获时间才更能保证沉香的产量和品质。
3.4 小结
阻隔层是白木香抵御伤害的重要屏障,但是它的出现限制了沉香层的加厚,严重影响了沉香的产量和品质。如何消除和减少阻隔层的产生,成为研发更为高效结香技术的瓶颈。本研究系统研究了阻隔层的形成过程和组织化学特性,同时探讨了木间韧皮部在阻隔层形成中的作用及影响阻隔层形成的因素,这对研究如何消除或减少阻隔层的形成、适宜结香树龄的筛选,以及结香技术的优化升级奠定了一定的理论基础,对提高沉香的产量和品质具有很好的价值和意义。
1.Liu Y Y,Chen H Q,Yang Y,et al.Whole-tree agarwood-inducing technique:an efficient novel technique for producing high-quality agarwood in cultivatedAquilariasinensistrees[J].Molecules,2013,18(3):3086-3106.
2.国家药典委员会.中华人民共和国药典2015年版(一部)[K].北京:中国医药科技出版社,2015:185.
3.Jung D.The cultural biography of agarwood-perfumery in eastern Asia and the Asian neighbourhood[J].Journal of the Royal Asiatic Society,2013,23(1):103-125.
4.吕芳华,高维新.沉香贸易现状及发展前景研究[J].现代商贸工业,2013,25(6):65-66.
Lü F H,Gao W X.Trade situation and development prospect onAquilariaagallochaRoxb.[J].Modern Business Trade Industry,2013(06):65-66.
5.张兴丽.伤害诱导的白木香防御反应与沉香形成的关系研究[D].北京:北京林业大学,2013.
Zhang X L.Studies on Relationships between Wound-induced Defense response and Agarwood formation inAquilariasinensis[D].Beijing:Beijing Forestry University,2013.
6.CITES.Amendments to appendices Ⅰ,Ⅱ and Ⅲ of cites[C].//Proceedings of the thirteenth meeting of the conference of the parties.Bangkok,Thailand,2004.
7.黄俊卿,魏建和,张争,等.沉香结香方法的历史记载、现代研究及通体结香技术[J].中国中药杂志,2013,38(3):302-306.
Huang J Q,Wei J H,Zhang Z,et al.Historical records and modern studies on agarwood production method and overall agarwood production method[J].China Journal of Chinese Materia Medica,2013,38(3):302-306.
8.张玉秀,刘培卫.木间韧皮部的定义、分布、发育和生理功能[J].湖北农业科学,2015,54(15):3589-3592.
Zhang Y X,Liu P W.Intexylary phloem:definition,distribution,development and function[J].Hubei Agricultural Sciences,2015,54(15):3589-3592.
9.Carlquist S.Interxylary phloem:diversity and functions[J].Brittonia,2013,65(4):477-495.
10.Rao K R,Dayal R.The secondary xylem ofAquilariaagallocha(Thymelaeaceae) and the formation of ‘agar’[J].IAWA Journal,1992,13(2):163-172.
11.Shigo A L.Compartmentalization of decay in trees[J].Scientific American,1985,252(4):96-103.
12.Shigo A L.Compartmentalization:a conceptual framework for understanding how trees grow and defend themselves[J].Annual Review of Phytopathology,1984,22:189-214.
13.Blanchette R A,Biggs A R.Defense mechanisms of woody plants against fungi[M].Berlin Heidelberg:Springer,1992:1-12.
14.张争,杨云,魏建和,等.白木香结香机制研究进展及其防御反应诱导结香假说[J].中草药,2010,41(1):156-159.
Zhang Z,Yang Y,Wei J H,et al.Advances in studies on mechanism of agarwood formation inAquilariasinensisand its hypothesis of agarwood formation induced by defense response[J].Chinese Traditional and Herbal Drugs,2010,41(1):156-159.
15.Rioux D,Ouellette G B.Barrier zone formation in host and nonhost trees inoculated with Ophiostoma ulmi.Ⅱ.Ultrastructure[J].Canadian Journal of Botany,1991,69(9):2074-2083.
16.Smith K T.Compartmentalization today[J].Arboricultural Journal:The International Journal of Urban Forestry,2006,29(3):173-184.
Natural Science Foundation of Hainan Province(314183);PUMC Youth Fund and the Fundamental Research Funds for the Central Universities(33320140078);Traditional Chinese Medicine Industry Special Project(201407005);Modernization of Traditional Chinese Medicine of Hainan Province(2015ZY13)
introduction:LIU Pei-Wei(1983—),male,research assistant,research on anatomy and active ingredients of medicinal plant.
date:2016-06-02
BarrierZoneFormationinAgarwoodFormationofAquilariasinensis
LIU Pei-Wei1,2YANG Yun1,2ZHANG Yu-Xiu1,2YANG Yong1,2LIU Yang-Yang1,2
(1.Institute of Medicinal Plant Development,Chinese Academy of Medical Sciences & Peking Union Medical College,Haikou 570311;2.Hainan Provincial Key Laboratory of Resources Conservation and Development of Southern Medicine,Hainan Branch,Haikou 570311)
With the sliding section, histochemical staining, paraffin section, scanning electricity microscope and spectrophotometry, we studied the anatomical structure and histochemcial characteristics of barrier zone. The barrier zone was arisen from interxylary phloem. Barrier zone in different thickness located between agarwood zone and transition zone. The barrier zone was formed of abnormal parenchyma and xylem. Barrier zone contained a small amount of starch and more total sugar. Our results might be used to eliminate and delay the barrier zone, and improve whole-tree agarwood-inducing technique.
Aquilariasinensis;interxylary phloem;barrier zone;whole-tree agarwood-inducing technique
海南省自然科学基金(314183);北京协和医学院协和青年科研基金(33320140078);中医药行业科研专项(201407005);海南省中药现代化项目(2015ZY13)
刘培卫(1983—),男,助理研究员,主要从事南药植物结构和活性成分研究。
2016-06-02
R282
A
10.7525/j.issn.1673-5102.2016.05.010
