连香树叶片发育与生态适应研究
2016-11-10马永红
马永红
(1.西南野生动植物资源保护教育部重点实验室,南充 637009; 2.西华师范大学,南充 637009)
Author introduction:MA Yong-Hong(1977—),male,Associate professor,mainly studied the systematic and evolutionary of botany and wild plant resources.
连香树叶片发育与生态适应研究
马永红1,2
(1.西南野生动植物资源保护教育部重点实验室,南充 637009;2.西华师范大学,南充 637009)
用石蜡切片法对连香树(Cercidiphyllumjaponicum)叶片进行解剖学观察和研究,并从叶片不同发育阶段的形态变化、解剖结构差异等方面,探讨了其生态适应性。获得如下主要结果:(1)形态方面,叶片发育呈现边缘内卷期、幼叶开展期和生长成熟期三个时期。第1、2对幼叶的叶柄基部具发达的膜质叶鞘,叶鞘的发育呈现出:第一对幼叶的叶鞘发达、第二对较弱、其它幼叶无叶鞘;(2)解剖结构显示:连香树叶为典型异面叶、海绵组织发达;上下表皮均具较厚的角质层和气孔器、气孔外凸、叶片内机械组织和叶脉较发达、叶柄中段具发达的周韧维管束且有维管束鞘,显示连香树对干旱和阴湿环境均具较强的生态适应能力。(3)营养器官的基本组织细胞中存储有大量后含物,可能为繁殖时期器官的构建提供主要营养物质,而顶芽产生的第一对叶,仅对早春其它器官发育提供一定的物质补充。
连香树;叶片解剖;发育;生态适应
叶的发育是植物形态建成的一个重要方面[1],在不同发育阶段其形态结构可能完全不同、与之相关的生理功能和对生态环境的适应能力都存在显著差异。另外,该器官暴露面大,与其它器官相比受环境影响最大,最能反映植物对生态环境适应的特点[2]。连香树(Cercidiphyllumjaponicum)是连香树科(Cercidiphyllaceae)连香树属(Cercidiphyllum)植物。该科植物作为一种先锋树种,在晚白噩纪和早第三纪北半球中等纬度地区,曾广泛分布于北美、欧洲和亚洲[3~7],现仅存连香树1种,为第三纪孑遗的单型科植物,且仅间断分布在中国和日本[8],现为国家Ⅱ级濒危保护植物。该物种在我国,目前主要分布于鄂、湘、赣、川、甘、晋、陕、皖、豫、浙等多省,但种群斑块化明显,且种群规模极为有限[9]。四川为连香树的主要分布中心,常见于盆地周边海拔1 000~2 000 m的山区。
有关连香树营养器官的解剖研究,国内仅见黎明和王东等[10]曾对叶柄维管束的变化和次生木质部导管的形态特征进行研究[11~12]。而叶片发育方面,仅见闫小玲[13],分析了该物种的系统演化地位,而基于连香树营养器官不同发育阶段的营养能量分配、对环境要素的差别要求及结构变化对生长环境的适应等鲜有研究报道。笔者通过对连香树不同发育阶段的叶片形态和解剖结构进行观察,对其相应的生态适应性进行初步分析,以期为该物种的生态保护、濒危机制研究提供依据和思路。
1 材料与方法
1.1 材料
幼叶及芽于2009年4月4日采自四川省峨眉山九老洞,海拔1 700 m。将新鲜幼叶及芽置于FAA(50%乙醇90 mL∶冰醋酸5 mL∶甲醛5 mL)中固定24 h,再转入到70%的酒精溶液保存,置于4℃冰箱中保存待用;成熟叶片取自蜡叶标本,采集于甘洛县马鞍山自然保护区海拔1 650 m的山坡疏林中。
1.2 方法
选择不同发育阶段叶近中部靠近中脉处2~3块(0.5 cm×0.5 cm)及其叶柄(0.5~1.0 cm);成熟叶片经10%的氢氟酸软化24 h,流水冲洗48 h。采用常规石蜡切片法,切片厚度8 μm;番红—固绿对染;中性树胶封片。叶表皮研究方法:取成熟叶片主叶脉中部,剪成约0.5 cm2的小片,煮沸排气3~5 min后,置于解离液(10%的铬酸水溶液100 mL,加浓硝酸5 mL)中3~5 h。用毛笔刷去残余叶肉,冲洗干净后用1%的苏木精溶液染色20 min,自来水反色,梯度脱水,中性树胶封片。MoticBA300型光学显微镜及成像系统下观察、拍照。
2 结果与分析
2.1 叶片的形态发育
连香树叶片形态发育可分为边缘内卷期、幼叶开展期和生长成熟期三个时期。其中第一对幼叶边缘内卷期自3月中旬开始至3月底(图版Ⅰ:3)。在该时期,每枚幼叶的叶柄基部已分化形成1枚发达的膜质叶鞘(图版Ⅰ:1~2),叶鞘卵圆形,边缘内卷,仅基部与叶柄基部愈合。其它幼叶还处于发育的早期。3月底叶片发育进入开展期,4月初第一对幼叶基本完全展开,叶片呈暗红色,随后便逐渐发育成熟,颜色逐渐转为绿色。刚开展的幼叶大小平均值达到1.87 cm×1.92 cm(长×宽),叶形呈卵形、近圆形或阔卵形,叶缘具钝锯齿,基出5~7脉明显,并在叶片远轴面显著凸起,形态与成熟叶片基本一致。该时期,其第二、三对幼叶正处于边缘内卷期。有趣的是第二对幼叶也存在膜质的叶鞘,位于叶柄的近轴面,几乎与叶柄同宽,但较第一对叶的叶鞘明显弱小,横切面呈“【”形(图版Ⅱ:3~6),而其他幼叶均无明显的叶鞘结构。连香树叶鞘发育呈现出的第一对幼叶的叶鞘发达→第二对弱小→其它幼叶未见的特征与该物种雄花序外层→中层→内层的雄花花被片发育所呈现出的1枚→1枚简化→无的规律基本一致[12]。成熟叶片并未发现膜质叶鞘的结构,说明在叶片在成熟过程中,叶鞘最终脱落。
从幼叶的发生来看:由长枝顶端的一对顶芽分化出第一对幼叶时,每芽最先只分化出1枚叶,并非成对产生,所以早春时期所产生的第一对叶可以认为是假对生,可能原因是连香树冬芽在萌发时,其生长锥在形成第一对叶原基后,只有远轴面的叶原基快速发育形成幼叶;或者生长锥最先只在远轴面产生一个叶原基并发育成第一枚叶。其它幼叶则成对产生于由顶芽生长锥发育而来的幼茎上。从生长速度方面,第一对叶明显生长较快。叶鞘的发育和第一对叶片的产生及发育过程等,至今还未见相关文献报道。
2.2 解剖结构观察
2.2.1 叶表皮发育
连香树幼叶的叶片上、下表皮均为一层细胞组成,细胞横切面呈长方形,排列紧密,两面光滑(图版Ⅰ:3~4)。成熟叶片的厚度约为179.52~182.21 μm,其中上表皮厚度为18.48 μm,约占叶片厚度的10%,细胞多长方形,叶脉处略呈柱状椭圆形,细胞垂周壁多平直(图版Ⅰ:5)。下表皮细胞较小,横切面呈不规则形,细胞垂周壁多弯曲。表皮厚度为8.68 μm,占叶片厚度的4.78%。成熟叶片表皮微形态特征显示:上表皮细胞呈不规则的多边形至扁圆形,垂周壁平直或弯曲或浅波状;下表皮细胞不规则形,垂周壁多呈浅波状。气孔器椭圆形,由两个保卫细胞组成,无副卫细胞,其长短轴比值的平均值为1.47。气孔器主要分布叶片下表皮,无规则型分布,气孔平均密度为0.165。气孔器在上表皮偶见,分布高度不均匀,平均密度仅约为0.048(图版Ⅰ:6~7)。
2.2.2 叶肉细胞的发育与分化
幼叶在边缘内卷期(发育早期),结构上除了主脉处出现微弱的基本组织和初生维管组织的分化外,叶肉还未明显分化。细胞横切面多呈等径的四边形,核大,细胞质较浓。但在该发育阶段,位于幼叶边缘的齿凹处的球形排水器十分发达。幼叶发育至平展时,其叶肉细胞开始分化。初步分化的栅栏组织位于上表皮下方,由3层经过径向伸长生长短柱状细胞组成(图版Ⅰ:3~4)。成熟后的叶片,栅栏组织细胞的层数和细胞形态与分化初期变化不大,但细胞壁较幼期厚,细胞径向有一定伸长。栅栏组织平均厚度为58.48 μm,约占成熟叶片厚度的32.5%(图版Ⅰ:5)。
海绵组织和栅栏组织为同步分化。在幼叶边缘内卷期,其横切面中部的细胞的核更大,细胞质更浓,显示出更为旺盛的分裂和代谢能力(图版Ⅰ:3~4)。随叶片的生长发育,由该部分细胞衍生出来的细胞经过生长、分化,细胞形态趋于不规则形,并出现了明显的细胞间隙,该区域的大部分细胞最终发育形成海绵组织。成熟叶片的海绵组织发达,平均厚度为101.76 μm,约占叶片厚度的56.53%。细胞形状多样,且细胞排列疏松,间隙发达(图版Ⅰ:5)。
2.2.3 叶脉的发育与类型
分化早期,主脉中心部分产生不连续的原形成层环,但厚度不均匀,远轴面厚度为近轴面的2~3倍。靠近远轴面的原形成层细胞经过平周分裂,向内衍生出的细胞经过生长分化,发育成叶脉较发达的木质部,向外衍生出的细胞经过生长分化,发育成叶脉较发达的韧皮部。近轴面的原形成层经过分裂和分化,形成基本组织细胞和少量的厚角组织细胞。幼叶维管束与上、下表皮之间的厚角组织发达,主脉和侧脉在叶片两面均显著凸起(图版Ⅰ:8);随着幼叶的进一步发育至成熟,叶片厚度增加,远轴面的皮层细胞体积明显增大,另外由于维管组织靠近远轴面,使得叶片的主脉在远轴面显著凸起,而在近轴面轻微突出。成熟后的叶脉仅具有一个“U”形的外韧维管束(图版Ⅱ:1~2)。维管束的宽度占整个主脉宽度的68.75%,厚度占整个主脉厚度的34.78%。黎明等发现连香树成熟主脉的维管束外具有厚壁纤维组成的维管束鞘[9],笔者的在本次研究中并未发现显著的维管束鞘存在,这可能与生长环境或叶片的成熟度有关。
2.2.4 叶柄的发育
在幼叶边缘内卷期,叶柄表皮仅一层细胞,由原表皮组成,细胞横切面呈等径的四边形或近圆形,细胞核大而显著,细胞质浓。皮层细胞由两层薄壁组织细胞组成,细胞排列紧密,细胞形态多样,细胞核明显,但细胞质较稀疏。皮层内则为厚度不均匀的原形成层环,细胞呈等径的圆形至多边形,细胞质浓厚,分裂活跃。原形成层环的中央由数个较大的薄壁组织细胞构成中央髓部。幼叶发育进入平展期以后,原形成层最初衍生出的细胞经过生长、分化形成皮层薄壁组织细胞,使皮层细胞由原来的2层发育至7~8层,其中少部分皮层薄壁组织细胞发育成厚角组织,原形成层衍生出部分细胞再分化形成维管组织。靠近远轴面的木质部主要由5~7层导管分子组成,细胞排列密集整齐且管径大小几乎一致,内始式发育;组成韧皮部的细胞层数与木质部相近,但细胞小,故韧皮部的平均厚度仅约为木质部平均厚度的1/4。近轴面的维管组织的厚度与远轴面相当,但分布不连续,显示出原形成层在产生维管组织的过程中,远轴面的细胞分裂和分化速度显著快于近轴面,无明显的维管柱鞘(图版Ⅱ:3~7)。
成熟叶柄中段的横切面呈圆形,具不规则且深浅不一的纵棱槽。其基本结构包括表皮、皮层和维管柱三部分。表皮由一层细胞构成,与幼期相比,成熟期的表皮细胞横切面观呈椭圆形或梭形,细胞外切向壁较厚。皮层由7~8层细胞组成,且多数细胞为厚角组织细胞,这些厚角组织细胞均由皮层薄壁组织细胞的初生壁角隅处加厚发育而成。叶柄的疏导组织仅具有一个周韧维管,且维管束外围由2~3层次生壁强烈加厚的纤维构成了发达的维管束鞘;与幼期相比,成熟叶柄的维管束在远、近轴面的厚度相当(图版Ⅱ:8)。
3 讨论
3.1 繁殖期的营养供给与生态适应
黎明等对连香树营养器官的解剖研究中发现,不论是在叶柄还是茎髓部的基本组织细胞中,均有大量的颗粒状贮藏物质[10];郭瑛对连香树叶片的叶绿素含量变化进行测定,发现展叶后的第19天,叶绿素含量才达最大值[14]。而连香树为先花后叶或花叶同放植物,说明在繁殖早期,由叶片合成的光合产物并不能完全满足繁殖器官和幼叶构建的需要。笔者认为连香树繁殖前期所需的大量营养物质和能量供给可能大部分由其它营养器官的基本组织细胞所贮藏的后含物提供,而第一对叶的快速生长为繁殖前期提供了一定的物质与能量补充,该物种次年能否正常开花繁殖与其上年生长环境、物质积累和植株健康状况密切相关。
3.2 叶鞘发育与生态适应
笔者首次观察到第一、二对幼叶具发达的叶鞘,且第一枚叶叶柄基部膜质叶鞘包被着当年还未完全伸展的枝芽。虽然连香树的冬芽由两枚褐色薄革质的芽鳞以套合的方式包被,但当第一对幼叶从芽中伸出时,芽鳞已无法满足对生长缓慢的芽尖生长锥进行有效保护,故第一对叶片叶柄基部发达的膜质叶鞘则暂时替代了芽鳞的作用,一定程度上增强了幼嫩器官适应早春环境剧烈变化的能力。
3.3 叶片解剖特征及生态适应
连香树叶片横切特征显示:气孔器多分布于下表皮,且微外凸;叶脉较发达,机械组织细胞较多,叶片较厚,这与黎明[10]和闫小玲[13]的研究结果基本一致;结合闫小玲在扫描电镜下的观察发现,其叶片上、下表皮外覆盖的角质层较厚等特征[13],显示连香树具有一定的耐旱适应性。但该物种表皮细胞只有一层,栅栏组织不发达且细胞排列较稀疏;海绵组织发达且具发达的细胞间隙等特征显示,该物种同时也适应阴湿环境。解剖特征表明,在该物种在其自然分布区内具有较广的生态幅,这与该物种既可以在较为干旱的阳坡生长,也常见于沟谷阴湿环境的野外分布基本一致。
3.4 维管束的变化与生态适应
王东和闫小玲研究发现,连香树叶片主脉由一个外韧维管束组成,而叶柄的维管束由叶柄基部至叶片基部呈现出分离—联合—分离的有规律变化[12~13]。维管束在叶柄两端以分离的形式存在是必然的,而在叶柄中段联合的现象在其他物种中并不常见。笔者通对该物种的幼叶、成熟叶的解剖结构观察结果与前者的研究结果基本一致。不同的是:(1)成熟叶片主脉具微弱的束中形成层,这可能与叶片的年龄、该物种的分布环境有关。(2)成熟叶柄中段只具一个发达的中央周韧维管束且维管束鞘发达,该结构在一定程度上有效增强叶柄的机械强度,表现出一定的抗旱、抗风等特点,同时也有利于增强物质和水分的输送能力,来满足早春幼嫩器官构建和发育对营养物质的需求。
4 结论
植物体任何结构的产生、强化与退化消失,都将揭示相应功能的增强与减弱,这与植物个体在生长发育过程中对生活环境的适应具有密切联系。对连香树的相关研究,多数学者仅关注了成熟个体或种群,很少系统关注在个体发育的整个过程中,不同发育阶段的营养能量分配、对环境要素的差别要求、结构变化对环境的适应、雌雄个体营养器官发育的差异等。或许这些方面是研究该物种濒危机制的另一思路,不过对于这些问题还需要持续的深入研究。
1.严松,严长杰,顾铭洪.植物叶发育的分子机理[J].遗传,2008,30(9):1127-1135.
Yan Song,Yan Changjie,Gu Minghong.Molecular mechanism of leaf development[J].Herditas,2008,30(9):1127-1135.
2.王勋陵,马骥.从旱生植物叶结构探讨其生态适应的多样性[J].生态学报,1999,19(6):787-792.
Wang Xunling,Ma Ji.A study on leaf-structure and the diversity of xerophytes ecology adaptation[J].Acta Ecologica Sinica,1999,19(6):787-792.
3.Manchester S R.Biogeographical relationships of North American Tertiary floras[J].Annals of the Missouri Botanical Garden,1999,86(2):472-522.
4.Crane P B.A re-evaluation of Cercidiphyllum-like Plant fossils from the British early Tertiary[J].Botanical Journal of the Linnean Society,1984,89(3):199-230.
5.Crane P R,Stockey R A.Morphology and development of Pistillate inflorescences in extant and fossil Cercidiphyllaceae[J].Annals of the Missouri Botanical Garden,1986,73(2):382-399.
6.Crane P R,Stockey R A.Growth and reproductive biology ofJoffreaspeirsiigen.et sp.nov.,aCercidiphyllum-like Plant from the Late Paleocene of Alberta,Canada[J].Canadian Journal of Botany,1985,63(2):340-364.
7.Jähnichen H,Mai D H,Walther H.Blätter und Früchte vonCercidiphyllumSiebold & Zuccarini im mitteleuropäischen Tertiär[J].Schriftenreihe für geologische Wissenschaften,1980,12(16):357-399.
8.周浙昆,Momohara A.一些东亚特有种子植物的化石历史及其植物地理学意义[J].云南植物研究,2005,27(5):449-470.
Zhou Zhekun,Arata Momohara.Fossil History of Some Endemic Seed Plants of East Asia and Its Phytogeographical Significance[J].Acta Botanica Yunnanica,2005,27 (5):449-470.
9.李文良,张小平,郝朝运,等.珍稀植物连香树(Cercidiphyllumjaponicum)的种子萌发特性[J].生态学报,2008,28(11):5445-5453.
Li Wenliang,Zhang Xiaoping,Hao Chaoyun,et al.Seed germination characteristics of rare plantsCercidiphyllumjaponicum[J].Chinese Journal of Applied Ecology,2008,28(11):5445-5453.
10.黎明,段增强,李莲枝,等.连香树营养器官的解剖学研究[J].河南农业大学学报,2005,39(2):178-181.
Li Ming,Duan Zengqiang,Li Lianzhi,et al.Anatomical Study on the Nutritive Organs ofCercidiphyllumjaponicum[J]. Journal of Henan Agricultural University,2005,39(2):178-181.
11.王东,高淑贞.中国连香树科的系统研究—Ⅱ.次生木质部的显微和超微结构[J].西北植物学报,1991,11(4):287-290.
Wang Dong,Gao Shuzhen.Study on The Relationships of CercidiphyllaceaeⅡ.Anatomy and MorpboIogy Study on Secondary Xylem[J]. Acta Botanica Boreali-Occidentalia Sinica, 1991,11(4):287-290.
12.王东,高淑贞.中国连香树科的系统研究—Ⅰ.叶的宏观结构及叶柄维管束变化[J].西北植物学报,1990,10(1):37-42.
Wang Dong,Gao Shuzhen.A Study On The Relationships of Cercidiphyllaceae Ⅰ.Leaf Arehiteeture and Vasculature Changes of petiole[J].Acta Botanica Boreali-Occidentalia Sinica,1990,10(1):37-41.
13.闫小玲.基于广义形态学特征对连香树科系统位置的研究[D].西安:陕西师范大学,2007.
YAN Xiaoling.Systematic Position of Cercidiphyllaceae Based on Morphological Data[D].Xi’an:The Master Degree Dissertation of Shanxi Normal University,2007.
14.郭瑛,赵秋玲,董菊兰.连香树单叶生长动态及其相关性研究[J].甘肃农业科技,2014,(5):38-41.
Guo Yin,Zhao Qiuling,Dong Julan.Study on Dynamic Comparation of Single Leaf and Their Correlation ofCercidiphyllumjaponicum[J].Gansu Agr Sci and Techn,2014,5:38-40.
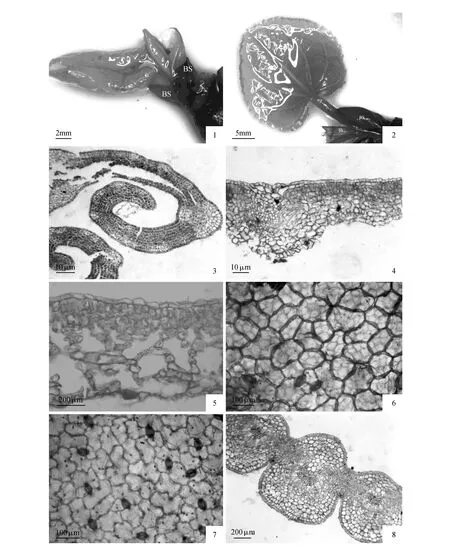
图版Ⅰ 1~2.芽鳞(BS),膜质叶鞘(SH)和幼叶形态;3.幼叶边缘内卷期横切面;4.幼叶横切,示组织分化初期;5.成熟叶片横切;6.上表皮,示表皮细胞形态和气孔器;7.下表皮,示表皮细胞形态和气孔器类型;8.幼叶叶脉横切,示厚角组织和输导组织PlateⅠ 1-2. Bud scale(BS),membranous sheath(SH) and the morphological of immature leaf; 3.Transection structure of immature leaf in stage of margin curl inward; 4.Transection structure of immature leaf, showing tissue differentiation in its early stage; 5.Transection structure of mature leaf; 6.Upper epidermis, showing morphological characteristics of the epithelial cell and the types of stomatal apparatuses; 7.Lower epidermis, showing morphological characteristics of the epithelial cell and the types of stomatal apparatuses; 8.Transection structure of the vein,showing collenchyma and conducting tissue
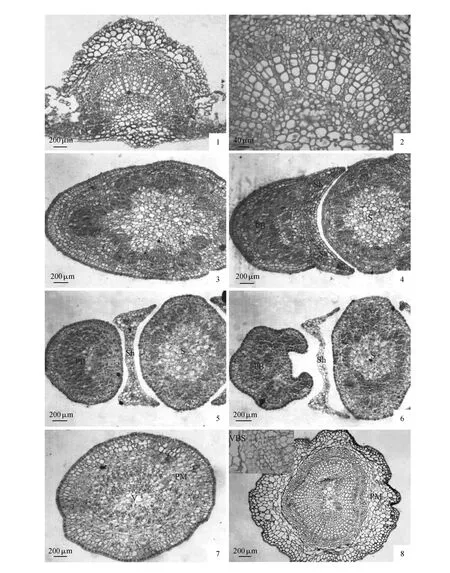
图版Ⅱ 1~2.成熟叶片横切,示叶脉和维管束;3~6.幼叶的叶柄横切,示叶柄形态变化(PB),叶鞘(Sh),幼茎(S);7.幼叶叶柄中段横切(PM),示组织分化;8.成熟叶柄中段横切(PM),示周韧维管束和维管束鞘(VBS)PlateⅡ 1-2.Transection structure of mature leaf,showing the vein and vascular bundles; 3-6.Transection structure of the petiole(PB),showing morphologic changes of the petiole in different parts,the sheath(Sh),immature stem(S); 7.Transection structure in mid-petiole of immature leaf(PM),showing tissue differentiation; 8.Transection structure of mature leaf(PM),showing amphicribral vascular bundle and developed bundle sheaths(VBS)
National Nature Science Foundation Project(31170389);Youth Science Fund Project of the education department of Sichuan Province(2006B038)
date:2016-02-29
LeafDvelopmentandItsEcologicalAdaptabilityofCercidiphyllumjaponicum
MA Yong-Hong1,2
(1.Key Laboratory of Southwest China Wildlife Rsources Conservation,Ministry of Education,Nanchong 637009;2.China West Normal University,Nanchong 637009)
Using the paraffin-section method, we studied the morphological variations and its ecological adaptability ofCercidiphyllumjaponicum, based on the developmental regularities and difference between the immature leaf and mature leaf in anatomical structure. Leaf development were existed in three stages of leaf margin curl inward, expand and mature stage. The first and second pair immature leaves base have a well developed membranous sheath, with the most developed sheath on the first pair immature leaves and more developed sheath on the second, but not found on the others. The leaf ofC.japonicumis typical bifacial leaf with developed spongy parenchyma, outer convex stoma apparatus in the upper and lower epidermis, developed cuticle, vein, mechanical tissue, a amphicribral vascular bundle in min-petiole and bundle sheaths, and theC.japonicumis with strong ecological adaptability to the drought and humid conditions. There are more ergastic substance stored in vegetative organs, which can offer the main nutrition for organ during in breeding season. The first pair immature leaves with faster growth rates also satisfy little nutritional requirements for other organ development.
Cercidiphyllumjaponicum;leaf dvelopment;ecological adaptability;anatomical structure
国家自然基金项目(31170389);四川省教育厅青年科学基金项目(2006B038)资助
马永红(1977—),男,副教授,主要从事植物系统进化和野生植物资源教学和研究。
2016-02-29
Q949
A
10.7525/j.issn.1673-5102.2016.05.011
