5株鼠样品中分离汉坦病毒毒株S基因序列比较与分析
2016-07-28邹春颖马思杰童淑梅
胡 群,邹春颖,马思杰,童淑梅,梅 勇
5株鼠样品中分离汉坦病毒毒株S基因序列比较与分析
胡群,邹春颖,马思杰,童淑梅,梅勇
大榭出入境检验检疫局,宁波315812
摘要:目的了解近年来从宁波口岸捕获的鼠类样品中分离的5个汉坦病毒株遗传学差异。方法利用RT-PCR方法,对5个汉坦病毒株S基因全长进行扩增、克隆和测序,并将这5个毒株的S基因序列分别与GenBank登录的20个汉坦病毒感毒株进行比较分析。结果DX1507株和DX1408株S基因核苷酸序列长度为1766 bp与汉城型(SEOV)病毒株 Rn-Dp7同源性最为接近,LJ1112株S基因核苷酸序列长度为1772 bp与汉城型(SEOV)病毒株Cherwell同源性最为接近,DX0903株和BL1402株S基因核苷酸序列长度为1725 bp与大别山型(DABV)病毒株 Yongjia-Nc-95和Yongjia-Nc-38同源性最为接近。结论宁波口岸分离到的汉坦病毒株分别为汉城型和大别山型,对宁波口岸开展汉坦病毒传播防控有重要意义。
关键词:汉坦病毒;宁波;序列分析;S基因
汉坦病毒(Hantavirus, HV)是重要的人兽共患病病原体,人类感染HV后主要导致2种严重疾病,即肾综合征出血热(hemorrhagic fever withrenal syndrome,HFRS)和汉坦病毒肺综合征(hantaviruspulmonary syndrome,HPS)。HV流行范围之广泛、危害严重,已经成为一个全球性的公共卫生问题[1-2],近年来不断有新型或新亚型病毒被发现,已知HV至少可分为40个血清型/基因型,其中已证实至少有22个型的HV可引起人类疾病[3-4]。
鼠类是HV的主要自然宿主,它感染后无临床症状,生长繁殖也不受影响,并能长时间携带并排出病毒,是人感染HV的主要传染源[5]。在国境口岸开展鼠类携带汉坦病毒的监测,对于防控汉坦病毒的传播具有重要意义。本文就近年来在宁波口岸分离到的5株不同来源的汉坦病毒S基因全序列进行测序分析,并与其他毒株进行比较,了解宁波口岸汉坦病毒的基因型别及遗传变异情况。
1材料与方法
1.1病毒毒株5株汉坦病毒株均为本实验室分离及鉴定。病毒株代号为MS1507为从宁波梅山港区媒介监测捕获的小家鼠中分离;病毒株代号为DX1408为从宁波大榭港区媒介监测捕获的褐家鼠中分离;病毒株代号为DX0903为从宁波大榭港区媒介监测捕获的社鼠中分离;病毒株代号为BL1402为从宁波北仑港区媒介监测捕获的褐家鼠中分离;病毒株代号为LJ1112为从英国入境集装箱中捕获的褐家鼠中分离。
1.2主要试剂RNA提取试剂盒和一步法RT-PCR试剂盒购自Qiagen公司,琼脂糖、5×TBE电泳缓冲液、6×Loading buffer溶液均购自上海生工生物工程技术服务有限公司。
1.3引物参照文献设计[6],上游引物SF:5′-TAGTAGTAGACTCCCTAAAGA-3′,下游引物SR:5′-TAGTAGTAGTATGCTCCCTAA-3′用于扩增S片段全基因。
1.4病毒RNA的制备将病毒株感染Vero E6细胞,培养6~ 7 d后, 用间接免疫荧光法检测细胞感染的程度, 待75%细胞有病毒感染时, 收获病毒, 利用RNA提取试剂盒按说明书从病毒细胞培养液中提取总RNA。
1.5一步RT-PCR反应体系和反应程序一步RT-PCR反应体系为50 μL,无RNA酶水20.2 μL,5×RT-PCR 缓冲液 10 μL,10 mmol/L dNTP 混合液 2 μL,酶混合液 2 μL,RNA酶抑制剂1 μL,20 μmol/L上下游引物SF、SR各2.4 μL,RNA模板10 μL。反应程序为60 ℃ 1 min,42 ℃ 10 min,50 ℃ 30 min,95 ℃ 15 min。反转录结束后,94 ℃ 30 s,52 ℃ 30 s,72 ℃ 1 min,35个循环,72 ℃ 7 min,4 ℃保存。
1.7PCR扩增产物检测、测序PCR扩增产物用1.5%的琼脂糖凝胶电泳,紫外灯观察结果。切下扩增条带,用胶回收试剂盒纯化后,送上海英骏生物技术有限公司进行测序。
1.8基因序列的分析与系统进化树构建用Mega 5.0生物学软件对5个汉坦病毒分离毒株和Genbank录入的三个基因型别的20个汉坦病毒流行株S基因(见表1)进行核苷酸同源性分析,计算遗传距离,并以邻位相连法(Neighbor-joining)绘制系统进化树。
2结果
2.1不同毒株的测序结果将5个毒株的S基因核苷酸序列测序结果拼接后显示,DX1507株和DX1408株S基因核苷酸序列长度为1 766 bp,LJ1112株S基因核苷酸序列长度为1 772 bp, DX0903株和BL1402株S基因核苷酸序列长度为1 725 bp。
表1HV毒株及来源
Tab.1Hantavirus strains and source used in the study

TypeStrainSourceGenBanklocusSEOVGM04-38Shandong,ChinaDQ469397.1HB55Henan,ChinaAF035832.1IR461UKAF458104.1L99Jiangxi,ChinaAF288298.1ZT71Zhejiang,ChinaEF117248.1Rn-DH27ChinaGQ279393.1HuBJ22ChinaGQ279379.1GaomiRn9ChinaGU592941.1CherwellUKKC626089.1HumberUKJX879769.1ShenyangRn139ChinaGU592951.1HTNV76-118KoreaM14627C1-1JapanD25529NC167Anhui,ChinaAB02711584FliShanxi,ChinaAF366569.1LR1Shanxi,ChinaAF288293.1DABVYongjia-Nc-95Zhejiang,ChinaJF796023.1Yongjia-Nc-38Zhejiang,ChinaJF796021.1Wencheng-Nc-470Zhejiang,ChinaJF796019.1Yongjia-Nc-15Zhejiang,ChinaJF796020.1
2.2不同毒株的同源性分析经Mega 5.2软件将5个毒株S基因序列与从GenBank中获得的20个汉坦病毒S基因分别进行核苷酸序列和氨基酸序列的同源性比较,表2可知,DX1507株和DX1408株与汉城型(SEOV)病毒株 Rn-Dp7同源性最为接近,遗传距离分别为0.024和0.015;LJ1112株与汉城型(SEOV)病毒株Cherwell同源性最为接近,遗传距离为0.027;DX0903和BL1402与大别山型(DABV)病毒株 Yongjia-Nc-95和Yongjia-Nc-38同源性最为接近,遗传距离分别为0.062和0.067。
2.3系统进化分析将5个病毒株的S基因全序列与从GenBank上获得的20个参考病毒所构建的系统进化树可知(图1),DX1507,DX1408,LJ1112与汉城型(SEOV)病毒株Rn-Dp7,ShenyangRn139,HuBJ22,GaomiRn9,Cherwell,Humber位于同一组,其中LJ1112与汉城型(SEOV)病毒株 Humbers位于同一分支。DX0903,BL1402与大别山型(DABV)病毒株Yongjia-Nc-95,Yongjia-Nc-38,Wencheng-Nc-470,Yongjia-Nc-15位于同一组。
表25个不同来源病毒与参考病毒株S基因基因距离Tab.2Genetic distance and standard error of L gene of DX1101 with different Hantavirus strains
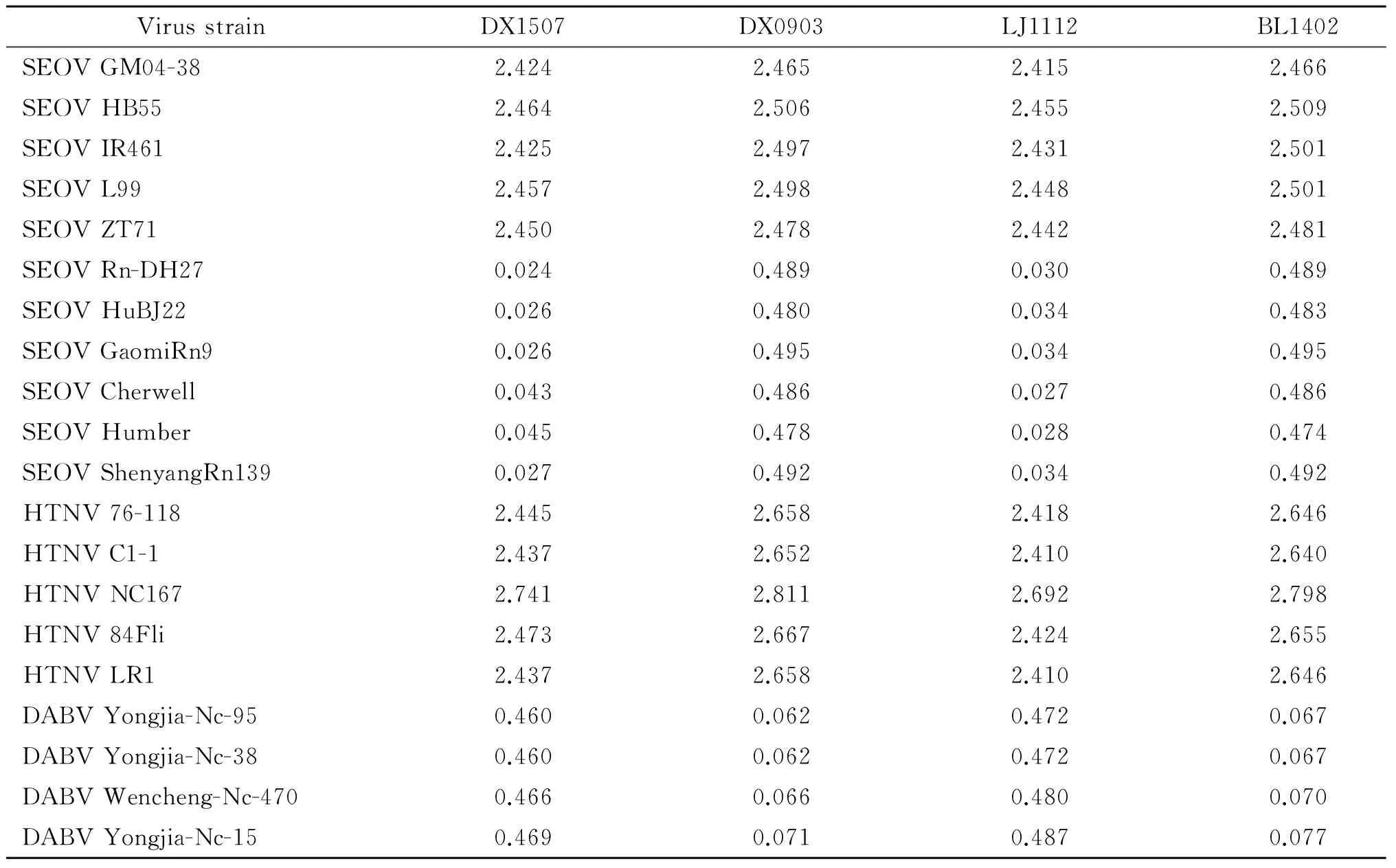
VirusstrainDX1507DX0903LJ1112BL1402DX1408SEOVGM04-382.4242.4652.4152.4662.399SEOVHB552.4642.5062.4552.5092.439SEOVIR4612.4252.4972.4312.5012.410SEOVL992.4572.4982.4482.5012.432SEOVZT712.4502.4782.4422.4812.426SEOVRn-DH270.0240.4890.0300.4890.015SEOVHuBJ220.0260.4800.0340.4830.018SEOVGaomiRn90.0260.4950.0340.4950.020SEOVCherwell0.0430.4860.0270.4860.035SEOVHumber0.0450.4780.0280.4740.037SEOVShenyangRn1390.0270.4920.0340.4920.021HTNV76-1182.4452.6582.4182.6462.428HTNVC1-12.4372.6522.4102.6402.420HTNVNC1672.7412.8112.6922.7982.703HTNV84Fli2.4732.6672.4242.6552.439HTNVLR12.4372.6582.4102.6462.420DABVYongjia-Nc-950.4600.0620.4720.0670.452DABVYongjia-Nc-380.4600.0620.4720.0670.452DABVWencheng-Nc-4700.4660.0660.4800.0700.459DABVYongjia-Nc-150.4690.0710.4870.0770.465
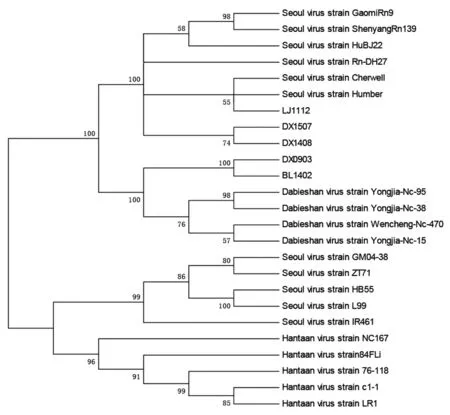
图1 不同基因型HV巢式RT-PCR SEO特异性引物S基因扩增产物构建的系统发生树Fig.1 Phylogenic tree of S gene of Hantavirus PCR product
3讨论
汉坦病毒是一种单股负链 RNA 病毒, 核酸基因组由大(L)、中(M)、小(S)3个节段组成,其中 S片段编码病毒核衣壳蛋白(NP),在病毒感染及复制过程中具有重要的作用[7,8],而且S基因具有相对的保守性,可通过对S基因的测定和分析来获取汉坦病毒的进化和变异信息,从而为汉坦病毒的进化研究和流行病学的调查发挥重要作用。
本研究共分析了5个汉坦病毒株的S基因全序列,在毒株选择上考虑了宿主种类和地域等因素,是本实验室近年来从鼠类样品中检测分离的代表株。从分析结果来看,宁波口岸5株汉坦病毒株,其中DX1507,DX1408,LJ1112基因型别为汉城型(SEOV),并且属于(SEO)和大别山(DOB)两个基因型,其中大别山型已证实在国内浙江地区流行传播,该基因型别的汉坦病毒可能来源于宿主溢出,即汉坦病毒在水平传播过程中,除在原宿主内流行外,随机感染了另一种动物,并发生病毒基因组变异,以适应新宿主的遗传环境,随宿主加以进化[9,10]。
病毒株LJ1112是从英国入境的集装箱中检获的鼠体中分离,从遗传距离和系统进化分析,可以知道与英国流行株的亲源关系,这也进一步证实了传染病病原体可能过鼠类等医学媒介在国际间传播,这对于加强口岸媒介病原体监测具有重要的意义。
参考文献:
[1] Li JL, Li DX, Progressing on the entiology of Hantaviruses[J].CHINESE J Exp & Cli Vir, 2005,19(2):198-201.
李家亮, 李德新. 汉坦病毒病原学研究进展[J].中华实验和临床病毒学杂志, 2005, 19(2): 198-201.
[2] Mertens M, Essbauer SS, Rang A, et al. Non-human primates in outdoor enclosures: Risk for infection with rodent-borne hantaviruses[J].Vet Microbiol, 2011, 147(3): 420-425.
[3] Zhou JH, Zhang HL. Genotyping and reasearch progress of hantaviruses[J]. Int J Vir, 2008,15(2):37-42.
周济华, 张海林. 汉坦病毒分型及致病性的研究进展[J]. 国际病毒学杂志, 2008, 15(2): 37-42.
[4] Zhang HL, Zhan YZ. Distribution and genetyping of hantaviruses in china[J]. Chin J vector & control, 2011,22(5):417-420.
张海林,张云智. 中国汉坦病毒基因型及分布[J]. 中国媒介生物学及控制杂志,2011,22(5): 417-420
[5] Wang XJ, Chen JY. Epidemiological analysis of epidemic hemorrhagic fever in China[J]. J Med Pes Con, 2000,16(4):2015-217.
王学娟,陈继永. 我国流行性出血热流行病学研究现状[J]. 医学动物防制,2000,16(4):215-217.
[6] Li L, YANG DJ, Su X, et al. Direct sequence analysis of hantaviruses S gene in rodent lung tissue. Tianjin Med J, 2005,33(10):653-656.
李力, 杨东靖, 苏旭等. 鼠肺组织直接用于汉坦病毒S基因序列分析[J]. 天津医药,2005,33(10): 653-656。
[7] Schmaljohn CS, Hasty SE, Dalrymple JM,et al. Antigentic and gentic proprerties of viruses linked to HFRS[J].Science, 1985,227(4690):1041-1044.DOI: 10.1126/science.2858126.
[8] Eliott LH, Kiley MP, Mc Cormick JB, et al. Hantaan virus: identification of virion proteins[J].J Gen Virol, 1984,65(8) :1285-1293.DOI:10.1099/0022-1317-65-8-1285.
[9] Lin XD, Wang W, Guo WP, et al. Cross-species transmissinon in the speciation of the currently known murinae-associated hantaviruses[J]. J virology, 2012, 86(20):11171-11182.DOI:10.1128/JVI.00021-12
[10] Ahn C, Cho J T,Lee J G,et al. Detection of Hantaan and Seoul viruses by reverse transcriptase-polymerase chain reaction(RT-PCR) and restriction fragment length polymorphism (RFLP) in renal syndrome patients with hemorrhagic fever[J]. Clin Nephrol, 2000, 53(2):79-89.DOI: 10.1128/JCM.39.7.2397-2404.2001.
DOI:10.3969/j.issn.1002-2694.2016.05.017
中图分类号:R373
文献标识码:A
文章编号:1002-2694(2016)05-502-04
收稿日期:2015-10-14修回日期:2016-01-26
Sequence comparison and analysis of S Gene of five Hantavirus strains separated from rodents
HU Qun, ZHOU Chun-ying,MA Si-jie, TONG Shu-mei, MEI Yong
(DaxieExit-EntryInspectionandQuarantineBureau,Ningbo315812,ZhejiangProvinca,China)
Abstract:To know the genetic differences of five hantavirus strains separated from rodents captured in Ningbo port area. The S genes of five hantavirus strains were amplified, cloned and sequenced. Then the S gene sequences of these strains were compared with that of 20 hantavirus strains in GenBank. The results showed that the DX1507 and DX1408 strains S segment are 1766 bp in length and with closest homology to the SEOV virus strain Rn-Dp7, the LJ1112 strains S segment are 1772 bp in length and with closest homology to the SEOV virus strain Cherwell, the DX0903 and BL1402 strains S segment are 1725 bp in length and with closest homology to the DOBV virus strain Yongjia-Nc-95and Yongjia-Nc-38. This study indicated the five hantavirus strains separated from rodents captured in Ningbo port area are belong to SEO or DOBV type, which has an important significant to control hantavirus spreading in Ningbo port.
Key words:Hantanvirus; Ningbo; sequence analysis; S gene
