茉莉酸甲酯对苋菜中苋红素积累的影响
2016-05-09朱昌华甘立军
金 颢, 朱昌华, 夏 凯, 甘立军
(南京农业大学 生命科学学院, 南京 210095)
茉莉酸甲酯对苋菜中苋红素积累的影响
金 颢, 朱昌华, 夏 凯, 甘立军
(南京农业大学 生命科学学院, 南京 210095)
摘要苋红素是一种重要的次生代谢物质,其合成受到光照等因素的诱导。分别以苋菜(Amaranthus mangostanus L.)子叶以及四叶期幼苗为材料,研究了茉莉酸甲酯(methyl jasmonate, MeJA)在苋红素积累中的作用。结果表明:0.02~20 mg/L MeJA在光照及黑暗下均能显著促进苋菜子叶苋红素的积累,且具有明显的浓度效应。在光照下20 mg/L MeJA处理所诱导的苋红素含量是对照组(光照)子叶苋红素含量的2.23倍,同时是黑暗下不加MeJA的对照组的10.91倍。对苋菜幼苗而言,茉莉酸甲酯可以促进苋菜幼苗中苋红素的积累,1.25~10 mg/L范围的茉莉酸甲酯均有效促进了苋红素的积累,且10 mg/L 作用效果最显著。但高浓度的茉莉酸甲酯会抑制苋菜幼苗的生长及叶片的展开。进一步的研究发现,MeJA可引起酪氨酸酶活的上升进而促进苋红素的合成。综上所述,茉莉酸甲酯可以促进苋菜子叶以及幼苗叶片和茎秆中苋红素的积累,但同时对于苋菜幼苗的生长有一定的抑制效应。
关键词苋菜;茉莉酸甲酯;机械伤害;苋红素
Inductive effect of methyl jasmonate on amaranthin accumulation in Amaranthus mangostanus L. seedlings
JIN Hao, ZHU Chang-hua, XIA Kai, GAN Li-jun
(College of Life Sciences, Nanjing Agricultural University, Nanjing 210095, China)
AbstractAmaranthin is one of the important plant secondary metabolites. Light can trigger the initiation of amaranthin biosynthesis in cotyledons ofAmaranthusmangostanusL. In this paper, withAmaranthusmangostanusL. as the material, the effect of MeJA (Methyl Jasmonate) on amaranthin synthesis was explored. MeJA ( 0.02-20 mg/L) induced amaranthin accumulation was dose-dependent both in light and darkness. When in darkness, 20 mg/L MeJA induced 10.91 fold amaranthin contents that of controls. While in light, the 20 mg/L MeJA induced 2.23-fold amaranthin in cotyledons exposed to light only. When used 4-leaf stage seedlings as material, MeJA (1.25-10 mg/L) could promote amaranthin accumulation both in cotyledons, leave, and stems ofAmaranthusmangostanusL. effect of 10 mg/L MeJA was the best, but high concentrations of MeJA could inhibit the growth of the seedlings and leaf expansion. The following experiments indicated that MeJA could induce amaranthin accumulation by promoting the activity of tyrosinase. From above, MeJA could promote amaranthin accumulation both in seedlings and cotyledon ofAmaranthusmangostanusL., but the application of MeJA had inhibitory effect on the plant growth and development.
KeywordsAmaranthusmangostanusL.; methyl jasmonate; mechanicnl wounding; amaranthin
苋红素作为一种重要的次生代谢物质[1],在生物制药、食品添加、工业制造等领域应用广泛,其产生的机制也被人们关注并研究[2]。早先的研究发现光是调控苋红素合成的重要因素[3, 4]。另外,细胞分裂素类物质如激动素、玉米素、6-BA等均可以促进在光照条件下或黑暗下苋属植物苋菜红素类物质的积累[5, 6]。作为甜菜红素中的一种,其合成中研究较多的酶类有酪氨酸酶、多巴双加氧酶、葡糖基转移酶等[7]。研究人员在太阳花愈伤组织中检测到酪氨酸羟化酶的活性[8],该酶是检测甜菜红素合成的重要指标。同时,果蔬在生产、加工运输等过程中很容易受到机械伤害,因此,伤害以及茉莉酸对植物的影响的研究也一直为人们所关注[9]。
茉莉酸甲酯(methyl jasmonate, MeJA)是调控植物次生代谢物合成的重要激素[10],可以作为信号分子激发植物的化学防御[11],植物在受到机械损伤后,往往会产生一些诸如酚类、生物碱、萜类等次生代谢物质[10, 12]。比如外源施用MeJA能提高花烛佛焰苞花青苷的含量[13];伤害以及茉莉酸可以提高喜树幼苗中喜树碱的含量[14];茉莉酸甲酯可以促进马齿苋属(Portulaca)中甜菜红素的积累等[9]。但MeJA对苋红素的合成还未见系统地报道,因此我们探究了外源茉莉酸甲酯对于苋菜子叶以及幼苗中苋红素积累的影响。
1材料和方法
1.1材料培养
1.1.1苋菜种子处理选取大小、饱满度均一的苋菜(AmaranthusmangostanusL.)种子,经去离子水冲洗以及1%次氯酸钠消毒10 min。将种子均匀播于铺有3层滤纸的培养盘中黑暗萌发48 h(相对湿度为70%,温度25℃±0.5℃)后,将已萌发的苋菜种子于绿光灯下取出,去除种皮,并选取长势一致的材料,用剪刀剪去根部及部分胚轴,留下长约3~4 mm实验用。
1.1.2苗期苋菜的培养将苋菜(AmaranthusmangostanusL.)种子均匀铺散于小花盆中,采用蛭石:营养土(体积比为2∶1)培养幼苗。所有苋菜幼苗均于南京农业大学牌楼实验基地的温室中培养(白天温度18℃~26℃;夜间温度12℃~15℃)。待苋菜苗长至4叶期,用不同方式处理。之后每隔1 d取样测定各项指标。
1.1.3苋菜子叶期处理茉莉酸甲酯处理:将30根子叶均匀放于铺有2层滤纸的9 mm培养皿中,将茉莉酸甲酯溶液(茉莉酸甲酯用少量乙醇助溶,之后稀释为所用浓度)加入培养皿中,其浓度梯度为:0、0.02、0.2、2和20 mg/L。光照组的培养皿放于光下(10 000 lx)照射9 h之后转至黑暗中,两组同样在24 h时分别取样并拍照观察。对照组的培养皿中加入磷酸缓冲液(pH 6.3)。实验重复5次以上。
1.1.4苗期茉莉酸甲酯处理待温室中的苋菜幼苗长至4叶期时,用茉莉酸甲酯对幼苗喷施处理,浓度为:0、1.25、2.5、5和10 mg/L。对照组喷施清水。处理当天以及之后每隔1 d进行取样测定苋红素的含量,另外在处理后第10天,取样测定苋菜生长指标及苋红素的含量。实验重复3次以上。
1.2测定指标
1.2.1苋菜幼苗叶片及茎秆中苋红素含量的测定
参照Piattelli[15]的方法并进行一定的改进。分别称取苋菜幼苗的第4、第5及第6叶片及茎秆的鲜重,使用液氮研磨充分,加入适量的去离子水,在4℃下8 000 r/min离心15 min,之后取上清液用于531 nm处吸光值的测定。根据甜菜素的消光系数5.66×104计算苋红素的含量[23]。
1.2.2酪氨酸酶活性的测定参照Yamamoto[8]的方法,并进行一定的修改。3 mL反应液(60 mmol/L磷酸缓冲液(pH 6.3); 2 mmol/L酪氨酸,30 mmol/L抗坏血酸以及0.1 mg/L过氧化氢酶),在25℃下放置15 min后,加入200 μL酪氨酸酶提取液,反应开始,30 min后加入2 mL 50%的磷酸溶液终止反应。之后于4℃下10 000 r/min离心15 min,用上清液在279 nm 测定其吸光度,使用煮过的酪氨酸酶提取液,按照同样的操作步骤作对照。定义吸光度改变0.01为一个酪氨酸酶活力单位,酶活性以U/mg·protein表示。实验重复3次以上。
1.3数据处理和分析
用GraphPad5.01、Excel2003及PhotoShopCS4等进行作图及数据分析,用Spass16.0进行差异显著性分析。
2结果与分析
2.1茉莉酸甲酯促进子叶中苋红素的积累

图1 茉莉酸甲酯在光/暗下促进苋菜子叶中苋红素积累的效应
其中光照强度为10 000 lx,温度25℃; 所有处理放于黑暗中保存24 h再进行苋红素的测定;a、b、c等字母表示各组MeJA处理的苋红素含量在P< 0.05水平上的显著性差异。
如图1所示,光/暗下MeJA均可以促进苋菜子叶中苋红素的积累。黑暗下,0.02 mg/L MeJA诱导苋菜子叶苋红素的合成量是对照的2.7倍,当MeJA的浓度为20 mg/L时,黑暗下诱导的苋红素的含量是对照组的5.96倍。光和MeJA的共同诱导可以促进更多的苋红素的合成。20 mg/L 的MeJA光下所诱导的苋红素含量是仅光照9 h所诱导苋红素含量的2.23倍,同时是暗下对照组的10.91倍。
2.2茉莉酸甲酯对苋菜幼苗株高和地上部鲜重的影响
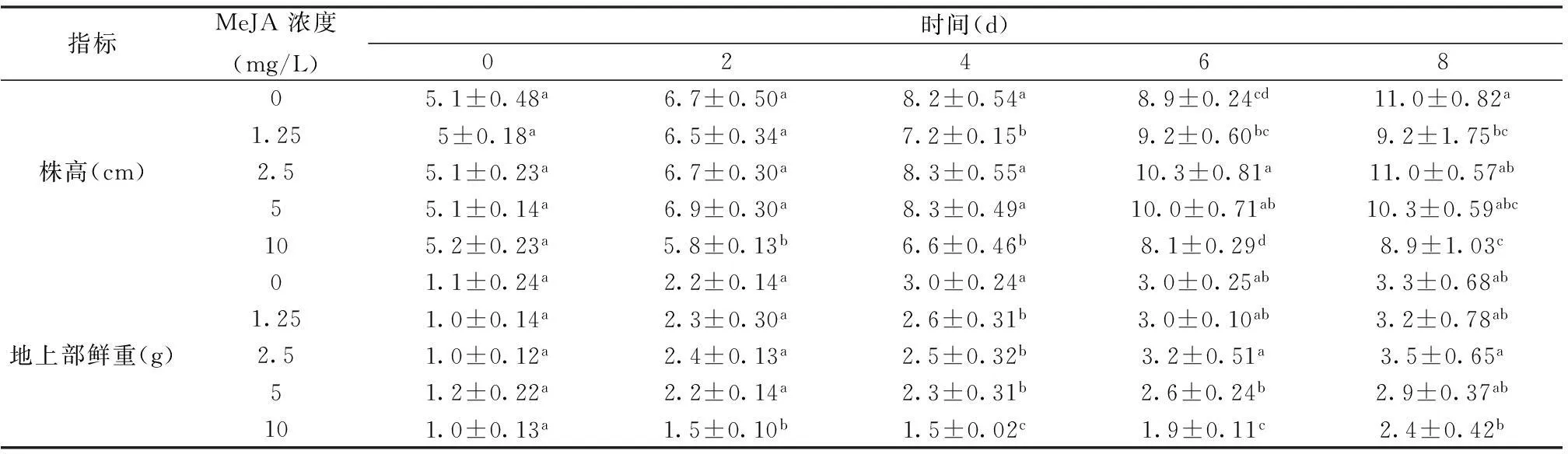
表1 茉莉酸甲酯对苋菜幼苗株高以及地上部鲜重的影响
在第0天时喷施MeJA,每隔1 d采样测定数据;小写字母a、b、c、d等表示不同浓度MeJA处理组的株高及地上部鲜重在P< 0.05水平上有显著性差异。

表 2 茉莉酸甲酯对苋菜幼苗根长和地下部鲜重的影响
在第0天时喷施MeJA,每隔1 d采样测定数据;小写字母a、b、c、d等表示不同浓度MeJA处理组的根长及地下部鲜重在P< 0.05水平上有显著性差异。

图 2茉莉酸甲酯处理对苋菜幼苗叶片及
Fig 2 Effect of MeJA on amaranthin accumulation in the leaf and the stem ofAmaranthusmangostanusL. Bars with a, b, c etc indicate significant differences compared to corresponding control samples atP< 0.05 level.
a: 茎秆; b: 第4叶; c: 第5叶; d: 第6叶。
由表1可见,在处理第8天时,5 mg/L MeJA处理的幼苗株高是对照的93%,地上部鲜重是对照的87%;而10 mg/L MeJA处理的苋菜幼苗的株高在第8天时,株高是对照的80%,地上部鲜重是对照的72%。以上说明,对苋菜幼苗喷施MeJA时,低浓度的溶液对于幼苗的株高及地上部鲜重无明显影响;高浓度的溶液对幼苗的株高的伸长以及地上部鲜重出现显著的抑制作用。
2.3茉莉酸甲酯对苋菜幼苗根长和地下部鲜重的影响
由表2可知, 10 mg/L MeJA处理的苋菜幼苗的根长有明显下降趋势,在处理第8天时,根长是对照组的63%,地下部鲜重却与对照组相当,差异不显著, 结合实际分析,在MeJA处理后,苋菜幼苗的不定根的数量增多,使得根部虽长度受到抑制,但重量仍不受影响甚至有所增加。
2.4茉莉酸甲酯处理对苋菜叶片及茎秆中苋红素积累的浓度效应
由图2可见,用不同浓度的MeJA处理苋菜幼苗10 d后,MeJA的浓度为5 mg/L时,茎秆中的苋红素的含量与对照相比,差异达到显著水平,而当MeJA的浓度为10 mg/L时,茎秆部位的苋红素的含量是对照的1.70倍。对于幼苗的第4、第5及第6叶片,1.25~10 mg/L范围的MeJA均有效促进了苋红素的积累,且10 mg/L 作用效果最显著,第4、第5及第6张叶片中苋红素的含量分别是对照的2.53、4.22及5.11倍。
2.5 茉莉酸甲酯处理时苋菜叶片及茎秆中苋红素的时间变化
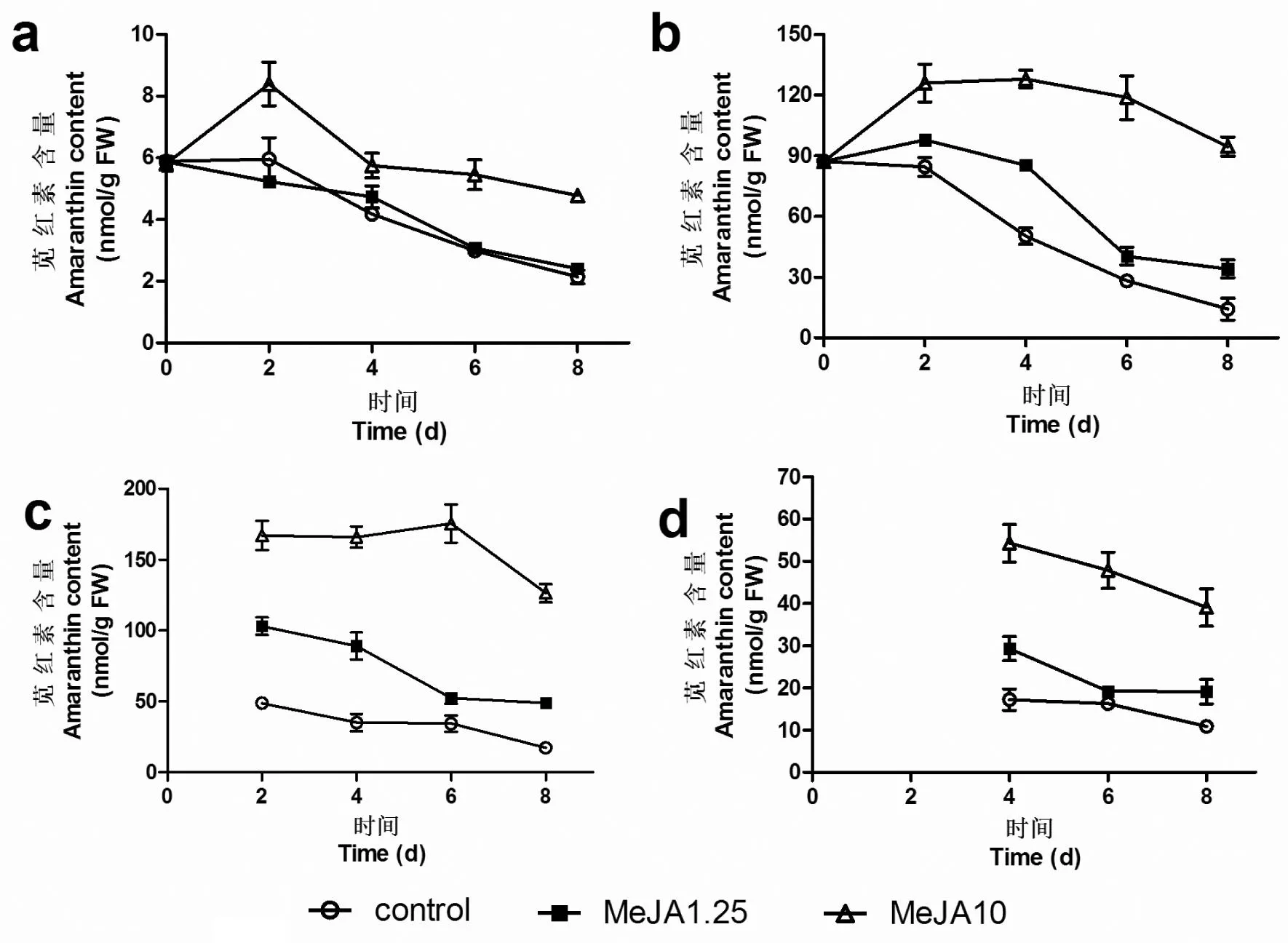
图 3 茉莉酸甲酯处理后苋菜幼苗叶片及
a: 茎秆; b: 第4叶; c: 第5叶; d: 第6叶; control:喷施清水处理; MeJA1.25:MeJA喷施浓度为1.25 mg/L; MeJA10: MeJA喷施浓度为10 mg/L。
由图3可见,苋红素含量随着时间的推移均呈现下降的趋势。在第10天时, 对照组茎秆中苋红素含量是处理当天的36.4%,而第4、第5及第6叶中的含量分别是处理当天(五叶、六叶与新生叶完全长出时比较)的35.9%、16.8%和49%。10 mg/L MeJA处理组的苋红素含量一直在较高水平,下降趋势也较对照组缓慢。结果表明,在苋菜的生长过程中,不同部位的苋红素的含量会随着时间的推移而降低,且老叶中苋红素的含量高于新生叶中的含量。
2.6茉莉酸甲酯及机械伤害对苋菜处理叶片中酪氨酸酶活性的影响
由图4可知,MeJA处理幼苗后,叶片中的酪氨酸酶的羟化活性呈现上升趋势,在处理后1 d,MeJA处理组叶片酪氨酸酶的羟化活性达到62 U/mg·Pro,是处理当天叶片的1.76倍,但在之后的几天里,MeJA处理组的酶活呈现下降的趋势;机械伤害组的叶片中酪氨酸酶的羟化活性也在处理后出现上升的现象,在处理后第2天达到最高,56 U/mg·Pro,而在之后,机械伤害组的酪氨酸酶羟化活性随时间推移而出现下降。用MeJA和机械伤害处理后,叶片中的酪氨酸酶均呈现先上升后下降的趋势,也许是因为酶活性的丧失等因素造成。
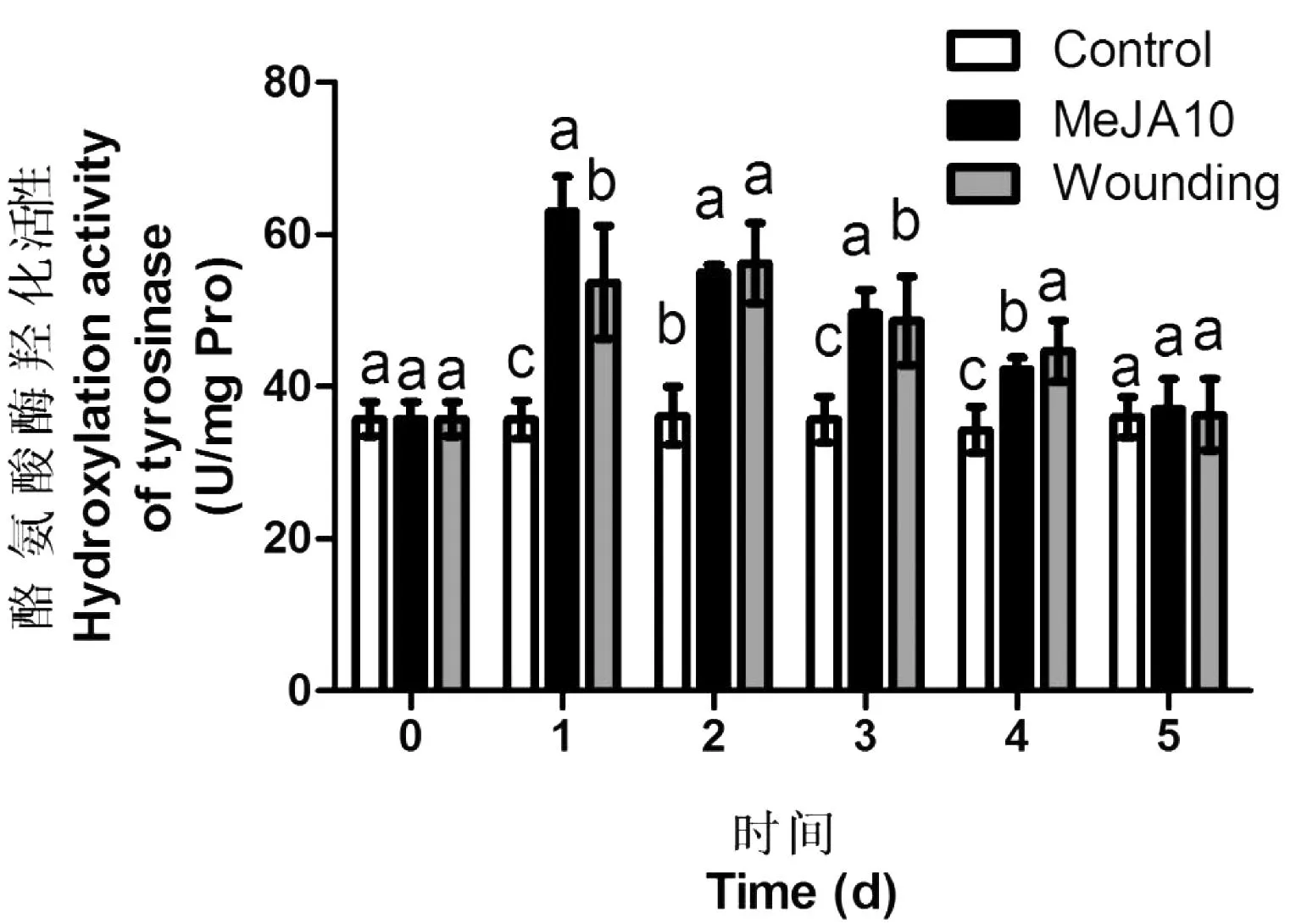
图4 茉莉酸甲酯以及机械伤害处理后苋菜幼苗叶片中
Control:喷施清水处理; MeJA10:MeJA喷施浓度为10 mg/L; Wounding:机械伤害处理。
3讨论
环境或者一些生物非生物胁迫均会引起植物次生物质的代谢,伤害是一种常见的刺激信号,可以激发植物体内茉莉酸信号的转导,进而引起植物的防御系统,加强了植物次生代谢,使植物获得抗性。同时,茉莉酸甲酯是调控植物次生代谢物合成的重要激素,可以调控多种植物次生代谢物质的产生。在本文中,我们发现,茉莉酸甲酯明显促进苋菜子叶以及幼苗中苋红素的积累。Cao等[16]研究表明,茉莉酸可以作为诱导物强烈促进苋菜萌发期时苋红素的积累,且在茉莉酸浓度为100 μmol/L时,苋红素的积累量达到最高,我们的结果与之相似。
同时,茉莉酸甲酯作为伤信号,外源施加于植物时往往给植物的生长发育带来很大的影响。Singh等[17]的研究表明,在水稻中,外源施加茉莉酸甲酯可以缓解钙离子引起的氧化性胁迫,但同时也在一定程度上抑制了水稻幼苗的生长。谢阳姣等[18]的研究表明,用茉莉酸甲酯喷施苦玄参幼苗,虽然提高了苦玄参苷的积累量,但同时会抑制其株高、叶片以及分枝的生长,将其成熟期提前,缩短了其生育期。在本文中,叶面喷施茉莉酸甲酯在一定程度上抑制了苋菜幼苗株高的生长以及叶片的展开,降低了幼苗生物量的积累。所以,在生产中若是为了提升生物量,茉莉酸甲酯的应用会使苋菜的生育期缩短,生物量下降,这无疑在一定程度上限制了其应用;但是若以获得更多的苋红素的产量为目的,可以考虑合理地应用茉莉酸甲酯,选择合适的浓度,在幼苗将近成熟的时期使用,将会获得较好的效果。
参考文献:
[1]Strack D, Vogt T, Schliemann W. Recent advances in betalain research[J]. Phytochemistry, 2003, 62(3): 247-269.
[2]Cai Y, Sun M, Corke H. Characterization and application of betalain pigments from plants of theAmaranthaceae[J]. Trends in Food Science & Technology, 2005, 16(9): 370-376.
[3]Kochhar V K, Kochhar S, Mohr H. An analysis of the action of light on betalain synthesis in the seedling ofAmaranthuscaudatus, var.Viridis[J]. Planta, 1981, 151(1): 81-87.
[4]G ring H, Drfler M. Amaranthin accumulation in continuous red and blue light by seedlings ofAmaranthuscaudatusL.[J]. Biologia Plantarum, 1981, 23(3): 193-197.
[5]Stobart A K, Pinfield N J, Kinsman L T. The effects of hormones and inhibitors on amaranthin synthesis in seedlings ofAmaranthustricolor[J]. Planta, 1970, 94(2): 152-155.
[6]Stobart A K, Kinsman L T. The hormonal control of betacyanin synthesis inAmaranthuscaudatus[J]. Phytochemistry, 1977, 16(8): 1139-1142.
[7]Han X H, Gao Z J, Xiao X G. Enzymes and genes involved in the betalain biosynthesis in higher plants[J]. African Journal of Biotechnology, 2009, 8(24):6735-6744.
[8]Yamamoto K, Kobayashi N, Yoshitama K, et al. Isolation and purification of tyrosine hydroxylase from callus cultures ofPortulacagrandiflora[J]. Plant and Cell Physiology, 2001, 42(9): 969-975.
[9]Bhuiyan N H, Adachi T. Stimulation of betacyanin synthesis through exogenous methyl jasmonate and other elicitors in suspension-cultured cells ofPortulaca[J]. Journal of Plant Physiology, 2003, 160(9): 1117-1124.
[10]Santino A, Taurino M, De Domenico S, et al. Jasmonate signaling in plant development and defense response to multiple (a) biotic stresses[J]. Plant Cell Reports, 2013, 32(7): 1085-1098.
[11]Ketabchi S, Majzoob S, Charegani H A. Effect of salicylic acid and methyl jasmonate on phenylalanine ammonia-lyase activity and total phenol in wheat infected byPratylenchusthornei[J]. Archives of Phytopathology and Plant Protection, 2015, 48(1): 10-17.
[12]Shahabinejad M, Shojaaddini M, Maserti B, et al. Exogenous application of methyl jasmonate and salicylic acid increases antioxidant activity in the leaves of pistachio (PistaciaveraL. cv.Fandoughi) trees and reduces the performance of the phloem-feeding psyllidAgonoscenapistaciae[J]. Arthropod-Plant Interactions, 2014, 8(6): 525-530.
[13]Ram M, Prasad K V, Singh S K, et al. Influence of salicylic acid and methyl jasmonate elicitation on anthocyanin production in callus cultures ofRosahybridaL.[J]. Plant Cell Tissue and Organ Culture, 2013, 113(3): 459-467.
[14]牧 文, 刘文哲. 伤害及诱导子对喜树幼苗中喜树碱含量的影响[J]. 西北植物学报, 2007, 27(5): 977-982.
[15]Piattelli M, Nicola M G D, Castrogiovanni V. The effect of kinetin on amaranthin synthesis inAmaranthustricolorin darkness[J]. Phytochemistry, 1971, 10(2): 289-293.
[16]Cao S, Liu T, Jiang Y, et al. The effects of host defence elicitors on betacyanin accumulation inAmaranthusmangostanusseedlings[J]. Food Chemistry, 2012, 134(4): 1715-1718.
[17]Singh I, Shah K. Exogenous application of methyl jasmonate lowers the effect of cadmium-induced oxidative injury in rice seedlings[J]. Phytochemistry, 2014, 108:57-66.
[18]谢阳姣, 何志鹏, 林 伟. 茉莉酸甲酯对苦玄参生长及苦玄参苷积累的影响研究[J]. 作物杂志, 2013(2): 80-84.
中图分类号Q946;TS264.4
文献标识码A
文章编号2095-1736(2016)02-0054-05
作者简介:金颢,硕士研究生,从事植物激素生理和生物技术研究,E-mail:806496252@qq.com。通信作者:甘立军,副教授,从事植物激素生理生化研究, E-mail:ganlj@njau.edu.cn。
基金项目:国家自然科学基金项目(30300216)
收稿日期:2015-08-05;修回日期:2015-08-28
doi∶10.3969/j.issn.2095-1736.2016.02.054
