3种不同饵料对越冬后再摄食至第一次蜕壳前中华绒螯蟹幼蟹能量密度和营养积累研究
2016-05-09王福辰马明君成永旭
王福辰, 王 春, 马明君, 成永旭
(上海海洋大学 淡水水产重点实验室, 上海 201306)
3种不同饵料对越冬后再摄食至第一次蜕壳前中华绒螯蟹幼蟹能量密度和营养积累研究
王福辰, 王春, 马明君, 成永旭
(上海海洋大学 淡水水产重点实验室, 上海 201306)
摘要以3种不同饵料饲喂越冬后再摄食至第一次蜕壳前的中华绒螯蟹幼蟹,测试全个体能量密度、肝胰腺总脂和粗蛋白含量、肌肉总脂和粗蛋白含量。结果显示,个体能量密度呈先上升、后下降的走势,且配合饲料组幼蟹再摄食早期显著高于再摄食前和临近蜕壳前(P<0.05);临近蜕壳前与再摄食前的能量密度均无显著性差异(P>0.05);大规格幼蟹的能量密度始终高于小规格幼蟹,差异不显著(P>0.05);不同饵料组同规格幼蟹的能量密度,玉米组高于鲫鱼肉组和饲料组,且大规格差异显著(P<0.05)。肝胰腺总脂和粗蛋白含量未出现显著性差异(P>0.05)。小规格幼蟹肌肉总脂和粗蛋白均呈显著下降走势(P<0.05),而大规格幼蟹肌肉总脂无显著变化(P>0.05),肌肉粗蛋白含量显著升高(P<0.05)。
关键词中华绒螯蟹幼蟹;越冬后再摄食;能量密度;营养生化
Studies on energy density and nutrient accumulation in the young Eriocheir sinensis living after the last winter before the first molting in the second year
WANG Fu-chen, WANG Chun, MA Ming-jun, CHENG Yong-xu
( Key Laboratory of Freshwater Aquatic Genetic Resources, Ministry of Agriculture,Shanghai Ocean University, Shanghai 201306, China)
AbstractDifferent body-size young Chinese mitten crabs (Eriocheirsinensis), living from the end of the last winter to the beginning of the first molting in the second year, were fed with three different diets, the corn, the crusion carp meat, and the formulated diet. And the energy density of the whole crab, the content of total lipid and crude protein in the hepatopancreas and muscle were determined. The result showed that, the energy density of the whole crab rised firstly, and then declined to the same level with that before feeding. The energy density early in the feeding of formulated diet group was significantly higher than that of the other groups (P<0.05). The energy density of big body-size crabs was higher than that of small body-size, but the difference was not significant (P>0.05). And the energy density of the big body-size crab in the group fed with corn was significantly higher than that of the others (P<0.05). The content of total lipid and crude protein in the hepatopancreas in all groups showed no significant differences (P>0.05). The content of both total lipid and crude protein in the muscle of small body-size crabs declined significantly (P<0.05), while the total lipid of big body-size crabs showed no significant differences (P>0.05), but the content of crude protein rised significantly (P<0.05).
KeywordsChinese mitten crabs (Eriocheirsinensis); refeeding after winter; energy density; biochemical nutrition
中华绒螯蟹(Eriocheirsinensis)俗称河蟹、大闸蟹,属广食性(或杂食性)动物,其食物主要包括动物饵料、植物饵料,以及有机碎屑等。人工养殖中一般投喂的饵料包括野杂鱼、螺蛳、玉米、豆粕及配合饲料等[1]。中华绒螯蟹养殖生产上经常发现因蜕壳失败而死亡的个体,或者蜕壳后个体由于新壳不能及时硬化而死亡的现象,这种现象被称之为蜕皮综合征(MDS),MDS形成可能与甲壳动物蜕壳前体内的营养物质积累不足或不平衡有关,如脂类、蛋白等营养物质[2]。能量物质是生物体进行生理代谢活动的基本条件,动物获取能量物质的主要途径为摄食,其可利用的能量物质主要有3种,即脂类、蛋白质和碳水化合物。已有的研究指出,中华绒螯蟹的主要能量物质是脂类物质和蛋白质,肝胰腺是甲壳动物体内脂类主要的吸收和储存器官[3-7],而肝胰腺中的脂类成分更容易受饵料成分的影响[8-11]。
中华绒螯蟹在越冬结束后再摄食至第一次蜕壳前的这一阶段,关于幼蟹体内营养和能量物质的积累状况尚没有相关研究。本实验选用生产上常用的3种不同类型饵料,即玉米、鲫鱼肉和配合饲料,分别饲喂2种不同规格的幼蟹,通过测试幼蟹的各项生化指标,包括全个体能量密度、肝胰腺总脂和粗蛋白含量、肌肉总脂和粗蛋白含量,旨在探究越冬结束后再摄食至第一次蜕壳前,幼蟹在营养和能量物质积累方面表现出的规律;对比不同规格幼蟹在营养积累方面是否存在差异性;并对不同类型的饵料对该阶段幼蟹营养强化的效果进行比较。
1材料与方法
1.1实验幼蟹养殖及样品采集
本实验选取的中华绒螯蟹幼蟹,来自于崇明岛上海海洋大学产学研基地,于同一时间和地点采集,挑选十足完整健全、有活力、未早熟的幼蟹,以体重为标准分两种类型:小规格(5.17±1.21)g/只,大规格(11.13±1.05)g/只。养殖实验所投喂的饵料包括:干玉米粉碎颗粒(粒径小于1.0 mm),冰鲜鲫鱼(Carassiusauratus)肉,配合饲料为本实验室研制并长期固定使用的河蟹幼蟹饲料。实验用幼蟹饲养于室内循环水养殖系统中,水温保持18~20℃,单体隔离饲养,饱食投喂。
以投喂的3种不同饵料和大小2种规格为区别分成6个实验组,每个实验组设3个重复平行。分别于3个时间点进行样本采集,即第一次投喂饵料之前采集再摄食前的样本,于喂食满5 d时采集再摄食早期的样本,于养殖20 d左右、幼蟹表现出明显蜕壳迹象时采集临近蜕壳前的样本。每一批幼蟹样本采集后于实验室暂养箱中饥饿1 d,以消除消化道中食物对生化指标的影响。之后迅速解剖致死,分别收集全个体样品、肝胰腺样品和肌肉样品,并全部放置于-20℃冰箱保存。
3种饵料的能量密度、总脂含量、粗蛋白含量,经生化测试结果如表1所示。
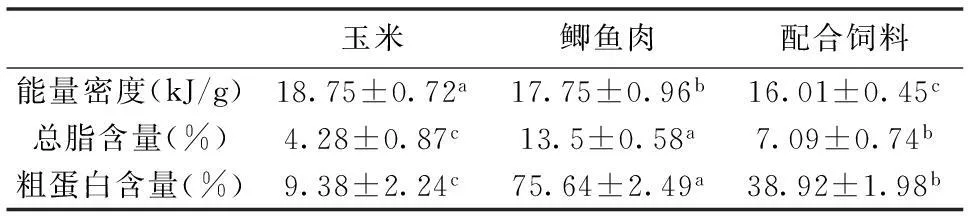
表1 3种饵料能量密度、总脂及粗蛋白含量(干重)
上标不同小写字母表示同一行差异显著(P<0.05)。
1.2生化测试方法及数据分析
总脂含量采用Folch[12]等法测定,粗蛋白含量采用凯氏定氮法,能量密度测定使用上海爱斯特公司生产的DM3200型氧弹热量仪。
实验数据均以平均值±标准差(Means±SE)形式表示,实验结果使用SPSS 19.0进行分析,在单因素ANOVA基础上,采用Duncan’s多重比较进行组间的差异性比对,当P<0.05时被认为具有显著性差异。
2结果与分析
2.1不同饵料组中两种规格幼蟹的能量密度差异性
表2所示,再摄食前,相同饵料料组不同规格的幼蟹能量密度均未见显著差异(P>0.05),不同饵料组相同规格的幼蟹能量密度也均未见显著差异(P>0.05)。再摄食早期,相同饵料组不同规格幼蟹,仅在鲫鱼肉组表现为大规格幼蟹显著高于小规格幼蟹(P<0.05),不同饵料组仅小规格幼蟹表现出鲫鱼肉组显著高于玉米组和饲料组(P<0.05)。临近蜕壳前,不同饵料组仅大规格幼蟹能量密度表现出玉米组显著高于鲫鱼肉组和饲料组(P<0.05),相同饵料组不同规格幼蟹能量密度均未见显著差异(P>0.05)。
除鲫鱼肉组小规格幼蟹之外,其它同饵料组中同规格幼蟹在3个时间点,均呈现先上升后下降的走势,且饲料组两种规格幼蟹均表现为再摄食早期的能量密度显著高于再摄食前和临近蜕壳前(P<0.05)。3个饵料组的同规格幼蟹在再摄食前和临近蜕壳前的能量密度均未见显著差异(P>0.05)。
2.2不同饵料组两种规格幼蟹肝胰腺和肌肉的总脂含量差异性
表3所示,对于肝胰腺中总脂含量,各个时间点相同饵料组不同规格幼蟹的肝胰腺总脂含量均未见显著差异(P>0.05);不同饵料组同规格幼蟹的肝胰腺总脂含量也均未见显著差异(P>0.05)。
对于肌肉中总脂含量,再摄食前,相同饵料组不同规格幼蟹的肌肉总脂含量,均表现为小规格幼蟹显著高于大规格幼蟹(P<0.05),不同饵料组相同规格幼蟹的肌肉总脂含量均未见显著差异(P>0.05);再摄食早期和临近蜕壳前,相同饵料组不同规格幼蟹的肌肉总脂含量均未见显著差异(P>0.05),不同饵料组相同规格幼蟹的肌肉总脂含量也均未见差异性(P>0.05)。
同饵料组中同规格幼蟹,在3个不同时间点,肝胰腺总脂含量均表现为临近蜕壳前高于再摄食前,但差异不显著(P>0.05);肌肉总脂方面,同饵料组小规格幼蟹肌肉总脂含量均呈下降走势,且再摄食前显著高于再摄食早期和临近蜕壳前(P<0.05),同饵料组大规格幼蟹肌肉总脂含量均未见显著差异(P>0.05)。
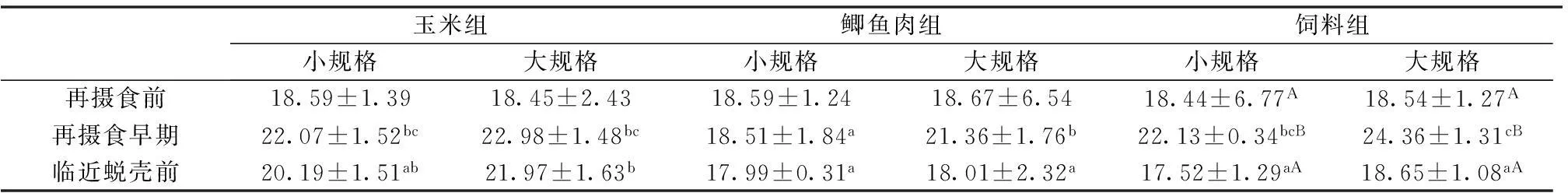
表2 不同饵料组两种规格幼蟹在3个时间点的能量密度
上标不同小写字母表示同一行差异显著(P<0.05);上标不同大写字母表示同一列差异显著(P<0.05);下同。

表3不同饵料组两种规格幼蟹在3个时间点的肝胰腺及肌肉总脂含量
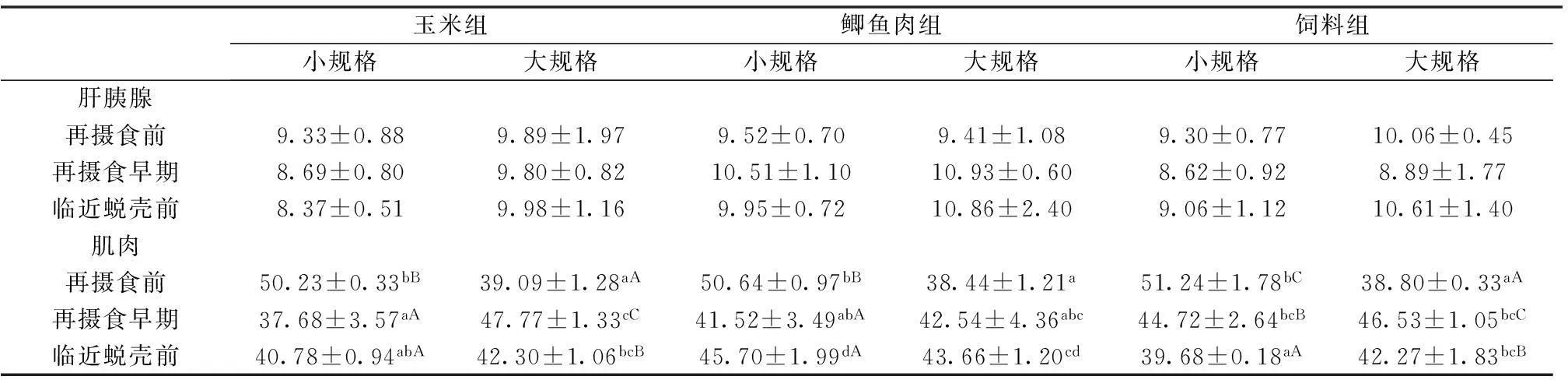
表4 不同饵料组两种规格幼蟹在3个时间点的肝胰腺及肌肉粗蛋白含量
2.3不同饵料组中两种规格幼蟹肝胰腺和肌肉的粗蛋白含量差异性
表4所示,对于肝胰腺中粗蛋白含量,各个时间点相同饵料组不同规格幼蟹均未见显著差异(P>0.05);不同饵料组相同规格幼蟹也均未见显著差异(P>0.05)。对于肌肉中粗蛋白含量,再摄食前,各个饵料组均表现为小规格幼蟹显著高于大规格幼蟹(P<0.05),不同饵料组同规格幼蟹均未见显著差异(P>0.05);再摄食早期,玉米组大规格幼蟹肌肉粗蛋白含量显著高于小规格幼蟹(P<0.05);临近蜕壳前,相同饵料组不同规格幼蟹的肌肉粗蛋白含量表现为,饲料组大规格幼蟹显著高于小规格幼蟹(P<0.05),玉米组及鲫鱼肉组不同规格幼蟹均未见显著差异(P>0.05),不同饵料组相同规格幼蟹表现为,小规格幼蟹鲫鱼肉组显著高于玉米组和饲料组(P<0.05),大规格幼蟹各个饵料组之间未见显著差异(P>0.05)。
同饵料组中同规格幼蟹,在3个不同时间点,肝胰腺粗蛋白含量未见显著性差异(P>0.05);肌肉粗蛋白方面,各个饵料组中小规格幼蟹的肌肉粗蛋白含量均表现下降走势,且再摄食前显著高于再摄食早期和临近蜕壳前(P<0.05);对于大规格幼蟹,玉米组及饲料组均表现为再摄食早期>临近蜕壳前>再摄食前(P<0.05),而鲫鱼肉组表现为持续升高的走势,差异不显著(P>0.05)。
3讨论
3.1越冬后再摄食阶段幼蟹体内的营养和能量物质变化
实验结果显示,这一时期幼蟹全个体能量密度呈先上升、后下降的变化走势,并且对比再摄食前后的能量密度来看并没有显著的变化。但再摄食早期幼蟹能量密度表现出上升,表明再摄食早期幼蟹体内高能量密度的物质比低能量密度的物质积累速度更快,从而导致幼蟹个体能量密度呈上升走势。随着再摄食的继续,低能量密度的物质积累速度开始超过高能量密度的物质,导致幼蟹整体能量密度开始下降。这表明,越冬后再摄食早期,幼蟹会率先积累用于供能的物质,而在随后的摄食过程中,非主要用于供能的物质会逐渐积累起来。最终临近蜕壳前,幼蟹整体的能量密度重新回到再摄食前的水平,这说明幼蟹越冬后再摄食前后会保持营养的总体均衡。同时本实验选用3种营养组成差异显著的饵料进行饲喂,而临近蜕壳前各组幼蟹的能量密度虽有差异但不显著,可能因为幼蟹体内存在某种营养调节机制以均衡能量和营养物质的组成比例。
中华绒螯蟹体内的主要能量物质包括脂类和蛋白质,它们大量存在于肝胰腺和肌肉中,随时进行着大量复杂的物质转化和能量转移[13-15]。高能量密度的脂类主要储存在肝胰腺内,低能量密度的蛋白质主要储存在肌肉内。由本实验的结果发现,肝胰腺中总脂含量在整个再摄食阶段内虽然有所波动,但差异不显著。分析原因一方面是因为食物中脂类物质含量低,如玉米;另一方面原因可能是机体为了维持营养均衡,以及维持正常的运动,而将脂类分解和转化成其它小分子物质,如脂肪酸、氨基酸、碳水化合物等,但这并不会引起幼蟹整体能量密度仍然会升高。肌肉是个体运动的直接器官,小规格幼蟹的再摄食过程中,肌肉总脂和蛋白质的相对含量均呈下降走势,分析原因一方面表明小规格幼蟹在维持必要的活动时,仍然需要消耗肌肉组织中储备的脂类和蛋白质;另一方面是大量小分子营养和能量物质参与到肌肉运动中,导致肌肉中脂类和蛋白质相对含量下降。大规格幼蟹肌肉中的总脂保持稳定,而蛋白质含量有所升高,表明其通过脂类物质提供能量足够支持其运动所需,而蛋白质得以积累。后续工作进一步测试脂肪酸、氨基酸及碳水化合物的含量后才可探明确切原因。
3.2不同规格幼蟹在能量和营养物质积累方面的差异性
由表3和表4来看,尽管再摄食前不同规格幼蟹肌肉总脂和蛋白质含量有显著差异性,但经过再摄食至临近蜕壳前,不同规格幼蟹肌肉的总脂和蛋白质水平趋于接近,这说明不同规格幼蟹在营养的积累规律上存在差异性。再摄食的整个阶段,摄食相同饵料的大规格幼蟹能量密度始终略高于小规格幼蟹,但总体上不存在显著性差异,且二者的变化走势也相同,表明这一阶段不同规格的幼蟹储存能量物质的规律表现出一致性。
不同规格幼蟹的肝胰腺总脂和蛋白质含量未出现显著性差异,这可能与养殖中饱食投喂有关。值得注意的是,肌肉中脂类和蛋白质含量在喂食之前均表现为小规格幼蟹显著高于大规格幼蟹。分析原因,从生态学角度来讲,在极端的低温环境下,动物通过提高大分子脂类和蛋白质在机体内的所占比例,降低水分含量,从而避免被低温环境冻伤[16, 17],在洄游性鱼类脂肪含量一般较高,其对于其抵御饥饿具有重要意义[18-20];同时,对于幼蟹在越冬后的摄食竞争中,小规格幼蟹机体组织中更多地储备营养和能量物质,从而能够争取到更多的生存机会。
3.3再摄食前后不同饵料对幼蟹营养和能量积累状况的差异
中华绒螯蟹养殖生产中常用的饵料主要分3类:植物性饵料,包括玉米、豆粕和麦麸等;动物性饵料,包括野杂鱼、螺蛳等;人工配合饲料。本实验选取了玉米、鲫鱼肉和配合饲料3种不同类型饵料,比较3种饵料对越冬后再摄食幼蟹营养积累的效果。由实验中临近蜕壳前的结果显示,喂食玉米的幼蟹能量密度高于饲料组和鲫鱼肉组,且大规格幼蟹呈现显著性差异。虽然脂类物质的能量密度高于糖类,但玉米中由于富含淀粉,因此从摄入的绝对能量值分析,饱食投喂最终导致玉米组幼蟹的能量密度高于鲫鱼肉组和饲料组。同时,玉米组两种规格幼蟹肝胰腺总脂含量均高于其它两组,但差异不显著。王耀光[21]认为中华绒螯蟹属杂食性动物,对淀粉有很好的消化能力。从食物中获得一定量的淀粉作为能量源可以有效节约脂类物质[22-24],并被储存在肝胰腺中,而肝胰腺中蛋白含量低且稳定,因而导致玉米组幼蟹肝胰腺总脂含量高于其它两组。值得注意的是,饲料组幼蟹再摄食早期能量密度显著升高,而在之后则显著下降,这表明饲料对于幼蟹再摄食早期快速积累能量物质,以及后期快速积累非主要供能物质的效果均要好于玉米和鲫鱼肉,这或许与饲料合理的营养配比有关。
杨霞等[25]研究结果显示,饲料中用棉籽粕替代鱼粉的含量高于一定量时,幼蟹全个体粗蛋白含量随棉籽粕含量增加而显著降低,并且肝胰腺胃蛋白酶活性也显著降低。这与本实验中玉米组得到的结果较为一致,中华绒螯蟹体内的蛋白质主要存在于肌肉中,因此玉米组中两种规格幼蟹的肌肉蛋白质含量均低于鲫鱼肉组。关于水产动物饲料中植物蛋白源的研究,很多报道均指出适量的玉米蛋白粉替代鱼粉可以保持饲料对养殖动物的脂类和蛋白质需求量[26, 27]。但如果持续投喂纯植物性饵料,抑或造成养殖动物某些营养物质不足,从而影响生长发育[28, 29]。而鲫鱼肉组幼蟹最终肌肉蛋白含量为3组中最高,表明动物性饵料在幼蟹蛋白质积累方面具有优势。
关于越冬后至第一次蜕壳前这一时期,中华绒螯蟹的养殖是否需要投饵的问题尚无相关报道。养殖经验指出,即使不投喂饵料,幼蟹也可以完成越冬后的前一到二次蜕壳,不会影响最终的产量。考虑到在池塘养殖中,在即便不投喂饵料的情况下,幼蟹仍然可以从池塘底质中获取食物,因此本实验在循环水养殖模式下并未设立不喂食组。本实验初步探究了这一时期幼蟹经过再摄食后,机体能量和营养物质的积累规律,为越冬后投喂何种类型的饵料提供参考。但尚未涉及喂食与不喂食的区别,以及喂食对于越冬后的第一次蜕壳成活率。以上两个问题对于从根本上探明河蟹养殖在越冬后是否应该投饵具有关键意义,仍有待进一步研究。
参考文献:
[1]王武,王成辉,马旭洲. 河蟹生态养殖(第二版)[M].北京:中国农业出版社,2013.
[2]马明君,王春,吴旭干,等. 中华绒螯蟹幼蟹蜕皮周期中肝胰腺和肌肉脂类的动态变化[J]. 水产学报, 2014, 38(11):1889-1898.
[3]林仕梅,罗莉,叶元土. 中华绒螯蟹的营养研究进展[J]. 饲料工业, 2011, 22(08): 15-17.
[4]楼伟风,李爱杰,徐家敏. 中国对虾(PenaeusorientalisKishiouys)粗蛋白、氨基酸含量的比较分析[J]. 青岛海洋大学学报:自然科学版, 1989, 19(52): 69-79.
[5]Wu X G, Cheng Y X, Sui L Y, et al. Effect of dietary supplementation of phospholipid and highly unsaturated fatty acids on reproductive perfrmance and offspring quality of the Chinese mitten crab(Eriocheirsinensis) female broodstock[J]. Aquaculture, 2007, 273(4): 602-613.
[6]Wu X G, Cheng Y X, Sui L X, et al. Biochemical composition of pond-reared and lake-stocked Chinese mitten crabEriocheirsinensis[J]. Aquaculture Research, 2007, 38(4): 1459-1467.
[7]成永旭,堵南山,赖伟. 不同阶段中华绒螯蟹肝胰腺的脂类及脂肪酸组成[J]. 动物学报, 1998, 44(4):420-429.
[8]陈炳良,堵南山. 中华绒螯蟹的食性分析[J]. 水产科技情报, 1989(1):2-5.
[9]纪连元,杨志刚,阙有清,等. 饵料对雌中华绒螯蟹可食部分含量和体成分的影响[J]. 上海海洋大学学报,2012, 21(4):561-567.
[10]Luo Z, Tan X Y, Chen Y D, et al. Apparent digestibility coefficients of selected feed ingredients for Chinese mitten crab,Eriocheirsinensis[J]. Aquaculture, 2008, 285(1-4):141-145.
[11]Kucharski L C R, Dasilvar R S M. Effect of diet composition on the carbohydrate and lipid metabolism in an estuarine carb,Chasmagnathusgranulata[J]. Comparative Biochemistry and Physiology, 1991, 99(1-2):215-218.
[12]Folch J, Lees M, Stanley G H S. A simple method for the isolation and purification of total lipids from animal tissues[J]. Journal of Biological Chemistry, 1957, 226:497-509.
[13]Holland D L. Lipid reserves and energy metabolism in the larvae of benthic marine invertebrates[J]. Biochemical and Biophysical Perspectives in Marine Biology, 1978, 4:85-123.
[14]MIKAMI S, Greenwood J G, Takashima F. Functional morphology and cytology of the phyllosomal digestive system ofSivacusciliatusandPanulirusjaponicus(Decapoda, Scyllaridae and Palinuridae)[J]. Crustaceana, 1994, 67(2):212-225.
[15]Al-Mohanna S Y, Nott J A, Lane D J W. Mitotic E and secretory F-cells in the hepatopancreas of shrimpPenaerssemisulcatus[J]. Journal of the Marine Biological Association of the United Kingdom, 1985, 65:901-910.
[16]常杰,葛滢.生态学[M]. 北京:高等教育出版社, 2010.
[17]孙儒泳,李庆芬,牛翠娟,等. 基础生态学[M]. 北京:高等教育出版社, 2002.
[18]Xie X J, Long T C, Zhang Y G, et al. Reproduction investiment in the southern catfish (Silurusmetidionalis)[J]. J Fish Biol, 1988, 52:259-271.
[19]Hutchings J A, Pickle A, McGregor-Shaw C R, et al. Influence of sex,body size,and reproduction on over winter lipid depletion in brook trout[J]. J Fish Biol, 1999, 55(5): 1020-1028.
[20]Simpkins D G, Hubert W A, Martinez Del Rio C, et al. Constraints of body size and swimming velocity on the ability of juvenile rainbow trout to endure periods without food[J]. J Fish Biol, 2004, 65(2): 530-544.
[21]王耀华,解绶启,朱晓鸣,等. 饲料中不同淀粉水平对中华绒螯蟹幼蟹生长、饲料利用的影响[J]. 水生生物学报,2012,36(6):1011-1017.
[22]江星,陈立侨,孙盛明,等. 中华绒螯蟹对10种常见饲料蛋白源的表观消化率[J]. 海洋渔业,2013,35(2):209-216.
[23]陈立侨,堵南山,赖伟. 中华绒螯蟹对蛋白质和脂肪消化率的初步研究[J]. 水产养殖,1993(6):15-18.
[24]朱晓鸣,崔奕波,光寿红. 中华绒螯蟹对三种天然饵料的选食性及消化率[J]. 水生生物学报,1997,21(1):94-97.
[25]杨霞,叶金云,张易祥,等. 普通棉籽粕和发酵棉籽粕替代鱼粉对中华绒螯蟹幼蟹生长性能、体成分及肝胰腺消化酶活性的影响[J]. 动物营养学报,2014,26(3):683-693.
[26]Regost C, Arzel J, Kaushik S J. Partial or total replacement of fish meal by corn gluten meal in diet for turbot(Psettamaxima)[J]. Aquaculture, 1999, 180(1-2):99-117.
[27]程媛媛,周洪琪,华雪铭,等. 玉米蛋白粉部分替代鱼粉对罗氏沼虾生长、氨基酸沉积率和肌肉营养成分的影响[J]. 中国水产科学,2009,16(4):572-579.
[28]陈立侨,堵南山. 中华绒螯蟹蟹种配饵中豆饼替代部分鱼粉的适宜含量[J]. 水产学报, 1994, 18(1):24-31.
[29]李旭光,周刚,张彤晴,等. 不同饵料饲喂对中华绒螯蟹一般营养成分的影响[J]. 江苏农业科学, 2011, 39(4):410-412.
中图分类号S963.2; S966.16
文献标识码A
文章编号2095-1736(2016)02-0076-05
作者简介:王福辰,硕士,研究方向为中华绒螯蟹养殖与营养生化,E-mail:fortunatella@yeah.net;通信作者:成永旭,博士,教授,研究方向为甲壳动物营养繁殖学,E-mail:yxcheng@shou.edu.cn。
基金项目:国家“八六三”高技术研究发展计划(2012AA10A409-5);国家农业科技成果转化项目(2012GB2C000147);上海市科委科研计划(13340721500,13320502100);上海市科委优秀学术带头人项目(12XD1402700)
收稿日期:2015-06-23;修回日期:2015-07-31
doi∶10.3969/j.issn.2095-1736.2016.02.076
