西北太平洋秋刀鱼耳石透明带的初步研究
2016-03-04朱清澄高玉珍花传祥杨明树
朱清澄,高玉珍,花传祥,杨明树
(1.上海海洋大学海洋科学学院,上海 201306;
2.国家远洋渔业工程技术研究中心,上海海洋大学,上海 201306;
3.大洋渔业资源可持续开发省部共建教育部重点实验室,上海海洋大学,上海 201306)
西北太平洋秋刀鱼耳石透明带的初步研究
朱清澄1,2,3,高玉珍1,花传祥1,2,3,杨明树1
(1.上海海洋大学海洋科学学院,上海 201306;
2.国家远洋渔业工程技术研究中心,上海海洋大学,上海 201306;
3.大洋渔业资源可持续开发省部共建教育部重点实验室,上海海洋大学,上海 201306)
根据2014年我国秋刀鱼捕捞船在西北太平洋采集的秋刀鱼(Cololabis saira)样本,提取1 062个耳石进行拍照处理,并根据耳石透明带特征对其进行划分,运用统计学方法研究耳石类型及与肉体长组的关系,分析各月份耳石类型变化。研究发现,83%小型鱼(KnL≤240 mm)和71%中型鱼(240 mm<KnL≤280 mm)耳石属于类型Ⅰ,大型鱼(280 mm<KnL≤314 mm)和特大型鱼(KnL>314 mm)耳石类型Ⅱ、类型Ⅲ占优。肉体长在280 mm以下的类型Ⅰ耳石比例较大,耳石没有形成透明带,随着KnL的增加,类型Ⅱ和类型Ⅲ在肉体长组的比例增加。耳石透明带最早在202 mm时开始形成,秋刀鱼耳石第1条透明带主要在260~280 mm之间形成,以中型鱼为主;秋刀鱼耳石第2条透明带主要在290~310mm之间形成,以大型鱼为主。雌性和雄性秋刀鱼耳石在各耳石类型下无显著差别(P=0.064>0.05)。耳石类型Ⅰ比例在月份间变化大,在10月和11月份耳石类型Ⅰ比例占优,分别为44.1%和36.4%,多为小型和中型鱼,6月和7月份耳石类型Ⅱ和类型Ⅲ比例大约为40%,多数为大型和特大型鱼,表明大型鱼比小型和中型鱼早到达索饵场进行索饵。
秋刀鱼;耳石类型;透明带
秋刀鱼(Cololabis saira)属于颌针鱼目(Beloniformes)竹刀鱼科(Scomeresoide)[1-2],是一种经济性较高的鱼类[3],主要分布于北太平洋[4],主要捕捞国家和地区有日本、俄罗斯、韩国、中国台湾省和中国大陆[5-7]。
秋刀鱼在5~7月份从亚热带地区向北迁移,到达亲潮海域(Oyashio waters)进行索饵,然后在7~8月份间向南洄游,到达黑潮海域(Kuroshio waters)进行产卵,产卵时间是从9月份到次年的6月份[8-10],秋刀鱼寿命一般是 2龄[11]。
秋刀鱼耳石透明带是在秋天和冬天形成[11],并用来判定0龄和1龄秋刀鱼[12]。秋刀鱼耳石轮纹每天形成1条[13],耳石日轮在耳石透明域中无法判读,很难推算秋刀鱼的孵化日期[14]和研究秋刀鱼的年龄结构,因此利用耳石透明带来研究秋刀鱼年龄结构,对其资源进行评估。早在60年代,堀田秀之[15]对小型群、中型群、大型群和特大型群秋刀鱼耳石透明带进行研究,简单概述小型、中型和大型秋刀鱼耳石透明带的特征,到90年代,一些学者根据透明带划分秋刀鱼耳石类型,将其应用在秋刀鱼年龄结构和洄游路线的研究上。本文根据已有的秋刀鱼耳石划分方法,初步研究西北太平洋秋刀鱼耳石透明带类型及与肉体长组的关系,分析透明带形成时的体长和耳石类型性别间差异,比较各月份耳石类型,为以后研究秋刀鱼种群结构、年龄结构、洄游路线提供基础信息。
1 材料与方法
1.1 样本采集
样本采集委托秋刀鱼捕捞渔船“鲁蓬远渔027”和“国际908”进行,调查时间分别为2014年6~7月和10~11月,调查采样海域分别为41°31′N~44°31′N、157°15′E~162°26′E;38°38′N~44°24′N、146°38′E~155°44′E(图1)。采集站点的样本从渔获物中随机获得,采集的样本经冷冻保藏运回实验室。
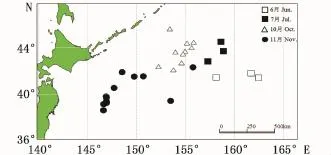
图1 秋刀鱼采样站点图Fig.1 Sam pling locations of C.saira
1.2 研究方法
1.2.1 基础生物学测定
在实验室经解冻后对秋刀鱼进行生物学测定,包括肉体长(knob length,KnL)、体质量(body weight,BW)、性别及性腺成熟度等,KnL是下颚前端至尾鳍银白色肉质末端的距离,其测量的标准偏差最小[16]。KnL和叉长的测定精度为0.1 cm,体质量测定精度为0.1 g,性腺成熟度划分方法参照《海洋水产资源调查手册》[17]。根据秋刀鱼肉体长将其分为4种类型[18,19],240 mm以下为小型鱼(KnL≤240 mm),240~280 mm为中型鱼(240<KnL≤280 mm),280~314 mm为大型鱼(280<KnL≤314 mm),314 mm以上为特大型鱼(KnL>314 mm)。
1.2.2 耳石提取与处理
从秋刀鱼头部平衡囊取出耳石,将取出的耳石放于有95%乙醇溶液的1.5 mL的离心管中并进行编号,获得2 800对耳石,随机抽取1 046个样本,其中秋刀鱼KnL范围194~343 mm,体质量范围29~192 g。将左耳石凸面向上置于Olympus光学显微镜×40倍下用CCD拍照。测量工作由2人同时进行,若两者误差小于5%,取两者的平均值;若两人测量误差大于5%,则重新对其测量。
1.2.3 耳石类型划分方法
按照巣山哲[18]法将耳石划分为以下5种类型:
类型Ⅰ:耳石边缘没有透明带(图2-A),或者边缘有狭窄的、不完全的透明域(图2-B)。
类型Ⅱ:耳石边缘有透明带,耳石中心附近的不透明部分外侧有宽的、完全的透明域(图2-C)。类型Ⅲ:耳石边缘有1条完整透明带,完整透明带外侧有不透明部分(图2-D,E)有些耳石具有完整透明带,其边缘部分有窄的、不完全的透明域(图2-E)。类型Ⅳ:耳石边缘有2条完整透明带图(2-F)。其它类型:不属于以上4种类型的耳石,有些耳石边缘部分有3条透明带(图2-G),有些耳石整体透明(图2-H)。
1.3 数据处理
利用Photoshop对耳石图片进行处理,划分耳石类型;利用Excel进行生物学数据处理。
2. 结果与分析
2.1 耳石类型
2.1.1 耳石类型统计
在1 062个耳石样本中,类型Ⅰ、类型Ⅱ、类型Ⅲ耳石样本数量占优势,分别为326、324、317个样本,具有两条透明带的秋刀鱼样本较少(表1)。小型(KnL≤240 mm)和中型(241<KnL≤280 mm)秋刀鱼耳石类型Ⅰ比重大,分别占83%、71%,大型(280<KnL≤314 mm)和特大型(KnL>314 mm)秋刀鱼耳石类型Ⅱ分别为37.5%和35%,类型Ⅲ比例分别为42.3%和42.8%。因此,秋刀鱼耳石不超过1条透明带比例占91.1%,大约74%的小型和中型秋刀鱼耳石边缘透明带还未形成,大型和特大型秋刀鱼耳石边缘有透明带比例分别为83.1%和和83.8%。
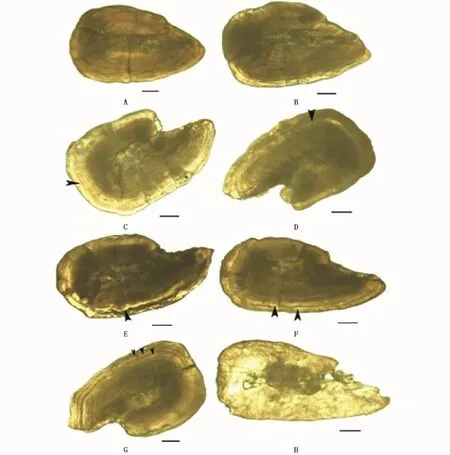
图2 秋刀鱼耳石形态示意图Fig.2 Statolith morphology figure of C saira
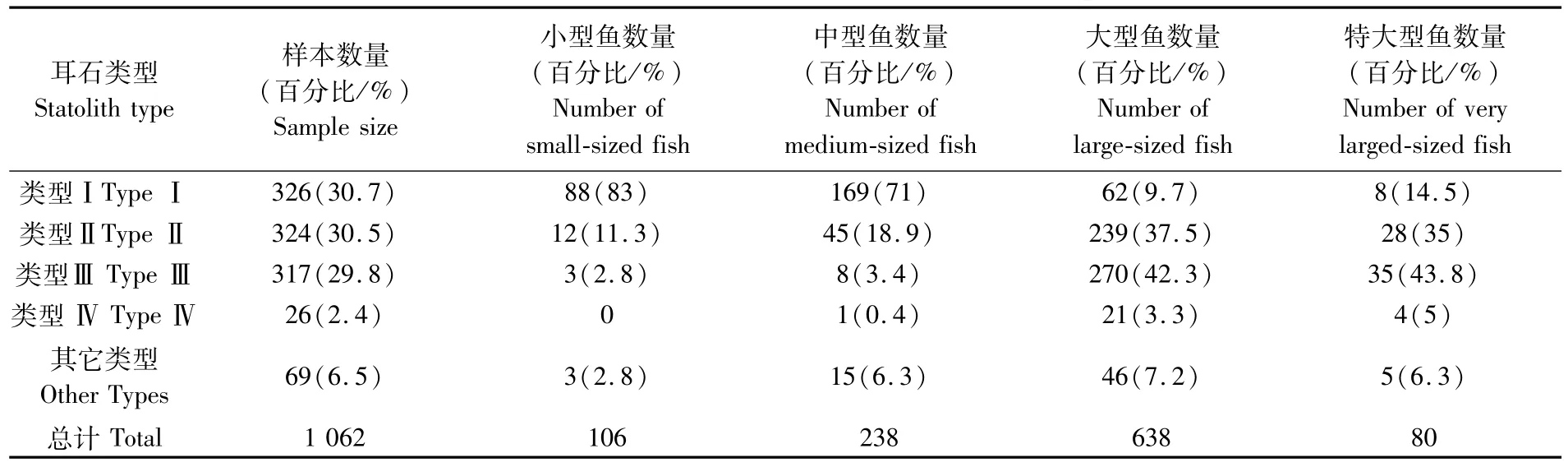
表1 秋刀鱼耳石类型统计分析Tab.1 Statistic analysis of C.saira statolith type
2.1.2 耳石类型与肉体长组的关系
结果显示(图3),测得KnL≤200 mm的个体为4 ind,其耳石类型为Ⅰ型,类型Ⅰ在肉体长组(200~220 mm、220~240 mm、240~260 mm、260~280 mm)的比例分别为82.1%、82.8%、74.1%、69.3%,在KnL≤280 mm比例较大,肉体长组240~260 mm之间耳石出现两条透明带。随着KnL的增加,类型Ⅱ和类型Ⅲ在每组肉体长组的比例增加,肉体长组280~300 mm、300~320 mm、320~340 mm的耳石类型Ⅱ和类型Ⅲ比例占优。因此,240~260 mm之间耳石出现两条透明带,280 mm以下耳石类型Ⅰ占优,耳石没有形成透明带,280 mm以上的耳石类型Ⅱ和类型Ⅲ比例增加。

图3 不同肉体长组的秋刀鱼耳石类型百分比Fig.3 Percentage of statolith type in different size classes of C.saira
2.1.3 耳石透明带形成时的体长
研究结果显示,耳石透明带最早在202 mm时开始形成(图4),第1条透明带形成时的肉体长范围为200~320 mm,其中小型、中型和大型秋刀鱼比例分别为15.2%、54.5%、29.8%。正在形成第1条透明带的耳石数量在260~280 mm之间出现峰值(图4-A),说明秋刀鱼耳石第1条透明带主要在260~280 mm之间形成,以中型鱼为主。第2条透明带形成时的肉体长范围为260~330 mm,最初的肉体长范围260~265 mm,正在形成第2条透明带的耳石数量在290~310 mm之间出现峰值,占69.6%,说明秋刀鱼耳石透明带主要在290~310 mm之间形成,以大型鱼为主。
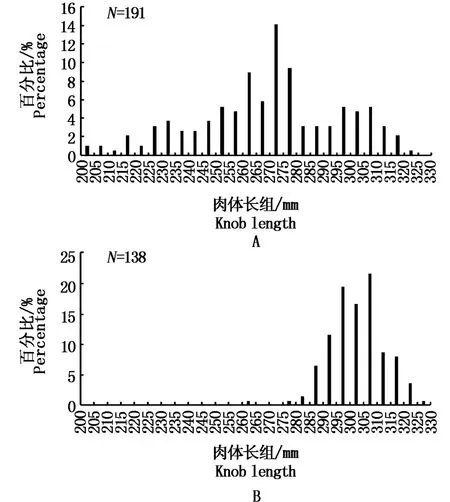
图4 秋刀鱼耳石透明带形成时的肉体长Fig.4 Knob length at the time of formation of hyaline zones in the Pacific saury
2.2 耳石类型性别差异
研究表明,雌性秋刀鱼KnL范围为195~329 mm,雌性秋刀鱼KnL范围为206~343 mm,雌雄比为1.7∶1,无法判别的秋刀鱼为146 ind,雌性和雄性秋刀鱼耳石各类型无差别(卡方值=8.874,P=0.064>0.05),雌性秋刀鱼耳石无透明带比例占27.1%,有透明带比例为65.2%,雄性无透明带耳石比例占24%,有透明带的耳石比例为70.6%,雌性和雄性秋刀鱼耳石具有透明带的比例占优,性别无法判读的秋刀鱼耳石类型I占60.3%,其耳石无透明带(表2)。从图5中看出,雌性和雄性秋刀鱼(<290 mm)耳石以类型I为主,290 mm以上以类型Ⅱ和类型Ⅲ为主。
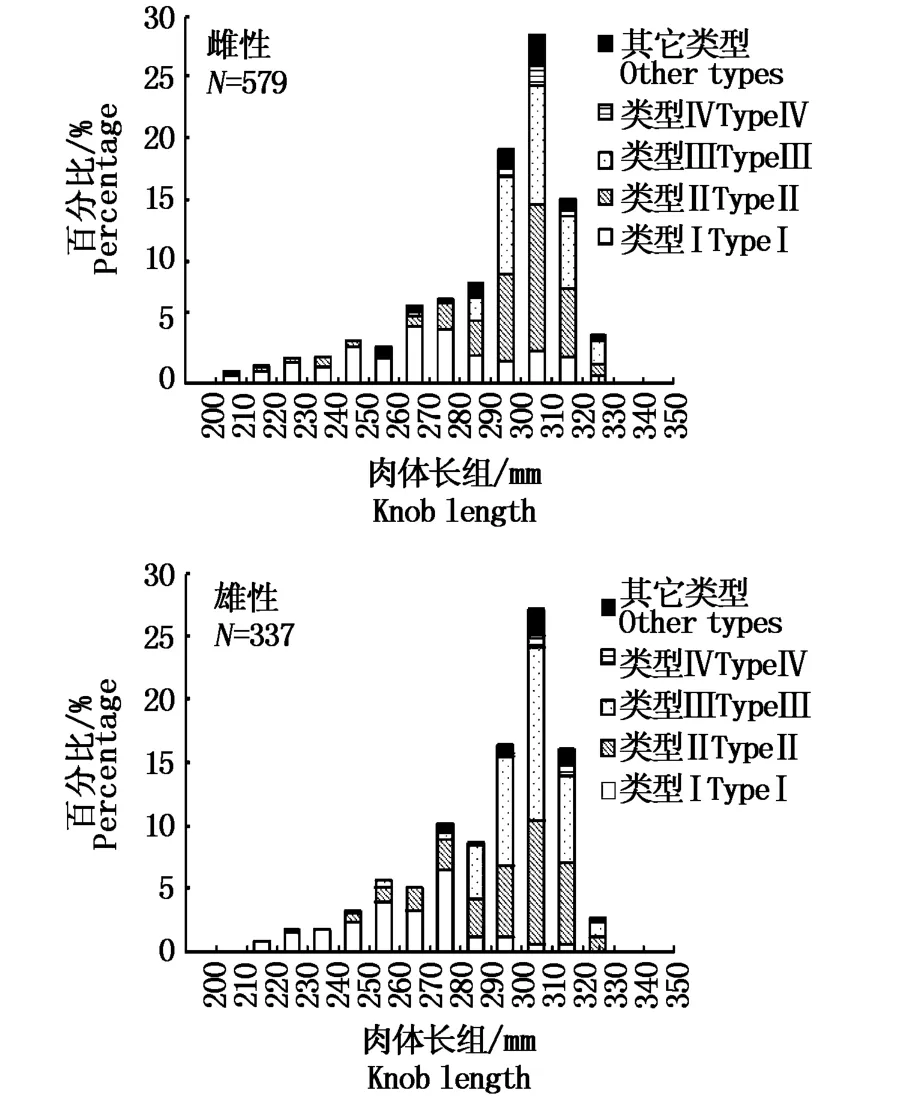
图5 不同性别秋刀鱼耳石类型分析Tab.5 Analysis of C.saira statolith type in different sexes
2.3 耳石类型月份变化
从图6中看出,耳石类型Ⅰ比例月份间变化大,6月和7月耳石类型Ⅰ比例小,而10月和11月耳石类型Ⅰ比例占优,分别为44.1%和36.4%,尤其是10月下旬和11月上旬,在此期间,还未形成透明带的个体较多,多为小型和中型秋刀鱼(图7)。6月和7月份耳石类型Ⅱ和类型Ⅲ比例大约在40%,多数为大型和特大型秋刀鱼,10月下旬和11月上旬比例较低,到11月中旬,其比例增加。具有两条透明带的个体数量较少,月份间比例变化不大。因此,6月和7月份秋刀鱼耳石有透明带的比例较高,其中KnL≥280 mm的秋刀鱼占优,而且6、7月是秋刀鱼北上洄游进行索饵的时间,所以大型比小型和中型秋刀鱼早到达索饵场进行索饵。
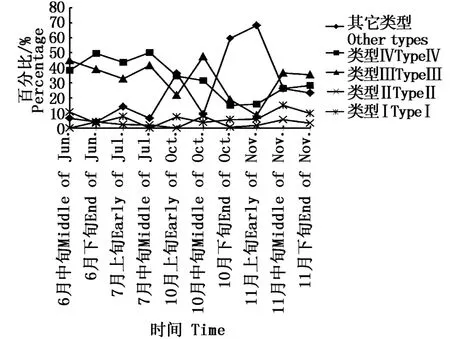
图6 不同月份间秋刀鱼耳石类型的变化Fig.6 Variation of statolith type of Pacific saury between differentm onths
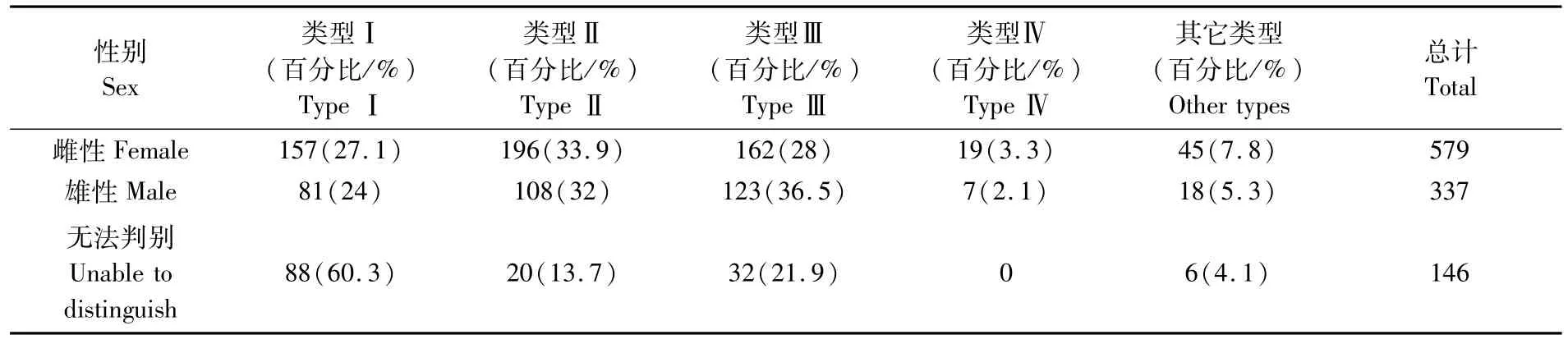
表2 秋刀鱼耳石类型性别统计Tab.2 Statistic of statolith type of C.saira in sex
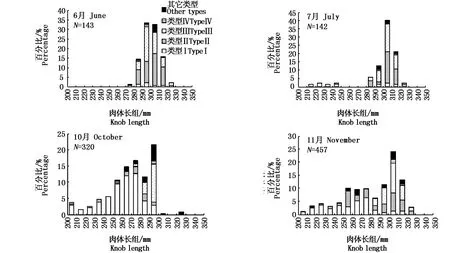
图7 耳石类型与肉体长组月份间变化Fig.7 M onthly changes in size composition and statolith type
3 讨论
3.1 耳石类型
本研究发现,83%小型(KnL≤240 mm)和71%中型秋刀鱼(240<KnL≤280 mm)耳石属于类型Ⅰ,大型(280<KnL≤314 mm)秋刀鱼耳石类型Ⅱ、类型Ⅲ比例分别为37.5%、42.3%;类型Ⅰ在体长组(200~220 mm、220~240 mm、240~260 mm、260~280 mm)的比例较大,体长组280~300 mm、300~320 mm、320~340 mm的耳石类型Ⅱ和类型Ⅲ比例占优,说明小型和中型秋刀鱼耳石属于类型Ⅰ,并其耳石没有透明带,大型秋刀鱼耳石属于类型Ⅱ和类型Ⅲ,形成1条透明带。巣山哲[18]发现大约80%小型和中型秋刀鱼(KnL≤280 mm)耳石属于类型Ⅰ,这与本研究一致,而15.8%和65.4%的大型秋刀鱼(>280 mm)分别属于类型Ⅱ和类型Ⅲ,这与本研究结果有差异,可能原因是样品采集位置和采集年份,不同地理位置耳石类型Ⅱ和类型Ⅲ数量也有差异[20],而且每年的秋刀鱼数量和大小波动较大[19],可能使得本研究中2014年大型鱼耳石类型Ⅱ、类型Ⅲ比例较小。
本研究结果表明,耳石透明带最早在202mm时开始形成,第1条透明带形成时的肉体长范围为200~320 mm,其中中型鱼比例占54.5%,说明秋刀鱼耳石第1条透明带群体以中型为主,第2条透明带形成时的肉体长范围为260~330 mm,以大型为主,这与巣山哲[21]研究相似。240~260 mm秋刀鱼耳石出现2条透明带(类型Ⅳ),可能是秋刀鱼从孵化到秋冬季形成透明带之前生长缓慢,使得较早形成第1条透明带,而第2条透明带形成时间还未确定,可能在其它季节形成[11]。
3.2 耳石类型性别差异
在本研究中,雌性秋刀鱼耳石无透明带比例占27.1%,有透明带比例为65.2%,雄性无透明带耳石比例占24%,有透明带的耳石比例为70.6%,说明雌性和雄性秋刀鱼耳石有透明带比例占优。雌雄性别比为1.7∶1,结合之前研究发现,秋刀鱼雌雄性别比接近 1∶1,最大达到1.42∶1[7,22],这可能与采样时间和采样地点、补充群体数量有关。雌性和雄性秋刀鱼(<290 mm)耳石以类型Ⅰ为主,290 mm以上以类型Ⅱ和类型Ⅲ为主,说明雌性和雄性秋刀鱼耳石类型在各肉体长组分布相类似。
3.3 耳石类型月份间变化
本研究发现,耳石类型I比例月份间变化大,10月和11月耳石类型Ⅰ比例占优,还未形成透明带的个体较多,多为小型和中型秋刀鱼。6月和7月份耳石类型Ⅱ和类型Ⅲ比例大约在40%,多数为大型和特大型,这与巣山哲[18]研究结果类似。秋刀鱼在5~7月份北上洄游进行索饵[23],因此大型比小型和中型秋刀鱼早到达索饵场进行索饵,使得6月和7月份秋刀鱼耳石有透明带居多。10月和11月秋刀鱼耳石类型Ⅰ比例占优,耳石样本几乎没有透明带,秋刀鱼耳石透明带在秋、冬季形成[24],说明10月和11月秋刀鱼样本为2014年春生和秋生群体。
致谢:本研究得到国家远洋渔业工程技术研究中心和大洋渔业资源可持续开发教育部重点实验室的资助,感谢在海上调查的同学,使得本研究得以顺利进行。
[1] 汤振明,黄洪亮,石建高.中国开发利用西北太平洋秋刀鱼资源的探讨[J].海洋科学,2004,28(10):56.
TANG ZM,HUANG H L,SHI JG.An approach to exploitation and utilization ofCololabis sairaresource in west Pacific Ocean[J].Marine Science,2004,28(10):56.
[2] 商李磊,朱清澄,晏 磊,等.西北太平洋公海秋刀鱼生物学特性研究[J].江苏农学,2012,40(10):198-201.
SHANG L L,ZHU Q C,YAN L,et al.Study on biological characteristics of Cololabis saira in the Northwest Pacific Ocean[J].Jiangsu Agricultural Science,2012,40(10):198-201.
[3] NAKAYA M,MORIOKA T,FUKUNAGA K,et al.Growth and maturation of Pacific sauryCololabis sairaunder laboratory conditions[J].Fisheries Science,2010(76):45-53.
[4] SUYAMA S,OSHIMA K,NAKAGAMIM,et al.Seasonal changes in statolith and somatic growth in age-0 Pacific saury Cololabis saira[J].Fisheries Science,2011(77):223-233.
[5] 沈建华,韩士鑫,樊 伟,等.西北太平洋秋刀鱼资源及其渔场[J].海洋渔业,2004(1):61-64.
SHEN J H,HAN S X,FAN W,et al.Resource and Fishing Grounds in the Northwest Pacific.Marine Fisheries,2004(1):61-64.
[6] 张 阳,朱清澄,晏 磊,等.西北太平洋公海春季秋刀鱼生物学特性的初步研究[J].海洋湖沼通报,2013(1):53-60.
ZHANG Y,ZHU Q C,YAN L,et al.Preliminary study on biological characteristics ofCololabis sairain the Northwest Pacific Ocean in spring[J].Transactions of Oceanology and Limnology,2013(1):53-60.
[7] 王晓杰,朱清澄,张 阳,等.西北太平洋秋刀鱼耳石形态学分析[J].江苏农业科学,2013,41(4):210-213.
WANG X J,ZHU Q C,ZHANG Y,et al.Morphologic features of otolithCololabis sairain the northwest Pacific Ocean[J].Jiangsu Agricultural Science,2013,41(4):210-213.
[8] WATANSBE Y,CHLO N.Larval production and mortality of Pacific saury,Cololabis saira,in the Northwestern Pacific Ocean[J].Fish Bulletin,1989(87):601-613.
[9] FUKUSHIM S.Synoptic analysis of migration and fishing conditions of pacific saury in the northwest Pacific Ocean[J].Tohoku Regions Fish Research Laboratory,1979(41):1-70.
[10] OOZEKIY,WATANABE Y.Comparison of somatic growth and statolith increment growthin laboratoryreared larvae of Pacific saury,Cololabis saira,under different temperature conditions[J].Marine Biology,2000(136):349-359.
[11] SUYAMA S,KURITA Y,UENOY.Age structure of Pacific sauryCololabis sairabased on observations of the hyaline zones in the statolith and length frequency distributions[J].Fisheries Science,2006(72):742-749.
[12] SUYAMA S,OSHIMA K,NAKAGAMIM,et al.Seasonal change in the relationship between statolith radius and body length in age-zero Pacific sauryCololabis saira[J].Fisheries Science,2009(75):325-333.
[13] WATANABE Y,KUJIY.Verification of Daily Growth Increment Formation in Saury Statoliths by Rearing Larvae from Hatching[J].Japanese Journal of Ichthyology,1991,38(1):11-15.
[14] SUYAMA S,NAKAGAM M.Comparison of the growth of age-1 Pacific saury Cololabis saira in the Western and the Central North Pacific[J].Fisheries Science,2012(78):277-285.
[15] 堀田秀之.鱗·耳石によるサンマのポピュレーション構造の分析とその成長[R].東北区水產研究所研究報告,1960(16):41-64.
HOTTA H.On the analysis of the population of thesaury(Cololabis saira)based on the scales of and the otolith characters,and their growth[R].Bulletin of Fish Research Laboratory of Tohoku Regions,1960(16):41-64.
[16] 木村喜之助.標準体長として測るべき魚体の部位に就いて[R].東北区水產研究所研究報告,1956(7):1-11.
KIMURA K.The standard length of the Pacific saury,Cololabis saira(Brevoort)[R].Bulletin of Fish Research Laboratory of Tohoku Regions,1956(7):1-11.
[17] 黄海水产研究所.海洋水产资源调查手册(第二版)[M].上海:科学技术出版社,1981.
Yellow sea fisheries research institute.Marine fisheries resources investigation handbook(second edition)[M].Shanghai:Press of Science and Technology,1981.
[18] 巣山哲,桜井泰憲.西部北太平洋におけるサンマの耳石透明帶の形成時期[R],東北区水產研究所研究報告,2000(63):97-108.
SUYAMA S,SAKURAI Y.Formation Period of the otolith hyaline zones of Pacific SauryCololabis saira(Brevoort)in the western North Pacific Ocean[R].Bulletin of Fish Research Laboratory of Tohoku Regions,2000(63):97-108.
[19] SUYAMA S,SAKURAI Y,SHIMAZAKI K,et al.Age and Growth of Pacific SauryCololabis saira(Brevoort)in the Western North Pacific Ocean Estimated from Daily Statolith Growth Increments[J].Fisheries Science,1996,62(1):1-7.
[20] SUYAMA S,NAKAGAM M,NAYA M,et al.Migration route of Pacific sauryCololabis sairainferred from the statolith hyaline zone[J],Fisheries Science,2012(78):1179-1186.
[21] 巣山哲.北太平洋におけるサンマCololabis saira(Brevoort)の年齢、成長および成熟に関する研究[R].横滨:水產総合研究センター研究報告,2002(5):68-113.
SUYAMA S.Study on the age,growth,and maturation process of Pacific saury,Cololabis saira(Brevoort)in the Noth Pacific[R].Bulletin of Fish Research Regions,2002(5):68-113.
[22] 叶旭昌,刘 瑜,朱清澄,等.北太平洋公海秋刀鱼生物学特性初步研究[J].上海水产大学学报,2007,16(3):264-268.
YE X C,LIU Y,ZHU Q C.Biological characteristics ofCololabis sairain North Pacific high sea[J].Journal of Shanghai Fisheries University,2007,16(3):264-268.
[23] SUNADA J S,Age and growth of Pacific saury,Cololabis saira[J].Fish Game,1974,60:64-73.
[24] HATANAKA M.Biological studies on the population of the saury,Cololabis saira(Breevort):Part I reproduction and growth[J].Tohoku of Journal of Agricultural Research,1956(6):227-269.
On statolith hyaline zones of Pacific saury Cololabis saira in the Northwest Pacific
ZHU Qing-cheng1,2,3,GAO Yu-zhen1,HUA Chuan-xiang1,2,3,YANG Ming-shu1
(1.College of Marine Sciences,ShanghaiOcean University,Shanghai201306,China;2.National Engineering Research Centre for Oceanic Fisheries,Shanghai201306,China;3.Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources,Ministry of Education,Shanghai201306,China)
Based on samples collected by Chinese saury fishery fleets from the Northwest Pacific in 2014,1 062 specimens were extracted and measured,and their statolith types were divided according to hyaline zones of statolith.The paper is to study hyaline zones of statolith and its relationship with knob length group using statistics,and to analyze the variation of statolith types in differentmonths.Results showed that83%of small-sized fish and 71%of medium-sized fish(KnL≤280 mm)exhibited typeⅠ,and in large-sized fish(280<KnL≤314 mm)and very large-sized fish(KnL>314mm)the dominantwas in typeⅡand typeⅢ,typeⅠofC.saira(KnL≤280 mm)had higher percentage,its statolith did not form hyaline zones,the proportion of typeⅡand typeⅢincreased in each group with the increasing ofKnL.Led statolith hyaline zones began to form in 202 mm,the first hyaline zones ofC.sairaformed between 260-280 mm,and medium-sized fish was dominant,the second hyaline zones ofC.sairaformed between 290-310 mm,and large-sized fish was dominant.Statolith type proportion ofmale and female Pacific saury had no difference(P=0.064>0.05).Variation of typeⅠstatolith in differentmonths is obvious.Percentage of typeⅠis dominant in October and November,and is44.1%and 36.4%respectively,moreover,it ismostly small and medium-sized fish,percentage of typeⅡand typeⅢis 40%approximately in June and July,and ismostly large-sized and very larged fish,suggesting that large-sized fish arrives in feeding ground earlier than small and medium-sized fish.
Cololabis saira;statolith type;hyaline zones
S 931.5
A
1004-2490(2016)03-0236-09
2015-08-10
国家科技支撑计划课题“大洋性渔业捕捞技术与新资源开发”(2013BAD13B05)
朱清澄(1960-),男,教授,研究方向为渔具渔法及海洋渔业管理。Tel:15692166362,E-mail:qczhu@shou.edu.cn
