2种大洋性柔鱼类角质颚形态识别法的比较
2016-03-04陈新军
苏 杭,方 舟,陈新军
(1.上海海洋大学海洋科学学院,上海 201306;2.国家远洋渔业工程技术研究中心,上海 201306;3.大洋渔业资源可持续开发省部共建教育部重点实验室,上海 201306;4.远洋渔业协同创新中心,上海 201306)
2种大洋性柔鱼类角质颚形态识别法的比较
苏 杭1,4,方 舟1,4,陈新军1,2,3,4
(1.上海海洋大学海洋科学学院,上海 201306;2.国家远洋渔业工程技术研究中心,上海 201306;3.大洋渔业资源可持续开发省部共建教育部重点实验室,上海 201306;4.远洋渔业协同创新中心,上海 201306)
依据中国鱿钓船2013年8~10月在北太平洋海域采集的柔鱼(Ommastrephes bartramii)和2014年4~7月在中东太平洋海域采集的茎柔鱼(Dosidicus gigas)样本,分别运用传统测量学法和傅里叶分析法对2种柔鱼类的角质颚进行识别,并对结果进行比较分析。运用传统测量学法共选取了12项角质颚参数值比较形态差异;角质颚外部轮廓形态通过软件转化为20组傅里叶代码(EFDs)后选取77个傅里叶值进行判别分析。结果表明,2种柔鱼类上、下角质颚形态在上头盖长(UHL)、上脊突长(UCL)、上喙长(URL)、上喙宽(URW)、上侧壁长(ULWL)、下脊突长(LCL)、下喙长(LRL)、下喙宽(LRW)、下侧壁长(LLWL)、下翼长(LWL)中存在显著性差异(P<0.01),上翼长(UWL)、下头盖长(LHL)不存在显著性差异(P>0.05)。主成分分析结果显示,传统测量法中下喙长(LRL)和下喙宽(LRW)与胴长(ML)之比主成分得分最高;傅里叶分析法显示上角质颚前16主成分可以解释总变异的83.58%;下角质颚前16主成分可以解释变异的73.76%。逐步判别分析将上头盖长(UHL)、上喙长(URL)、上侧壁长(ULWL)、下头盖长(LHL)、上喙长(URL)、上喙宽(URW)与胴长(ML)之比纳入判别分析函数,总别正确率为95%;傅里叶分析法将18个傅里叶值纳入判别函数,上颚总判别率为97.6%;下颚总判别率为85.7%。总体而言,2种方法对柔鱼类角质颚判别均有效,且上颚区分效果更明显。本文为头足类的种类判别提供了更多的且有效的识别方法。
角质颚;传统测量学法;傅里叶分析法;柔鱼;茎柔鱼;判别分析
头足类(Cephalopoda)属于软体动物门(Mollusca),种类繁多,目前已鉴别的约有700多种。头足类一般为一年生,属于短周期生命物种[1],广泛分布于三大洋,是目前国际上重点关注的海洋生物种类之一。自上世纪90年代以来,大洋性头足类已成为我国主要的远洋渔业捕捞对象,产量呈逐年上升趋势[2-4]。头足类种类结构复杂,其种类划分问题引起了国内外学者的关注[5-7]。头足类的硬组织,如耳石、内壳、角质颚等都具有形态稳定、不易腐蚀的特点,通常被应用于种类鉴别的研究中[8-10]。早期学者依据胴长组成和寄生虫感染情况进行了不同群体的划分[11-13],20世纪90年代初期BOOKSTEIN[14]和ROHLF[15-16]以数学形态学为基础创立了几何形态测量学法,该方法比起传统径向测量法更为直观和准确,同时因其在形态结构和数据分析上有重大突破,被誉为形态测量的革命[17-19]。其中外部形态法运用傅里叶分析将二维图像转化为傅里叶系数,结合统计学进行分析判别,目前已在鱼类硬组织中广泛应用[20-22],并取得了比传统测量学法更好的效果,但在头足类角质颚的分类应用还未曾见到,并且其分析效果也就未知。其中角质颚是头足类的重要摄食器官,各个部分都有明显独特的特征,不同种类间也有特征差异,可以通过其形态来探究头足类不同群体特征和食性转变的关系[22-23]。因此本文利用传统测量学法和傅里叶分析法对北太平洋柔鱼(Ommastrephes bartramii)和中东太平洋茎柔鱼(Dosidicus gigas)2种大洋性柔鱼类进行分类识别,比较2种方法的鉴别效果,为头足类种类识别提供新的途径和方法。
1 材料和方法
1.1 材料
柔鱼样本来源于北太平洋海域,探捕时间为2013年8~10月,作业海域在152°25′E~154° 37′E、41°29′N~43°27′N。茎柔鱼样本采集于中东太平洋海域,探捕时间为2014年4~7月,作业海域在114°59′W~119°30′W、0°28′N~1°12′N。样本采集时从2种渔货中分别随机抽取,经渔船冷冻保藏运回实验室。样本基本情况详见表1。
1.2 方法
1.2.1 角质颚外形测量
实验室解冻后按照胴部背面中线最前端至最后端的长度测定胴长(ML),测量时结果精确度精确至1 mm。从柔鱼和茎柔鱼样本的口球中提取角质颚,将外部清洗后用游标卡尺进行测量。
沿垂直和水平方向校准后依次测量角质颚的上头盖长(upper hood length,UHL)、上脊突长(upper crest length,UCL)、上喙长(upper rostrum length,URL)、上喙宽(upper rostrum width,URW)、上侧壁长(upper lateral wall length,ULWL)、上翼长(upper wing length,UWL)、下头盖长(lower hood length,LHL)、下脊突长(lower crest length,LCL)、下喙长(lower rostrum length,LRL)、下喙宽(lower rostrum width,LRW)、下侧壁长(lower lateral wall length,LLWL)、下翼长(lower wing length,LWL)共12项形态参数进行测量(图1),测量结果精确至0.1 mm。根据以上测量的外部形态数据,利用t检验对2种柔鱼类的角质颚进行差异性分析。
1.2.2 傅里叶分析
将2种柔鱼类角质颚置于Olympus光学显微镜40倍下拍照,获取平面图像。接下来采用“SHAPE”软件中的“Chain coder”程序和“Chc2Nef”程序将获得的角质颚图像转化为傅里叶系数。其中“Chain coder”程序可以将角质颚的外部形态描述成0~7的数字编码链信息,同时利用“Gray Scale”将外部形态转化为可识别的二进制灰度图像,保存为编码文件。然后将编码文件利用“Chc2Nef”程序依次将编码文件创建为傅里叶系数[24],一般认为20组傅里叶系数可以描述外部形态[25],每一组傅里叶系数由4个形态系数A、B、C、D组成,一共有80个系数组成。为了避免角质颚因大小、方向和摆放位置因素等的影响,所得傅里叶系数应做标准化处理,其中A1=1,B1=C1=0为常数,应当去除。最终每个角质颚外部形态由77个傅里叶系数组成。
1.2.3 传统判别函数建立
利用Wilks’Lambda法对12项参数值指标进行逐步筛选,同时采用逐步判别分析法(stepwise discriminant analysis,SDA)对筛选获得的参数进行判别分析,建立判别函数求得判别正确率[26]。

表1 2种柔鱼类样本基本信息Tab.1 Sam p le information of O.bartram ii and D.gigas
2 结果与分析
2.1 外部形态的差异分析
对于2种柔鱼类外部形态参数值统计详见表2。从表2中可以看出,在2种柔鱼类12项上、下角质颚参数值中的上头盖长(UHL)、上脊突长(UCL)、上喙长(URL)、上喙宽(URW)、上侧壁长(ULWL)、下脊突长(LCL)、下喙长(LRL)、下喙宽(LRW)、下侧壁长(LLWL)、下翼长(LWL)存在显著性差异(P<0.01),上翼长(UWL)、下头盖长(LHL)存在显著性差异(P>0.05)。下头盖长(LHL)形态参数值的平均值大于茎柔鱼,其它参数大小则均有所不同,通过观察发现2种柔鱼类角质颚形态有所差异:2种上头盖后缘的角度柔鱼大于茎柔鱼,下颚喙部柔鱼呈鹰喙状,而茎柔鱼则不明显,其它形态的辨识度不高(图2)。

图1 角质颚外部形态测量示意图(a:上角质颚长,b:下角质颚长)Fig.1 Scheme ofmorphometricmeasurement for beaks(a:upper beak length,b:lower beak length)

表2 柔鱼、茎柔鱼上、下角质颚测量参数比较Tab.2 M orphological com parison of upper beak and lower beak of O.bartram ii and D.gigas

图2 2种柔鱼类上、下角质颚外部形态示意图Fig.2 Scheme ofm orphometric m easurements of upper beak and lower beak of O.bartram ii and D.gigas
2.2 主成分分析
基于选取的相关系数,对不同种类间的柔鱼类角质颚进行主成分分析。处理后的形态特征参数,经计算获得相关系数矩阵,然后求出特征方程|R-λI|=0的p个非负的特征值λ1>λ2>…>λp≥0进行因子筛选,选取前面m(m<p)个主分量Z1,Z2,…,Zm为第1、2、…、m个主分量,当这m个主分量的方差和占全部总方差的60%以上,基本上保留了原来绝大部分因子的信息,即选取Z1,Z2,…,Zm作为主要因子分析差异[27]。
2.2.1 传统测量分析
为避免样品规格差异对形态参数值的影响,将12个角质颚形态参数分别除以胴长(ML),转化为上头盖长/胴长(UHL/ML)、上脊突长/胴长(UCL/ML)、上喙长/胴长(URL/ML)、上喙宽/胴长(URWML)、上侧壁长/胴长(ULWL/ML)、上翼长/胴长(UWL/ML)、下头盖长/胴长(LHL/ML)下脊突长/胴长(LCL/ML)、下喙长/胴长(LRL/ML)、下喙宽/胴长(LRW/ML)、下侧壁长/胴长(LLWL/ML)、下翼长/胴长(LWL/ML)的形态比例指标进行后续分析。主成分分析结果认为,柔鱼和茎柔鱼12项参数值前两个主成分贡献率已达到52.30%,第一主成分特征值为4.827。第一主成分主要反映在下喙长/胴长(LRL/ML),解释总变异的40.22%;第二主成分特征值为1.449,主要反映在下头盖长/胴长(LHL/ML)这一形态参数中,解释总变异的12.08%。
依据12项形态参数值得第一、二主成分的作出两个种类的散点图(图3)。散点图的分布可以看出主要在第二因子得分不同,而在第一因子处显示较大的重叠,第二因子区分不明显。

表3 传统测量法对柔鱼、茎柔鱼耳石形态的主成分分析Tab.3 Principal analysis of beak morphology for O.bartram ii and D.gigas based on traditionalmeasurement

图3 2种柔鱼类第一、第二主成分散点图Fig.3 Scatter plot of scores on the 1stand 2ndprinciple com ponents of O.bartramii and D.gigas
2.2.2傅里叶分析
主成分分析结果显示,2种柔鱼类上、下颚的前16个主成分累计贡献率分别达83.58%和73.76%。其中前5个主成分解释总变异的49.79%和42.02%,前10个主成分解释总变异的70.93%和60.01%。前16个主成分得分较高的傅里叶系数分别对应A2、C6、A6、B9、D8、D9、D6、A3、B5、A9、B3、D11、A3、C7、A7、B10。
两个种类的上、下颚傅里叶分散点图如图4所示。依据散点图分布可以看出2种类上颚主要在第一因子得分不同,而下颚主要在第二因子得分处不同。2种类傅里叶系数主成分分析认为二者重叠较为明显,区分效果不是很好。但上颚主成分分析第一因子得分差异较大,下颚则在第二因子得分处差异较大。
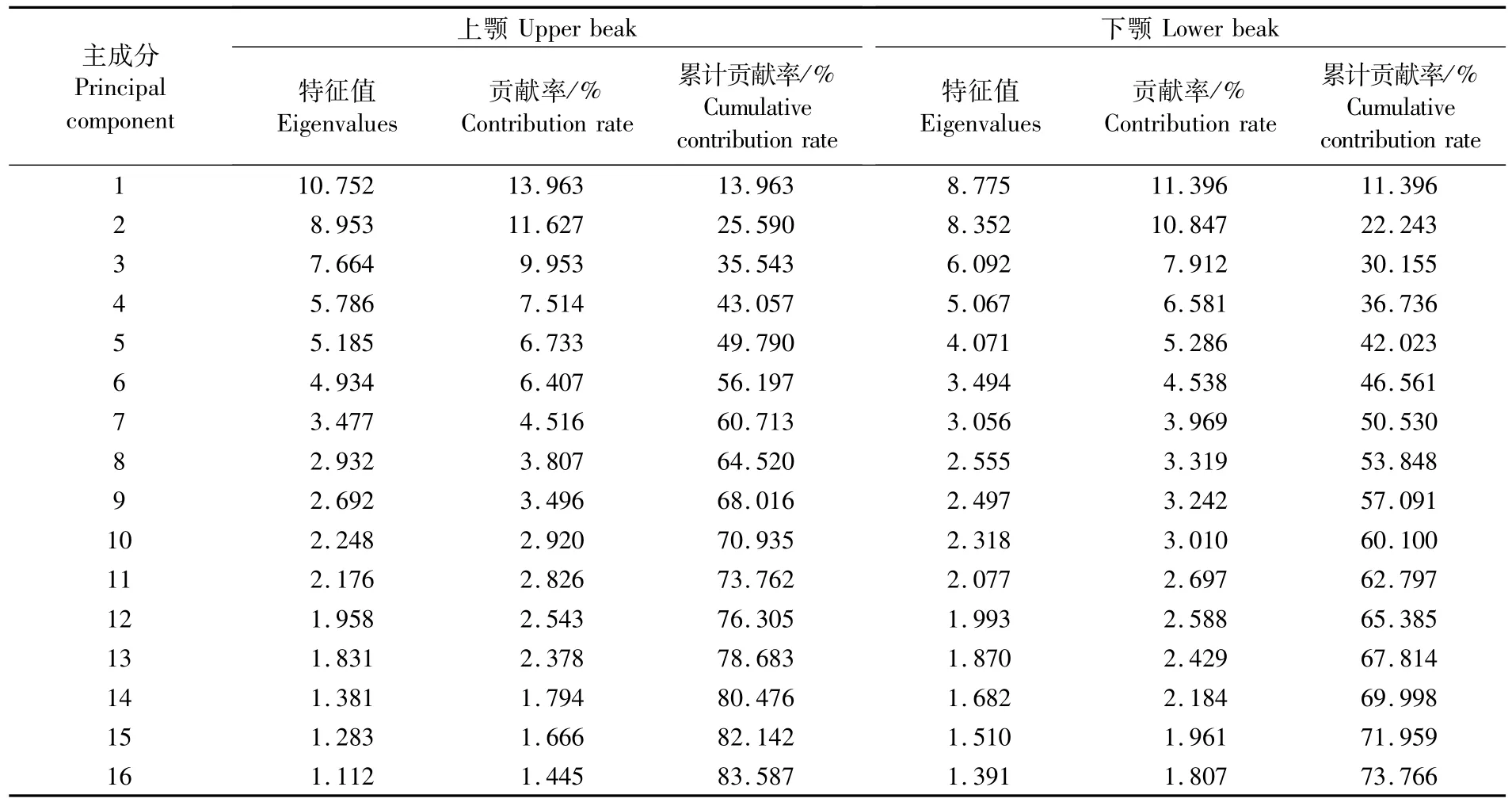
表4 傅里叶分析法对2种柔鱼类上、下角质颚形态的主成分分析Tab.4 Principal analysis of upper beak and lower beak morphology for O.bartram ii and D.gigas based on Fourier analysis

图4 傅里叶分析法2种柔鱼类上、下颚主成分因子分布图Fig.4 Scatter p lot of scores on the 1stand 2ndprincip le com ponents of upper beak and lower beak of O.bartrami and D.gigas
2.3 判别分析
2.3.1 传统测量判别分析
利用SPSS软件对两种柔鱼类的耳石进行判别分析,选择Wilks’Lambda的逐步判别法筛选出上头盖长/胴长(UHL/ML)、上喙长/胴长(URL/ML)、上侧壁长/胴长(ULWL/ML)、下头盖长/胴长(LHL/ML)、下喙长/胴长(LRL/ML)、下喙宽/胴长(LRW/ML)6项参数因子对2种柔鱼类进行种类鉴别,建立判别方程:
柔鱼:

茎柔鱼:

将测量的角质颚形态测量值依次代入上述判别函数,Y值即为归入该种类的判别正确率。判别结果显示北太平洋柔鱼的判别率为96.9%,中东太平洋茎柔鱼判别率为93.1%,总判别正确率为95.00%。交互验证结果与初始判别基本相同,北太平洋柔鱼的判别率为93.8%,中东太平洋茎柔鱼判别率为93.1%,总判别正确率为93.45%(表5)。
2.3.2 傅里叶判别分析
将角质颚外部形态提取的77个傅里叶系数进行逐步判别分析(SDA),采用Wilks’Lambda法筛选系数,最终选择A2、A3、A10、A13、A17、B6、B7、C2、C3、C6、C7、C9、C1、D3、D4、D5、D6、D7共18个傅里叶系数纳入逐步判别分析对2种柔鱼类上、下颚进行判别分类,判别方程如下:
上角质颚柔鱼:

上角质颚茎柔鱼:

下角质颚柔鱼:

下角质颚茎柔鱼:
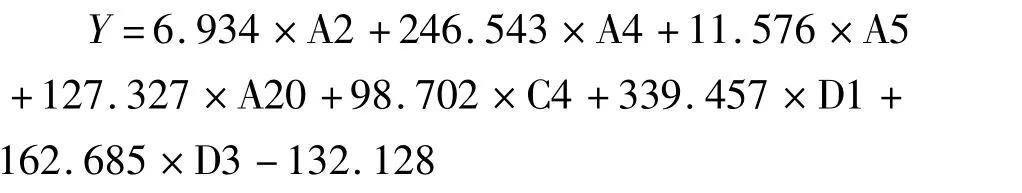
将对应的傅里叶系数代入上述判别方程函数,从表6和表7结果来看,上角质颚柔鱼和茎柔鱼的判别正确率分别为96.9%和98.3%,总判别率为97.6%;下角质颚柔鱼和茎柔鱼的判别率分别为80.0%和91.4%,总判别率为85.7%。交互验证法相比于初始判别有所降低,上角质颚和下角质颚总体判别率分别为94.4%和83.1%。

表5 传统测量法的2种柔鱼类判别结果Tab.5 Result of classification between O.bartram ii and D.gigas based on traditionalm easurement by SDA
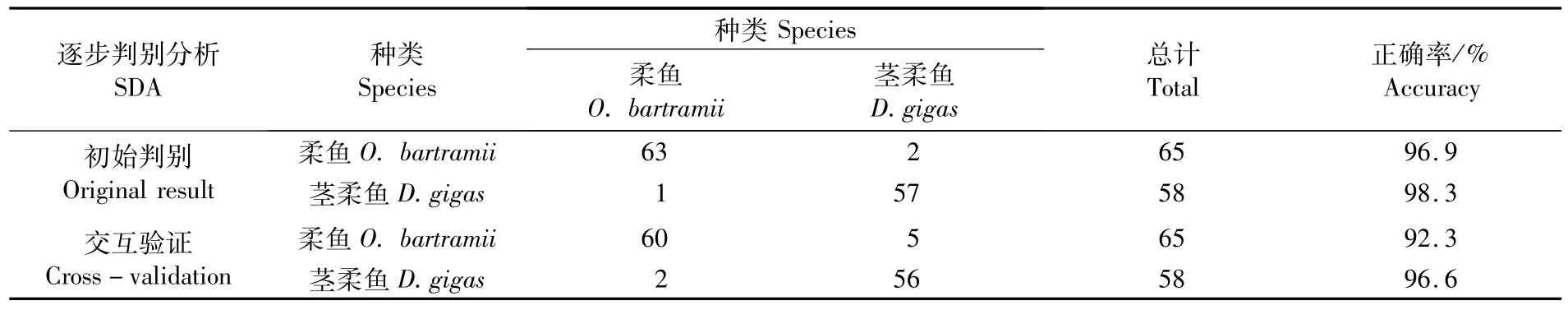
表6 傅里叶分析法的2种柔鱼类上颚判别结果Tab.6 Result of classification for upper beak between O.bartram ii and D.gigas based on Fourier analysis by SDA

表7 傅里叶分析法的2种柔鱼类下颚判别结果Tab.7 Result of classification for lower beak of O.bartram ii and D.gigas based on Fourier analysis by SDA
3 讨论
头足类的形态特征主要体现在外部胴体形态和硬组织中,相比于胴体特征,硬组织形态特征稳定,耐腐蚀,不会受到海洋环境和人为捕捞的影响,已经成为头足类种间、种群差异和物种鉴定的良好材料[28-29]。目前鉴定种类的方法有形态法、生化法和遗传法,其中形态法以其简单易行、易掌握等广泛应用于物种鉴定中。本文利用传统测量法对上颚7个形态值和下颚5个形态值进行参数分析,能够较为准确地体现出角质颚的外部形态特征。主成分分析结果显示2种柔鱼类差异主要体现在下喙长/胴长(LRL/ML)和下头盖长/胴长(LHL/ML)这两个形态参数中。VEGA[30]利用下颚的7个形态特征值对东南太平洋智利沿岸28种头足类进行种类划分,其结果与本文的下颚贡献率高相吻合。本研究利用传统测量法对两种类的角质颚建立判别函数,判别正确率达到95%,CHEN等[31]也通过此方法对柔鱼、茎柔鱼、鸢乌贼(Sthenoteuthis oualaniensis)和阿根廷滑柔鱼(Illex agentinus)4个种类的角质颚进行判别分析,发现种间判别率也均超过95%,这些说明传统侧量法在头足类判别中有良好的判别效果。
传统测量法最终将上头盖长/胴长、上喙长/胴长、上侧壁长/胴长、下头盖长/胴长、下喙长/胴长、下喙宽/胴长6项形态参数值纳入到判别分析中,总判别率达95%。KEAR[32]认为角质颚的摄食运动主要有上颚肌、侧肌和下颚肌组成。其中上头盖和上喙位于上颚肌,在运动中起闭合作用;上侧壁位于侧肌,负责侧壁展开以配合舌齿和口须的运动;下头盖、下喙位于下颚肌,控制舌齿和口须摄食,在运动中起主要作用。由于不同种类的头足类摄食习性不同导致运动模式也存在差异[33],因此这几个器官的参数值在本研究分类鉴定中具有较好的效果。
对于外部形态不规则,或不同年龄和生长阶段的硬组织而言,传统测量法无法检测其微小变化,因此会有一定的判别误差[34-35]。因此本文利用傅里叶分析法进行分析,比较2种方法鉴别效果。将上、下角质颚转化为20组傅里叶系数,对外部形态特征的描述更为精细,此方法在鱼类耳石中有广泛的应用[36-37]。傅里叶分析法对上、下角质颚进行判别分析,求得上角质颚判别率为97.6%,下角质颚总判别率为85.7%,傅里叶分析法对上颚有更好的判别效果。而CLARKE[38]认为下颚具有易采集、形态特征稳定且差异较为明显的特点,更适合作为分类材料,这与本文傅里叶分析法上颚判别率高于下颚有所不同,主要是由于傅里叶分析法是一种数学分析方法,而上颚的外部形态相比于下颚更趋近于拟合的椭圆,因此更适合椭圆傅里叶分析。
本文对北太平洋海域柔鱼和中东太平洋海域茎柔鱼上、下角质颚外部形态进行比较分析,利用传统测量法和傅里叶分析法分别进行识别,2种形态分析方法均有良好的识别效果,研究结果表明傅里叶分析法不仅适用于头足类耳石,在角质颚分类鉴别中同样有效,这为头足类的种类鉴定提供一个有效的识别方法。在今后的头足类种类识别研究中,应加以分析角质颚的长度比值[39]、生长特性[40],同时结合其微量元素信息[41],并与其它硬组织的形态特征进行对比分析,来提高头足类种类判别的准确性。
[1] 陈新军,刘必林,王尧耕.世界头足类[M].北京:海洋出版社,2009:189-215.
CHEN X J,LIU B L,WANG Y G.The world cephalopods[M].Beijing:China Ocean Press,2009:189-215.
[2] 王尧耕,陈新军.世界大洋性经济柔鱼类资源及其渔业[M].北京:海洋出版社,2005:58-264.
WANG Y G,CHEN X J.The world ocean economic cephalopods resources and fisheries[M].Beijing:China Ocean Press,2005:58-264.
[3] 周金官,陈新军,刘必林.世界头足类资源开发利用现状及其潜力[J].海洋渔业,2008,30(3):268-275.
ZHOU J G,CHEN X J,LIU B L.The world cephalopods resources development and utilization status and potential[J].Marine Fisheries,2008,30(3):268-275.
[4] 由上龍嗣,檜山義明,依田真里,等.平成17年マサバ对馬暖流系群の資源評価[R].西海区水産研究所,2006.
YOSHIAKIH,TADASHIA,YODA M,et al.Stock assessment of Tsushima Current Chubmackerel stock in 2005[R].West China Sea Fisheries Research Institute of Japan,2006.
[5] SMALEM J,CLARKE M R,KLAGESN TW,et al.Octopod beak identification-resolution at a regional level(Cephalopoda,Octopoda:Southern Africa)[J].South African Journal of Marine Science,1993,13(1):269-293.
[6] GROEGER J,PIATKOWSKIU,HEINEMANNH.Beak length analysis of the Southern Ocean squidPsychroteuthisglacialis(Cephalopoda:Psychroteuthidae)and its use for size and biomass estimation[J].Polar Biology,2000,23(1):70-74.
[7] LU C C,ICKERINGILL R.Cephalopod beak identification and biomass estimation techniques:tools for dietary studies of southern Australian finfishes[M].Museum Victoria,2002:65-69.
[8] 方 舟,陈新军,陆化杰,等.头足类角质类研究进展Ⅱ—运动、分类与生态[J].海洋渔业,2014,36(2):183-192.
FANG Z,CHEN X J,LU H J,et al.Progress of beak in cephalopodⅡ—movement,taxnomy and ecology[J].Marine Fisheries,2014,36(2):183-192.
[9] 苏 杭,方 舟,陈新军,等.基于两种形态学方法的柔鱼类耳石判别比较[J].海洋渔业,2015,37(4):293-301.
SU H,FANG Z,CHEN X J,et al.Comparative study of Ommastrephidae species indentification based on statolith shape[J].Marine Fisheries,2015,37(4):293-301.
[10] JACKSON G D,BUXTON N G,GEORGE M J.Beak length analysis ofMoroteuthis ingens(Cephalopoda:Onychoteuthidae)from the Falkland Islands region of the Patagonian Shelf[J].1997(77):1235-1238.
[11] MORI J.Geographical differences between the parasites’infection levels of the neon flying squid(Ommastrephes bartramii)from the North Pacific Ocean(Abstract)[R]//Reportof the1995 Meeting on Squid Resources.Contributions to the Fisheries Researches in the Japan Sea Block,No.36.Japan Sea National Fisheries Research Institute,Niigata,1998:85-86.
[12] NAGASAWA K,MORI J,OKAMURA H.Parasites as biological tags of stocks of neon flying squid(Ommastrephes bartramii)in the North Pacific Ocean[R]//O T,ed.Contributed Papers to International Symposium on Large Pelagic Squids.Tokyo:Japan Marine Fishery Resources Research Center,1998:49-64.
[13] YASTUA,TANAKE H,MOORI J.Population structure of the neon flying squid,Ommastrephes bartramii,in the North Pacific Ocean[R]//Ok T d.Contributed Papers to International Symposium on Large Pelagic Squids.Tokyo:Japan Marine Fishery Resources Research Center,1998:31-48.
[14] BOOKSTEIN F L.Size and shape spaces for landmark data in two dimensions[J].Statistical Science,1986:181-222.
[15] ROHLF F J.Relationships among eigenshape analysis,Fourier analysis,and analysis of coordinates[J].Mathematical Geology,1986,18(8):845-854.
[16] ROHLF F J,SLICE D.Extensions of the Procrustes method for the optimal superimposition of landmarks[J].Systematic Biology,1990,39(1):40-59.
[17] KENDALL D G.Shape-manifolds,procrustean metrics and complex projective spaces[J].Bulletin of the London Mathematical Society,1984,16(2):81-121.
[18] BOOKSTEIN F L.Biometrics,biomathematics and the morphometric synthesis[J].Bulletin of Mathematical Biology,1996,58(2):313-365.
[19] ROHLF F J,MARCUS L F.A revolution in morphometrics[J].Trends in Ecology and Evolution,1993,8(4):129-132.
[20] 郭弘艺,魏 凯,唐文乔,等.基于矢耳石形态特征的中国鲚属鱼类种类识别[J].动物分类学报,2010,35(1):127-134.
GUO H Y,WEI K,TANG W Q,et al.Sibling species discrimination for Chinese genus of coilia fishes based on sagittal otolith morphology[J].Acta Zoo Taxonomica Sinica,2010,35(1):127-134.
[21] CASTRO JJ,HERNANDEZ-GARCIA V.Ontogenetic changes in mouth structures,foraging behaviour and habitat use ofScomber japonicusandIllex coindetii[J].Scientia Marina,1995.
[22] 叶振江,孟晓梦,高天翔,等.中日2种花鲈耳石形态的地理变异[J].海洋与湖沼,2007,38(4):356-360.
YE Z J,MENG X M,GAO T X,et al.The geoeraphical discrimination in otolith morphology of sea bass:lateolabrax japonicasandL.maculates[J].Oceanologia Et Limnologia Sinica,2007,38(4):356-360.
[23] 王英俊,叶振江,刘 群,等.细条天竺鱼(Apogonichthys lineatus)与黑鳃天竺鱼(Apogonichthys arafurae)耳石形态识别的初步研究[J].海洋与湖沼,2010,41(2):281-284.
WANG Y J,YE Z J,LIU Q,et al.Using otolith to identify two fish species:Apogonichthy lineatusandA.aarafurae[J].Oceanologia Et Limnologia Sinica,2010,41(2):281-284.
[24] KUHL F P,GIARDINA CR.Elliptic Fourier features of a closed contour[J].Computer Graphics and Image Processing,1982,18(3):236-258.
[25] FERGUSONG,MESSENGER J,B.Gravity and light influence the countershading reflexes of the cuttlefish Sepia officinalis[J].Journal of Experimental Biology,1994,191(1):247-256.
[26] 李思发,李晨虹.尼罗罗非鱼品系间形态差异分析[J].动物学报,1998,44(4):450-457.
LISF,LIC H.Analysis ofmorphological variations among strains of line tilapia(Oreochroems niloticus)[J].Acta Zoo Sinica,1998,44(4):450-457.
[27] TANG Q Y,FENG M G.DPS data processing system:experimental design,statistical analysis and data mining[J].Science,2007,12(4):43-45.
[28] IVERSON I L K,PINKAS L.A pictoral guide to beaks of certain eastern Pacific cephalopods[J].Fish Bulletin,1971(152):83-105.
[29] ALLCOCK A L,PIERTNEY S B.Evolutionary relationships of Southern Ocean Octopodidae(Cephalopoda:Octopoda)and a new diagnosis of Pareledone[J].Marine Biotechnology,2002(140):129-135.
[30] VEGA M A.Uso de lamorfometría de lasmandíbulas de cefalópodos en estudios de contenido estomacal[J].Latin American Journal of Aquatic Research,2011,39(3):600-606.
[31] CHEN X,LUH,LIU B,et al.Species identification of Ommastrephes bartramii,Dosidicus gigas,Sthenoteuthis oualaniensis and Illex argentinus(Ommastrephidae)using beak morphological variables[J].Scientia Marina,2012,76(3):473-481.
[32] KEAR A J.Morphology and function of themandibular muscles in some coleoid cephalopods[J].Journal of the Marine Biological Association of the United Kingdom,1994,74(4):801-822.
[33] NIXON M.The feeding mechanisms and diets of cephalopods-living and fossil[M]//WIEDMANN J,KULLMANN J.Cephalopods present and past,Germany:Springer,1988:641-652.
[34] BIRD J L,EPPLER D T,CHECKLEY JR D M.Comparisons of herring otoliths using Fourier series shape analysis[J].Canadian Journal of Fisheries and Aquatic Sciences,1986,43(6):1228-1234.
[35] CASTONGUAYM,SIMARD P,GAGNON P.Usefulness of Fourier analysis of otolith shape for Atlantic mackerel(Scomber scombrus)stock discrimination[J].Canadian Journal of Fisheries and Aquatic Sciences,1991,48(2):296-302.
[36] CAMPANA SE,CASSELMAN JM.Stock discrimination using otolith shape analysis[J].Canadian Journal of Fisheries and Aquatic Sciences,1993,50(5):1062-1083.
[37] 李辉华,郭弘艺,唐文乔,等.2种耳石分析法在鲚属种间和种群间识别效果的比较研究[J].淡水渔业,2013,43(1):14-18.
LIH H,GUO H Y,TANGW Q,et al.Comparative study of two otolith shape analysis for genusCoillaspecies and stocks identification[J].Freshwater Fisheries,2013,43(1):14-18.
[38] CLARKE M R.A handbook for the identification of cephalopod beaks[M].Oxford:Clarendon Press,1986:273.
[39] 杨林林,姜亚洲,刘尊雷,等.东海太平洋褶柔鱼角质颚的形态学分析[J].中国海洋大学学报,2012,42(10):13-15.
YANG L L,JIANG Y Z.LIU Z L,et al.Variation analysis on partial morphometric measurements of beak oftodarodes pacificusinhabiting East China Sea[J].Periodical of Ocean University of China,2012,42(10):13-15.
[40] 方 舟,陈新军,陆化杰,等.北太平洋两个柔鱼群体角质颚形态及生长特征[J].生态学报,2014,34(19):5405-5415.
FANG Z,CHEN X J,LU H J,et al.Morphology and growth of beaks in two cohorts for neon flying squid(Ommastrephes bartramii)in the North Pacific Ocean[J].Acta Ecological Sinica,2014,34(19):5405-5415.
[41] 李云凯,贡 艺,陈新军.稳定同位素技术在头足类摄食生态学研究中的应用[J].应用生态学报,2014,25(5):1541-1546.
LI Y K,GONG Y,CHEN X J.Applications of stable isotope analysis in the trophic ecology studies of cephalopods[J].Chinese Journal of Applied Ecology,2014,25(5):1541-1546.
Beak shape identification of two oceanic squids based on differentmethods
SU Hang1,4,FANG Zhou1,4,CHEN Xin-jun1,2,3,4
(1.College of Marine Sciences,Shanghai Ocean University,Shanghai201306,China;2.National Engineering Research Center for Oceanic Fisheries,Shanghai Ocean University,Shanghai201306,China;3.Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources,Ministry of Education,Shanghai Ocean University,Shanghai 201306,China;4.Collaborative Innovation Center for Distant-water Fisheries,Shanghai201306)
Identification of upper beaks and lower beaks was investigated in geometric among two species of Ommastrephide:Ommastrephes bartramiicaught in the North Pacific from August to October in 2013,Dosidicus gigas caught in equatorialwaters of the Equator from April to July in 2014.Three upper beaks and lower beaks were analyzed based on traditionalmeasurement and Fourier analysis for species identification.Twelve variables were measured on the upper beak and lower beak;digitalized photographs of beaks were transformed to Ellipse Fourier descriptors(EFDs).Finally,77 standardized EFDswere analyzed by principal component analysis(PCA)and stepwise discriminant analysis(SDA).Results showed that the upper hood length(UHL),upper crest length(UCL),upper rostrum length(URL),upper rostrum width(URW),upper lateralwall length(ULWL),lower crest length(LCL),lower rostrum length(LRL),lower rostrum width(LRW),lower crostwall length(LLWL),lower wing length(LWL)had significant differences between two species(P<0.01),upperwall length(UWL),lower hood length(LHL)had no significant difference between two species(P>0.05).Results of PCA showed that the highest variables were lower rostrum length(LRL)and lower rostrum width(LRW),first sixteen components accounted for 83.58%of total contribution of mutation,and the scatter-plot revealed relative overlapping within two species.There were 16 principal components with 83.58%cumulative contribution of upper beak and 73.76%cumulative contribution of lower beak.SDA showed that dorsal lateral length(DLL/ML),lateral dome length(LDL/ML),wing length(WL/ML),rostrum width(RW/ML)entered as coefficients in the function,total correct classification was 95%;18 harmonics of 77 EFDs entered in SDA in Fourier analysiswith total correct classification of97.6% and 85.7%.Overall,traditionalmeasurement and Fourier analysis can identify Ommastrephes bartramii andDosidicus gigaseffectively with their beak morphology,these findings can offer an effective measurement to identify species for Ommastrephes bartramii.
beak;traditionalmorphometrics;elliptical fourier analysis;Ommastrephes bartramii;Dosidicusgigas;stepwise discriminant analysis
S 931.5
A
1004-2490(2016)03-0225-11
2015-07-26
国家自然科学基金(NSFC41476129;NSFC41276156);国家863计划(2012AA092303);上海市研究生教育创新计划和国家科技支撑计划(2013BAD13B01)
苏 杭(1992-),男,河南商丘人,硕士研究生,主要从事渔业资源学研究。E-mail:615923118@qq.com
陈新军,教授。Tel:021-61900306,E-mail:xjchen@shou.edu.cn
