内隐序列学习意识的具身机制*
2016-02-01张剑心查德华黄建平刘电芝
张剑心 汤 旦 查德华 黄建平 刘电芝
(苏州大学教育学院,苏州 215123)
2005年 Science杂志遴选出 125个重大科学问题,第 2个就是“生物的意识是怎么产生的”?无意识–意识时刻伴随着人类,无意识知识上升为意识知识后,意识就能够对知识进行灵活控制、整合和迁移(Dehaene,Charles,King,&Marti,2014;Dehaene,Changeux,&Naccache,2011),可见意识状态怎样,很大程度决定着人类的学习效果与工作质量。在内隐序列学习进程中,意识是怎么产生的?对该问题的探讨,有助于揭示意识产生的本源。
已有意识理论,如神经可塑性理论和新异刺激理论只考虑刺激驱动,未考虑人的主观驱动因素;全局工作平台(globle workspace)理论虽考虑了主观驱动,但忽略了身体感受的关键作用。具身认知理论和具身意识理论及研究表明,人的认知活动(包括意识)无不与具身感受紧密相关,但迄今为止内隐序列学习意识与身体存在何种关系尚无揭示。本文将以具身意识理论为基础,探索内隐序列学习规则意识产生的具身原因和过程。先介绍内隐序列学习意识的已有理论和缺陷,再详细介绍能弥补这些缺陷且揭示意识根本机制的具身意识理论,然后就把具身意识理论引入内隐序列学习意识领域,最后对未来研究进行展望。
1 内隐序列学习意识的三类理论
近年来研究者们普遍认可且与内隐序列学习意识紧密相关的意识理论可分为三类:
1.1 全局工作平台理论
全局工作平台理论(globle workspace theory,Dehaene,Changeux,Naccache,Sackur,&Sergent,2006)认为意识要产生,必须在全脑建立长时程的工作平台,即感知觉运动脑区与额叶—顶叶—扣带回的认知控制系统产生长时程连接(Dehaene et al.,2011;Wyart,Dehaene,&Tallon-Baudry,2012)。意识是各脑区的整合,形成了区别于其它表征的不可分的独立表征,同时抑制其它表征(Tononi,2008)。该理论在掩蔽刺激意识范式中得到了很多证据。Del Cul,Baillet和Dehaene (2007)发现看到和没看到刺激的试次,在感知觉脑区的早期加工没有差异;但是在中期,意识到的刺激激活了额叶—顶叶—扣带回认知控制系统,出现了 P3b,并且beta波段同步增强(Gaillard et al.,2009);在后期认知控制系统产生自上而下的调控,从而在感知觉运动区出现了更大的 gamma波(Ray &Maunsell,2010),这表明只有意识到的刺激才建立了全局工作平台。Imamoglu,Kahnt,Koch和Haynes (2012)采用大脑网络技术,直接证明了只有意识到的视觉刺激在前额叶和初级皮层建立了长时程的网络联结。King等(2013)则在听觉意识中验证了全局工作平台理论。Dehaene等(2014)总结了全局工作平台的脑区,包括背外侧前额叶、下顶叶、中颞叶和楔前叶,被研究者们称为“富人俱乐部” (rich club)网络(van den Heuvel &Sporns,2011;van den Heuvel,Kahn,Goñi,&Sporns,2012)。该理论还在睡眠研究和意识障碍研究中得到证实和发展,揭示了更广泛的意识支持网络,包括脑干、丘脑、脑岛等(Schröter et al.,2012)。
该理论认为,意识的产生是有或无,其关键在于全局工作平台能否激活,这取决于刺激呈现前(即试次前)随机波动的大脑觉醒状态,包括注意、工作记忆、目的动机、情感等(Aru,Bachmann,Singer,&Melloni,2012;Sadaghiani,Hesselmann,&Kleinschmidt,2009;Boly et al.,2007),因此试次前基线状态是决定意识的重要因素。但是造成随机波动的因素特别是身体因素尚不清楚,具身意识理论或能提供原因;且试次前随机波动的解释忽视了试次中的刺激驱动,而表征质量理论可对此完美解释(见下一节)。
掩蔽刺激意识范式和内隐序列学习意识范式是无意识转化为意识的两大经典范式,具有共通之处;全局工作平台理论很可能适用于几乎所有意识(Dehaene et al.,2014;Dehaene &Changeux,2011),因此 Rünger和 Frensch (2010)采用全局工作平台理论来定义和解释内隐序列学习意识。Rose,Haider和 Büchel (2010)及 Wessel,Haider和Rose (2012)采用内隐序列学习的变式,证明右侧前额叶和枕叶的gamma波的耦合增强能预测内隐转化为外显知识,相应的右侧腹外侧前额叶皮质和腹侧纹状体的功能联结增强,提供了内隐序列学习意识符合全局工作平台理论的证据。但笔者认为该理论主要是对意识的描述,没有给出意识的来源;且既未考虑意识产生的刺激驱动机制,又没有揭示对刺激的动机、情感、经验、元认知等主观因素对意识的影响机制——随机波动还只是假设性和描述性的,因为这些主观因素的波动可能仅仅是表现为随机而已,可能它们实质上是被某些认知和人格特质所决定,而充当了中介变量的角色。而且该理论产生于掩蔽刺激意识范式,研究的是比较低级的知觉意识,只有看见/没看见两种意识状态,对应的全局工作平台的激活就只有全或无两种状态;内隐序列学习意识是由隐藏的序列规则产生的高级意识,包含了上述诸多主观因素,参与脑区更多且能够产生各种具有渐进性的中间意识状态(详见下一节),因此全局工作平台理论还不能直接应用于内隐序列学习意识,需要采用新的假设——全局工作平台渐进激活并加以验证,并可引入支持渐进意识的表征质量理论来弥补。
1.2 表征质量理论和神经可塑性理论
表征质量理论则考虑了刺激驱动和渐进意识。Destrebecqz和 Cleeremans (2001,2003)发现随反应刺激间隔RSI (response stimulus interval)增加,内隐序列学习从完全内隐向外显渐进变化,由此Cleeremans和 Jiménez (2002)提出了意识表征质量理论:表征质量程度决定了意识程度,内隐和外显只是表征质量从低到高渐进变化的连续体上的两点,两者之间只有表征质量的量的差异,没有质的差异,因此他们推测应该存在具有意识渐进性的多种中间意识状态。之后果然 Dienes和Scott (2005)、Norman,Price和Duff (2006)、Norman,Price,Duff和 Mentzoni (2007)、Fu,Dienes和 Fu(2010)、Norman (2010)等在人工语法或内隐序列学习范式中都证实存在介于完全内隐和外显之间的多种中间意识状态,支持了意识表征质量理论(详见综述:张剑心,黄颀,张润来,刘电芝,2015)。因此张润来、刘电芝、张剑心和程昊旸(2013)认为该理论是内隐序列学习意识的良好的解释性理论,揭示了意识产生的具体机制。但是张剑心等(2015)认为 Cleeremans和 Jiménez (2002)的意识表征质量理论只用单一的表征质量系统来说明所有意识状态,无法解释内隐外显的实验性分离(Destrebecqz &Peigneux,2005),更无法解释内隐序列学习的加工脑区(纹状体)和意识脑区(扣带回)的分离(Simon et al.,2011),因此他们对意识表征质量理论进行改进,提出了双系统渐进意识理论——表征质量和意识是两个相对独立的系统,两个系统及互动的渐进变化产生了多种意识状态。
Cleeremans (2007,2011)在意识表征质量理论基础上,进一步提出具有普适性的神经可塑性理论——即意识产生是由于较强、独特、持久的表征重塑了全脑网络,这就同样打破了 Cleeremans和Jiménez (2002)的单系统假设,而关注广泛的脑区;并从本质上揭示了全局工作平台理论的独立表征的形成机制。神经可塑性理论就是意识的学习产生理论,在内隐序列学习意识、脑损伤患者的意识恢复、新的感知觉刺激的感受获得、社会认知和情感的获得等领域得到广泛支持(Askenasy&Lehmann,2013)。如:盲视患者可通过触觉训练,在视觉区建立视觉意识(Kupers,Pietrini,Ricciardi,&Ptito,2011);对两种难分辨的新气味进行训练,会产生新的感受意识(Li,Howard,Parrish,&Gottfried,2008);对掩蔽刺激的训练提高了阈下刺激的感知觉意识(Atas,Vermeiren,&Cleeremans,2013;Schwiedrzik,Singer,&Melloni,2009)。不过这些意识训练都存在脑区特异性的局限。Allen和Williams (2011)认为新的社会情感的获得很可能来自镜像神经元系统(MNS)的重塑;Davidson和McEwen (2012)则证实了压力和内部心理预防等社会认知因素通过对大脑网络的重塑来影响社会情感认知。
神经可塑性理论解释了意识产生机制,为意识训练提供了理论基础,但其训练有特异性的限制,普适性的意识训练(如对感受敏感性的训练)尚未得到探究(Frith,2011)。这需要考虑意识感受的基线,改变意识感受基线,就能增加意识敏感性,将普适于所有任务意识的提高,而这是具身意识理论的核心思想。
1.3 新异刺激理论
表征质量理论和神经可塑性理论只解释了恒常刺激意识产生的自身原因,而新异刺激理论则揭示了新异刺激对恒常刺激意识的促进机制。新异刺激理论认为:内隐序列学习意识的产生,不是恒常刺激表征质量增强引起的神经重塑,而是新异刺激的意外出现打断了恒常刺激的表征加工,使人产生了阻碍感和对比,从而对恒常刺激产生意识。Rünger和Frensch (2008)在内隐序列学习中设置两个组:实验组先学习6个组段的简单序列规则 A (6个不同位置组成的序列如 1-5-2-6-4-3)作为恒常刺激,再学习 6个组段的简单序列规则B (如 4-6-2-3-5-1)作为新异刺激,最后用生成任务测量 A的意识程度;控制组则一直学习 A,然后用生成任务测量A的意识程度;结果发现实验组对A的意识程度显著高于控制组。由于实验组只在训练阶段前期学习了 6个组段的 A,而控制组在训练阶段学习了12个组段的A,按照表征质量理论,应该是控制组的 A意识高于实验组;但是 Rünger和 Frensch (2008)却得到了相反结果,证明实验组更大的意识程度肯定有很大一部分来自于新异刺激 B的意外事件效应。Schwager,Rünger,Gaschler和 Frensch (2012)改变了 Rünger和Frensch (2008)的实验设计:实验组同样先学习6个组段的 A (恒常刺激),再学习 6个组段的 B(新异刺激),但是最后用生成任务测量新异刺激B (而非A)的意识程度;控制组则一直学习12个组段的 B,然后用生成任务测量 B的意识程度;结果发现实验组和控制组对B的意识程度没有差异,证明恒常刺激对新异刺激的意识促进效果至少与表征质量等同,其机制仍然是新异刺激的意外事件效应。两个研究对比,表明新异刺激和恒常刺激能互相促进各自的意识程度,当然新异刺激对恒常刺激意识的促进会更大些。
Rünger (2012)甚至在表征质量理论的经典实验中得到了反对表征质量理论的结果。他认为Cleeremans和 Jiménez (2002)的表征质量理论所依据的实验(Destrebecqz &Cleeremans,2001,2003)存在一个关键问题:内隐序列学习掺杂了非主序列和转移组块,这两种新异刺激可能导致了意识产生,而非表征质量增长的作用。因此Rünger (2012)采用了 Destrebecqz和 Cleeremans(2001,2003)的实验程序,但删除了非主序列和转移组块,结果没有发现训练 RSI对再认任务测得的意识的主效应,这表明在没有任何新异刺激时,训练阶段更大的RSI就无法增加意识程度了。由于 Cleeremans和 Jiménez (2002)是将 RSI作为表征质量的自变量,因此Rünger (2012)认为自己的实验证明了表征质量理论不成立。但是笔者认为这样的论断过于武断,因为还存在另外两种解释:RSI不是纯粹的表征质量的自变量;或再认任务对意识程度不敏感。比如Fu等(2010)操纵训练组段的数量,发现只训练6或7个组段只能产生内隐知识,训练15个组段则能产生受控意识,证明训练组段的数量是表征质量的自变量——该自变量从逻辑上和定义上都比RSI更纯粹。当然Fu等(2010)的实验仍然有非主序列,因此需加以删除以进一步验证,但至少Rünger (2012)的实验无法推翻其结果。
上述大部分实验的新异和恒常刺激的数量相等,且是分别集中学习,这就存在一个问题:新异刺激本身的表征质量就高,可能会产生迁移效应来促进恒常刺激的意识程度。张剑心,武燕,陈心韵和刘电芝(2014)证明在RSI为0 ms、250 ms、500 ms、750 ms、1000 ms时只需要学习8个组段,高情感开放性者就都能习得非主序列(概率仅为0.125),低情感开放性者则无法习得非主序列,这表明新异刺激与恒常刺激的作用是相互的,使得即使新异刺激的概率很低仍能被习得。那么当两者数量相等时,就更无法排除新异刺激的迁移效应。因此黄建平,张剑心和刘电芝(2014)在内隐序列学习中以转移组块作为新异刺激,操纵转移组块的数量(与学习组段相比,概率为1/6或1/12)和位置,由于新异刺激概率很低,因此能尽量避免新异刺激本身产生较高的表征质量和迁移效应,结果发现只有当存在两个转移组块且分布在训练阶段的前后期,才能显著提高恒常刺激的意识程度。该研究一方面采用更标准的低概率新异刺激的设置证明了新异刺激理论,另一方面更重要的是表明要促进恒常刺激的意识程度,新异刺激本身的表征质量必须足够,而其表征质量可能是非线性增长的:不只需要有足够数量,还需要处于恰当的位置,才能促使新异和恒常刺激的表征质量都非线性增长。可见虽然新异刺激理论宣称与表征质量理论有本质不同,甚至推翻了表征质量理论,但实质上能够用改进了的非线性表征质量理论来解释。
从以上所有研究可见新异刺激理论尚不成熟,且存在一个重大局限:新异刺激必然以恒常刺激为前提,因此解释力有限——比如不适用于只有恒常刺激没有新异刺激的情况。
1.4 三类理论的关系和缺陷
综上所述,全局工作平台理论对意识状态进行了精确描述,但缺乏刺激驱动和渐进意识机制;表征质量理论和神经可塑性理论则提供了刺激驱动的意识产生和变化机制,还对渐进意识的存在进行了很好地预测和解释,但是意识脑区可塑性具有特异性,难以迁移;新异刺激理论揭示了意识产生和突变机制,但需新异和恒常刺激对比,解释力有限。更关键的是,三类理论都忽略了身体对意识的关键作用,因此需要引入将身体看作是意识来源的具身意识理论。
2 具身意识理论
具身意识理论起源于现象学理论。Thompson和Varelam (2001)以及Noë(2004)从心理学意义上提出具身意识理论,认为身体是意识产生的最关键原因:意识来源于外部/内部的感知觉运动新刺激打破原身体状态基线,产生新的身体感受(Gallagher,2005;Thompson,2007);身体状态包括自主神经系统状态、运动/情感状态(Shapiro,2014,综述)。Bower和 Gallagher (2013)进一步提出外部/内部的新刺激除了感知觉运动刺激外,还应该包括突发的外部/内部动机、情感和自我认知等新刺激。
具身意识理论原理为:(1)大脑时刻接收着身体感受(各感知觉通道、情感、自我等),形成了身体感受基线;各种新刺激干扰了身体感受基线,从而对新刺激产生意识。此处新刺激不是新异刺激,因为新异刺激必须是和恒常刺激成对出现的,该理论将新出现的恒常刺激也看做是打破身体感受基线的新刺激,对所有刺激表征形成的普适机制进行了回答。每个外部/内部刺激都会产生特定身体状态,身体将特定身体状态传给大脑,在感知觉运动—情感脑区形成特定的身体感受地图,所有身体感受地图组成了身体图谱。当外部/内部刺激改变身体状态,在身体图谱中进行参照就产生相应感受和意识。全新刺激则产生新的身体感受,并增添到身体图谱中。(2)感受的产生,可以不需身体亲自参与,存在一种“as if”的模拟机制:感知到别人的动作和感受时,身体模拟出同样状态,从而产生相同感受和意识;“as if”模拟系统还能通过自我想象或言语来模拟自身身体状态。“as if”模拟系统已获得镜像神经元系统 MNS研究的广泛证实(Simonetti,2014)。
支持具身意识理论的大量实证研究散见于自主神经系统与意识、身体感知觉运动意识、情感意识、自我意识等四大领域,下面分别归纳介绍。
2.1 自主神经系统与意识
自主神经系统影响着意识体验。Bernstein(2015)通过对交感神经和副交感神经各自的基线强度和激活敏感度差异的探索,区分出了 4种类型的人,发现他们对环境/社会刺激和自身冲动的敏感性和抑制显著不同,造成了对外界和自身的感受差异,进而表现出不同的情感和认知状态。可惜没有探索其与意识的关系。于晓琳、张建保和王珏(2009)通过小波包能量和近似熵方法,研究了意识控制心率时脑电活动和心率变异性的变化规律,发现心率的减慢和加快,可通过意识对自主神经的主动调节进行控制。反之,Furman,Waugh,Bhattacharjee,Thompson和Gotlib (2013)探索心率的感知如何影响认知,发现有越多心率意识的人对感性图片有更强烈的情绪,能更好识别他人面部表情,更快学会躲避电击,这表明心率意识与意识敏感性相关。Sandman (2006)通过大量实验发现,心率的增强/减弱会提高/降低知觉敏感性和认知反应。
2.2 运动MNS与意识
镜像神经元系统(MNS)包括三大子系统(胡晓晴,傅根跃,施臻彦,2009):运动(运动前区—初级运动皮层M1—顶叶前部等)、情感(边缘系统杏仁核内侧颞叶—脑岛—扣带回等)、语言(运动前区—初级运动皮层 M1—布罗卡区等)。情感/语言MNS包含了运动MNS的一些脑区,情感/语言加工时除了激活各自的 MNS,还会激活运动 MNS(特别是运动前区)。但为了简明,本文将分开论述和考察三大子系统。
本文总结出运动MNS会产生四类意识:
(1)对外界/内部刺激的感知觉运动特征的本体感受和预测。本体感受器、各感知觉通道的初级皮层和运动前区是产生初级意识的脑区(Fogel,2011)。本体感受的产生包括刺激驱动和主动预测。Cole和Paillard (1995)发现,本体感受器受损患者的感知觉丧失;Gallagher (2003,1995)发现,被试会提前对外界刺激产生自上而下的预测,这一预测过程由本体感受器、各感知觉通道的初级皮层、运动前区、小脑和纹状体交互来实现(Legrand,2006;Castiello &Jeannerod,1991)。流畅性是内隐序列学习意识的重要因素(郭秀艳等,2008;Aizenstein et al.,2004),而刺激驱动和主动预测都可能是流畅性的来源。
(2)对外界/内部刺激意义和意图的意识。Simonetti (2014)综述相关研究发现,感知觉运动、动作意图、表情识别、语言和逻辑推理,都会激活运动MNS (运动前区—初级运动皮层—顶叶前部),该系统是模拟的关键网络。运动前区负责刺激意义,是高级意识产生及认知控制的脑区(Chong &Mattingley,2009)。Bonini等(2010)实验发现,有意图的动作对运动前区的激活大于无意图动作;Fogassi等(2005)实验发现,猕猴观察不同意图的动作,运动MNS激活模式不同。但是已有研究局限在动作意图领域,而内隐序列学习中刺激意义不明显:对刺激位置按键是纯反应时任务,没有明显而直接的生存/社会意义,其意义内隐存在于序列规则——前面的刺激位置决定后面的位置(其因果关系和趋势预测的属性类似动作意义和意图)。其意识是否正是来自于对内隐意义的获得呢?值得探索。
(3)对外界/内部刺激的动机和情感的意识。它由运动和情感MNS的交互实现。由于两者相对独立,情感MNS又是具身意识的重要内容,因此将在下一节详细介绍。
(4)对外界/内部刺激的语言表征,由运动/情感和语言 MNS的交互实现。语言表征(主观意识报告)是上述三类意识的测量指标之一。由于由语言 MNS负责加工的主观意识报告在内隐序列学习意识领域已有很多探索(全面而经典的探讨参见Rünger &Frensch,2010),且不是本文的新意和重点,因此不再详述。
Koziol,Budding和Chidekel (2012)综述相关研究,认为身体的各系统,在前额叶的感知觉联合皮层、运动前区和初级运动区产生了紧密连接,并且和认知控制系统、情感MNS连接,形成了多脑区交互。这使得上述四类较低级的感知觉运动意识,能交互形成复杂意识,并与认知控制和情感系统交互,产生受自上而下控制的高级意识(Fogel,2011,见图1)。
2.3 情感MNS与意识
情感 MNS和感知觉运动(包括运动 MNS)意识有何关系呢?Decety和Ickes (2009)的综述总结了感知觉运动和社会认知的加工过程:(1)刺激通过本体感受器进入初级感知觉皮层,进行初级刺激表征;(2)通过脑岛进入边缘系统和杏仁核,进行动机启动和情感估价;(3)间脑导水管(midbrain aquetube),动作反射准备;(4)运动 MNS,模拟产生运动意象和意义;(5)带着情感动机和运动意象的高级刺激表征进入前额叶,进行认知控制和行为选择,并通过脑岛自上而下作用于低级皮层。可见感知觉运动和情感两者在认知加工过程上有紧密的互动,感知觉运动加工从一开始就带有情感因素,因此情感MNS必然参与了对感知觉运动的主观感受和意识的产生过程,而且是在运动MNS发挥作用之前(Lindquist,Wager,Bliss-Moreau,Kober,&Barrett,2012)。Gu,Liu,van Dam,Hof和 Fan(2013)的 fMRI实验则发现所有感知觉运动脑区,都通过脑岛和情感MNS (边缘系统—杏仁核—内侧颞叶—扣带回前部)紧密联系。Gu,Hof,Friston和 Fan (2013)综述了脑岛的有关研究,发现脑岛与情感 MNS的其它脑区(边缘系统—杏仁核—内侧颞叶)、所有感知觉运动脑区(包括运动MNS)、认知控制脑区(前额叶—顶叶—扣带回)都有紧密联系,并据此提出脑岛和意识关系的模型假设(见图2)。因此脑岛很可能参与了所有意识的产生过程和随后的意识控制加工。

图1 内感受、身体图式和具身自我意识的神经网络关联(译自Fogel,2011)
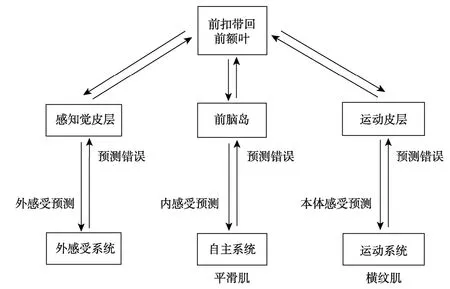
图2 前脑岛和意识关系模型假设(译自Gu,Hof,et al.,2013)
以上研究只能说明脑岛参与了多种意识过程,那么脑岛是不是情感意识的来源脑区呢?情感意识是指对情感的意识,诱发情感的原因可以是内部的心情、记忆、想象、自我认知,或者是躯体和内脏的感受,或者是外界的感知觉运动和社会刺激。Sridharan,Levitin和Menon (2008)用granger因果分析了信息流方向,发现在脑岛产生了自下而上和自上而下的信息交互,该交互被认为是意识产生的主要条件(Dehaene &Changeux,2011);并且是脑岛信息先流向扣带回。因此Sridharan等认为脑岛产生了情感意识“feeling”,然后将“feeling”传入扣带回、前运动区、前额叶等进行动作模拟、高级情感评估和认知控制,这些高级加工信息再自上而下回流脑岛,进行情感再加工和控制。这从大脑网络连接角度证明了脑岛而非扣带回是情感 MNS产生意识的关键脑区,虽然这还只是一种granger因果,不一定是真实因果。Gu等(2012)对大量的情感意识 fMRI研究做元分析发现,脑岛是连接感知觉运动和情感MNS的最重要脑区,它是产生情感意识的脑区,因为在各种不同认知任务中其激活程度都与情感意识程度相关;而非前扣带回(ACC),因为其激活程度只在某些特定任务中与情感意识程度相关,且可能仅仅是脑岛激活的结果。Picard (2013)发现癫痫使得脑岛激活增大,从而增强了情感意识,这从病理学的真实因果关系上证明了脑岛是情感意识的来源脑区。可见脑岛确实是产生情感意识的源发性脑区,而扣带回只是情感意识产生后被激活来参与意识控制。
内隐序列学习就是感知觉运动任务,被试对隐藏的序列规则会产生猜测、直觉、流畅、熟悉、记忆、规则等不同的主观感受和意识(Fu et al.,2010),它们是否来自情感 MNS特别是脑岛呢?或者某些主观意识来自情感MNS,另一些来自运动MNS及运动/情感MNS的交互?值得探索。
2.4 运动/情感MNS和自我网络三者交互与意识
运动/情感意识研究表明刺激加工包含了动机、情感、意义、预期等主观意识因素,这些都与自我意识有关,而自我意识主要由默认网络DMN负责。
DMN和意识相关,表现在三个方面:(1)意识障碍程度加深,DMN激活程度降低(Crone et al.,2011);(2)DMN在自我和社会认知中正激活,和自我意识正相关(Molnar-Szakacs &Uddin,2013);(3)DMN在与自我无关的外界认知任务中负激活,但与认知过程的意识漫游正相关(Christoff,Gordon,Smallwood,Smith,&Schooler,2009),与外界认知任务的意识的相关则有正有负。Stawarczyk,Majerus,Maquet和 D’Argembeau (2011)用 fMRI探讨了DMN和被试认知任务意识的关系:测量了任务相关且刺激相关、任务相关但刺激无关、任务无关但刺激相关、任务无关且刺激无关(意识漫游)四种意识状态,发现 DMN各子成分(特别是 PCC和MPFC)在与任务/刺激无关的后三种自变量水平下正激活,且随无关程度增加而递增。这表明即使在与自我无关的纯粹认知任务中,DMN也会产生来源于意识漫游的正激活。同时与任务紧密相关的意识,使得DMN的大部分区域,特别是PCC和 MPFC负激活;但是在小部分区域如左前下顶叶和右额内侧回产生正激活,表明任务意识与DMN存在复杂但紧密的相关。内隐序列学习意识就是认知任务的意识,和DMN会有紧密关联。关联方式可能有两种:序列规则与自我联系越强,意识程度越高(正相关);序列规则对自我打搅越高,越易获得意识(负相关)。
运动/情感 MNS既能独自产生主观感受,又通过脑岛—扣带回进行交互(图2),加上与自我系统 DMN、认知控制系统(额叶—顶叶—扣带回,Dehaene et al.,2011,详见1.1和图1)交互,则能产生新的高级意识。如 Carr,Iacoboni,Dubeau,Mazziotta和 Lenzi (2003)探究动作表征和共情的关系发现,运动皮层建立的动作内部表征调节着情感脑区激活模式,脑岛联结了动作表征的运动前区和情感体验的边缘系统。Molnar-Szakacs和Uddin (2013)综述了fMRI相关研究,证实自我意识网络 DMN和运动/情感 MNS在脑岛、后扣带回/楔前叶处重合(见图3)。
2.5 具身意识理论的总结
具身意识理论的关键进步是:把其它意识理论忽略的身体和感受作为意识产生的关键因素,为意识敏感性个体差异(Frith,2011)提供了具身解释。但是具身意识理论将意识定义为对身体状态的感受,关注意识产生的初级阶段,忽略了认知控制的高级意识阶段,全局工作平台理论可弥补该缺失;而反过来全局工作平台理论试次前随机波动的机制和原因,则可由MNS的激活模式来解释。
3 内隐序列学习意识的具身机制
从上述 1和 2的论述可见,全局工作平台理论、神经可塑性理论、新异刺激理论都应用于或产生于内隐序列学习意识研究,各自存在优点和缺陷;而具身意识理论提供了几乎所有意识产生的根本原因和具体机制——身体和感受因素,并能弥补其它三类意识理论的关键缺陷。但是遗憾的是内隐序列学习意识的研究,无论是实验报告、理论建构,还是综述文章,都未曾引入具身意识理论(张剑心等,2015;张润来等,2013;Rünger&Frensch,2010)。而内隐序列学习的序列规则意识,是在刺激变化和按键运动的感知觉运动具身认知中产生,且有情感、期待等情感具身和自我意识因素,其机制极可能就是具身的。因此本文首次将具身意识理论引入内隐序列学习意识,提出内隐序列学习意识是具身意识的假设,并探索其具身机制。

图3 MNS和DMN的重合(摘自Molnar-Szakacs &Uddin,2013)
3.1 内隐序列学习的运动具身机制
Penhune和Steele (2012)探查了经典的运动内隐序列学习(水平位置按键)脑区,发现小脑、初级运动皮层(M1)、基底节(主要是纹状体)都有激活。小脑负责形成优化的运动模块和运动控制及错误修正,初级运动皮层(M1)负责存储学习到的序列表征,纹状体负责刺激反应联结学习和位置预测,三者相对独立(尤其是纹状体)但在不同学习阶段频繁交互。Hardwick,Rottschy,Miall和Eickhoff(2013)对运动内隐序列学习已有研究的元分析发现,运动脑区(运动前区—初级运动皮层 M1—小脑认知区域)、初级躯体感觉区、纹状体都得到了激活,实际上已表明运动内隐序列学习本来就是具身的。Bo,Peltier,Noll和Seidler (2011)比较了两种表征内隐交替序列学习(按键/口头)和两种运动内隐交替序列学习(按键/口头)的差异,表征内隐序列学习是字母按照序列规则出现在屏幕中央,对字母做反应。结果发现:表征内隐序列和运动内隐序列在运动前区、M1区和纹状体都有相同程度的激活,而且表征内隐序列学习的内隐学习量居然还和小脑的纯运动区HVI正相关。这说明即使没有位置运动的纯粹表征内隐序列学习,仍然需要广泛的运动脑区。
可见内隐序列学习主要存在两个不同的学习系统:运动前区—初级运动皮层 M1—小脑 HVI区/小脑认知区域,负责序列的运动学习;纹状体则负责刺激—反应联结,及对下一步的预测(Penhune &Steele,2012)。前者和运动MNS (运动前区—初级运动皮层 M1—顶叶前部等)存在大量重合,实质上只差一步就能验证其具身机制;后者和情感 MNS有关键联系(见下一小节 3.2)。因此这两个学习机制都可能是具身的。
3.2 内隐序列学习意识的运动/情感具身本源
既然内隐序列学习存在两种机制,就有对应的两种意识产生机制。Wan等(2011)发现尾状核头部(包括纹状体)产生预测性的直觉;而流畅感(郭秀艳等,2008;Aizenstein et al.,2004)既可能由运动前区—初级运动皮层M1——小脑HVI区/小脑认知区域产生(运动表征流畅感),也可能由纹状体产生(预测流畅感)。已有研究证实内隐序列学习的这些机制能产生多种主观意识状态,如直觉、熟悉、流畅感、边缘意识、信心、元认知等,其行为和 ERP特征都存在差异(Fu,Bin,Dienes,Fu,&Gao,2013;Fu et al.,2010;Li,Hill,&He,2014;Dienes &Scott,2005;Norman et al.,2006,2007);产生的客观意识状态则有受控意识(包含减去排除,Fu et al.,2010)。可见内隐序列学习意识机制甚至不止两种,其中的主观意识状态就是主观感受体验,符合具身意识定义。
内隐序列学习意识的情感具身机制的研究则尚未见到,但有不少研究探索了与情感有关的人格特质与内隐序列学习意识的相关。Norman等(2006)探测了大五人格量表 NEO-PI-R (Costa &McCrae,1992)和内隐序列学习的关系,在RSI=0 ms时发现只有 openness/feeling情感开放性子量表与内隐序列学习反应时减少量、再认(内隐+外显成分)和包含任务(自动反应+受控意识)得分正相关。情感开放性指“高分者能够体验更深入,更丰富多样的情感状态,感觉到更强烈的快乐和不快乐等情绪。低分者则有些感觉迟钝,并且不相信那些感觉很重要(Costa &McCrae,1992)”,因此该研究表明内隐序列学习意识与情感体验有关。Norman等(2007)进一步采用 openness/feeling 子量表得分选出了高低情感开放性组,进行RSI=0 ms和1000 ms的内隐序列学习,却发现高低情感开放性组之间的内隐学习量和意识程度都没有差异,据此认为openness/feeling子量表对内隐序列学习意识的预测效度较低。Kaufman等(2010)测量了RSI=500 ms的内隐序列学习最后6个组段的主序列与非主序列的反应时差值与大五人格的相关,发现内隐序列学习量和直觉、开放性(美学、想象、情节、情感四个维度的总分)、冲动性相关。Kaufman等(2010)认为上述研究矛盾的原因可能是内隐序列学习只与经验开放性相关——美学、想象、情节三个开放性子维度测量的就是经验开放性,而与情感开放性不相关或低相关。张剑心等(2014)不同意 Kaufman等的观点,他们认为情感开放性能很好预测内隐序列学习意识,提出上述研究的矛盾只是由于各自 RSI和测量指标不同。因此张剑心等(2014)采用非主序列的学习量作为新指标,考察了高低情感开放性者在RSI=0 ms、250 ms、500 ms、750 ms和1000 ms的复杂概率内隐序列学习,发现高低情感开放性者在所有RSI的主序列和非主序列的学习都存在差异,并且分别在RSI=750 ms的再认任务(测量内隐+外显成分)和RSI=500 ms的旋转任务(测量边缘意识)差异显著,由此证明情感开放性能很好地预测内隐序列学习和意识。由于情感开放性测量的是人对情感的体验丰富度、深刻度和敏感性,因此张剑心等(2015)进而提出了内隐序列学习意识来源于对内隐序列规则的情感感受的理论。可惜的是这些还只是行为实验,尚缺乏电生理和脑成像研究;并且只证明了与情感开放性相关,尚未直接证明内隐序列学习意识就来源于情感体验。
要从脑机制上揭示两个认知过程相关,有个方法是证实两者的关键脑区存在重合,逻辑如Molnar-Szakacs和Uddin (2013)用运动MNS和默认网络DMN存在关键重合脑区来说明两者相关。因此本文首次将运动/情感 MNS和内隐序列学习意识脑区做比较:(1)运动MNS是多种意识(本体感受和预测、动作意图、情感、语言)产生的脑区,而内隐序列学习的运动前区—初级运动皮层M1—小脑HVI区/小脑认知区域与运动MNS大部分重合,意味着其是意识产生脑区。(2)情感MNS中扣带回前端负责处理预期性情感体验(Howell,2006)。内隐序列学习规则是前面位置预测后面位置,其关键脑区纹状体负责产生预测性直觉(Wan et al.,2011),而Lawrence (2012)认为直觉的本质就是具身意识。Destrebecqz和Peigneux (2005)发现内隐序列学习的受控意识,与纹状体和扣带回的连接程度显著正相关。这表明受控意识来自于扣带回对纹状体的预测性知识产生的预期情感体验。可惜该研究只选取了有限种子点做功能连接,没有考察整个情感MNS,但仍表明受控意识脑区与情感 MNS关键脑区之一扣带回有重合。Yang和Li (2012)采用fMRI研究了外显/内隐序列学习,发现外显学习激活脑岛,而内隐学习激活额叶—纹状体系统,表明脑岛是外显序列学习的脑区,而脑岛正是情感MNS的关键脑区。可惜该研究既没有考察情感MNS对外显序列学习的作用,如未考察情感 MNS的边缘系统等的激活是否能通过脑岛影响外显序列学习量;更没有探索脑岛是否是内隐序列学习意识产生的源发点之一,如只探索了脑岛对外显序列学习的作用,而外显序列学习在学习前直接给出了序列规则,没有意识产生过程,因此和内隐序列学习通过不断训练而产生对隐藏的序列规则的各种意识状态(包括完全外显),有本质差异。该研究还存在两个致命缺陷:一是它将内隐序列学习的所有被试合在一起平均,而这些被试对内隐序列规则的意识程度差异很大(Fu et al.,2013;张剑心等,2015),有的是完全内隐,有的则有受控意识,叠加平均当然就稀释了内隐序列学习意识对脑岛的激活;二是没有做意识测量阶段,因此无法得到客观/主观意识分数,既无法区分出内隐序列学习的不同意识程度的被试,更无法找到与意识相关的脑区。但是该研究毕竟证明外显序列学习(即对序列规则有完全的意识)与脑岛相关,那么内隐序列学习中产生的意识就很可能与脑岛相关,因此笔者正从上述两方面对该研究范式加以改进,以探索脑岛与内隐序列学习意识的关系。
综上所述,无论学习机制、具身意识定义还是加工脑区,内隐序列学习意识都与具身意识非常吻合。
4 未来研究展望
因此未来研究可通过实验来探索内隐序列学习意识是否就是一种具身意识,并考察其具体而特殊的具身机制;在此基础上,将具身意识理论与内隐序列学习意识的三个理论——全局工作平台理论、神经可塑性理论、新异刺激理论结合,互相补充完善。下面介绍本研究团队正在进行的三方面工作以及对未来研究的展望,期待能有幸与感兴趣的学者们交流合作。
4.1 证明内隐序列学习意识的运动/情感具身本源
内隐序列学习意识是对隐藏的序列规则的意识,相比于感知觉意识,它是更高级的规则意识,对人类学习和认知具有更重要的作用;相比于自我意识,它是对外界刺激规则的意识,对人类掌握外界事物的规律以对其加以应对和处理具有重要意义。已有具身意识研究集中在感知觉运动意识和自我身体意识,未探索过学习外部刺激的规则所产生的高级意识——如内隐序列学习规则意识;而在内隐序列学习意识领域,所有意识理论和研究又都忽略了身体因素。从上述2和3的论述可见,一方面身体体验(包括感知觉运动、情感和自我)是意识的本源;另一方面内隐序列学习意识正是在感知觉运动学习中产生的,包含了主观体验,并且与情感开放性有关。因此内隐序列学习意识的本源极可能就是一种具身意识。在上述3中只是通过比较两个领域已有研究的脑区重合性来推论,还不是直接证据。要获得直接证据必须要在内隐序列学习中证明运动/情感 MNS与意识相关甚至有因果关系。因此未来研究除了采用相关分析,更应该采用动态因果模型或 Granger因果来建立具有因果方向的大脑网络,探索运动/情感 MNS的信息流是否能够决定认知控制系统(前额叶—顶叶—扣带回)的激活程度(类似具身意识领域的研究Sridharan et al.,2008),以及这些脑区的连接程度与意识得分的相关性(类似内隐序列学习意识领域的研究 Destrebecqz &Peigneux,2005)。
另外,可采用相同的技术探索自我意识网络与内隐序列学习意识的关系——自我意识网络和运动/情感 MNS有关键重合之处,虽然不能认为自我意识网络就是 MNS的一部分,但是毕竟与MNS紧密相关,因此做这样的探索有助于揭示内隐序列学习意识的更多的具身机制。
4.2 证明全局工作平台理论、神经可塑性理论、新异刺激理论的具身基础
如果上一步证明了内隐序列学习意识确实存在具身机制,那么第二步就可以探索和澄清内隐序列学习意识的已有理论与具身意识理论的关系。全局工作平台理论、神经可塑性理论、新异刺激理论都忽略了身体因素,未来研究应该把身体因素引入到这三个理论。
全局工作平台理论认为某个试次能否激活全局工作平台,取决于试次前被试随机波动的主观状态(Aru et al.,2012),但是随机波动的原因并不清楚。未来研究可以分析试次前运动/情感 MNS以及自我意识网络 DMN对试次前随机波动的贡献,进而得到试次中全局工作平台激活的具身原因;而探索试次中运动/情感 MNS以及自我意识网络 DMN对意识的贡献将能够弥补全局工作平台理论忽略了刺激驱动因素的缺陷,并且把试次前主观驱动因素和试次中客观刺激驱动因素统合于具身机制。反过来,全局工作平台理论关注意识产生后对整个大脑认知控制网络的改变,具身意识理论则主要解释意识的起源,前者正好可以描述后者的意识产生后有什么特征,起什么作用,导致大脑网络发生什么变化。因此未来还可以采用大脑网络技术分析早期激活的运动/情感 MNS与中后期激活的认知控制系统之间的因果关系。
神经可塑性理论产生于内隐序列学习意识研究,认为随表征质量渐进增长,大脑网络被重塑从而产生独特持久的表征,形成意识。但是它将表征质量渐进增长的原因解释为训练,只是行为学上的描述性解释;具身意识理论则提供了训练导致表征质量渐进增长的机制性原因——运动/情感MNS在训练中不断被激活,形成了新的身体地图,从而打破原来的身体状态,产生了序列规则意识。未来研究应该考察随着内隐序列学习训练组块的增加,运动/情感 MNS是否发生着渐进变化,并且这种渐进变化是否是内隐序列学习表征质量(纹状体,Simon et al.,2011)和意识(扣带回,Destrebecqz &Peigneux,2005)渐进变化的源发性因素。
新异刺激理论也来自于内隐序列学习意识研究,认为恒常刺激意识的产生来自于新异刺激的意外事件效应。新异刺激理论几乎可以完全被具身意识理论替代,因为具身意识理论把恒常刺激都认为是打破原有身体状态的新刺激,使得理论解释的逻辑更加完备;并且比新异刺激理论更深一步,提出了新异刺激的影响发生在具身层面——产生新的身体状态地图。未来研究应该在新异刺激理论的已有研究中,证明新异刺激是否导致了运动/情感 MNS的改变,并且正是这种具身改变进一步导致了意识产生。
4.3 探索影响内隐序列学习具身意识的因素
内隐序列学习意识研究领域中,人格开放性和RSI是两大影响因素(张剑心等,2015)。张剑心等(2014)、Kaufman等(2010)、Norman等(2006)都证明人格开放性(或其中的情感开放性)影响内隐序列学习及其意识,并受反应刺激间隔 RSI调节。如果上两步证明了内隐序列学习意识确实存在具身机制,那么就可以推论人格开放性和 RSI可能正是通过具身机制来影响意识。未来研究可以探索人格开放性的对应脑区(女性主要在前额叶,男性在前额叶和扣带回,Sutin,Beason-Held,Resnick,&Costa,2009)与运动/情感MNS的连接模式和程度对意识的影响,并且考察 RSI如何调节人格开放性与具身意识关系。
郭秀艳,高妍,沈捷,王丽嘉,门卫伟,傅成,杨治良.(2008).同步内隐/外显序列学习:事件相关的 fMRI的初步研究.心理科学,31(4),887–891.
胡晓晴,傅根跃,施臻彦.(2009).镜像神经元系统的研究回顾及展望.心理科学进展,17(1),118–125.
黄建平,张剑心,刘电芝.(2014).内隐序列学习中转移组块的数量和位置效应.北京:中国科技论文在线.2014-05-12取自http://www.paper.edu.cn/releasepaper/content/201405-161.
于晓琳,张建保,王珏.(2009).意识控制心率过程中头皮脑电活动和心率变异性分析.中国医疗器械杂志,3 3,235–238.
张剑心,黄颀,张润来,刘电芝.(2015).内隐序列学习意识的理论、测量和影响因素.心理科学进展,23(5),793–805.
张剑心,武燕,陈心韵,刘电芝.(2014).高低情感开放性者概率内隐序列学习进程差异.心理学报,46(12),1793–1804.
张润来,刘电芝,张剑心,程昊旸.(2013).内隐学习意识性研究的两种理论取向:二分与渐进.心理科学,36(6),1517–1523.
Aizenstein,H.J.,Stenger,V.A.,Cochran,J.,Clark,K.,Johnson,M.,Nebes,R.D.,&Carter,C.S.(2004).Regional brain activation during concurrent implicit and explicit sequence learning.Cerebral Cortex,14(2),199–208.
Allen,M.,&Williams,G.(2011).Consciousness,plasticity,and connectomics:The role of intersubjectivity in human cognition.Frontiers in Psychology,2,20.
Aru,J.,Bachmann,T.,Singer,W.,&Melloni,L.(2012).Distilling the neural correlates of consciousness.Neuroscience&Biobehavioral Reviews,36,737–746.
Askenasy,J.,&Lehmann,J.(2013).Consciousness,brain,neuroplasticity.Frontiers in Psychology,4,412.
Atas,A.,Vermeiren,A.&Cleeremans,A.(2013).Repeating a strongly masked stimulus increases priming and awareness.Consciousness and Cognition,22,1422–1430.
Bernstein,W.M.(2015).A basic and applied model of the body–mind system.In G.H.E.Gendolla,M.Tops,&S.L.Koole (Eds.),Handbook of biobehavioral approaches to self-regulation(pp.397–418).New York:Springer Science Business Media.
Bo,J.,Peltier,S.J.,Noll,D.C.,&Seidler,R.D.(2011).Age differences in symbolic representations of motor sequence learning.Neuroscience Letters,504,68–72.
Boly,M.,Balteau,E.,Schnakers,C.,Degueldre,C.,Moonen,G.,Luxen,A.,… Laureys,S.(2007).Baseline brain activity fluctuations predict somatosensory perception in humans.Proceedings of the National Academy of Sciences of the United States of America,104,12187–12192.
Bonini,L.,Rozzi,S.,Serventi,F.U.,Simone,L.,Ferrari,P.F.,&Fogassi,L.(2010).Ventral premotor and inferior parietal cortices make distinct contribution to action organization and intention understanding.Cerebral Cortex,20(6),1372–1385.
Bower,M.,&Gallagher,S.(2013).Bodily affects as prenoetic elements in enactive perception.Phenomenology and Mind,4(1),78–93.
Carr,L.,Iacoboni,M.,Dubeau,M.-C.,Mazziotta,J.C.,&Lenzi,G.L.(2003).Neural mechanisms of empathy in humans:A relay from neural systems for imitation to limbic areas.Proceedings of the National Academy of Sciences of the United States of America,100,5497–5502.
Castiello,U.,&Jeannerod,M.(1991).Measuring time to awareness.Neuroreport,2,797–800.
Chong,T.T.J.,&Mattingley,J.B.(2009).Automatic and controlled processing within the mirror neuron system.In J.A.Pineda (Ed.),Mirror neuron systems(pp.213–233).New York:Humana Press.
Christoff,K.,Gordon,A.M.,Smallwood,J.,Smith,R.,&Schooler,J.W.(2009).Experience sampling during fMRI reveals default network and executive system contributions to mind wandering.Proceedings of the National Academy of Sciences of the United States of America,106,8719–8724.
Cleeremans,A.(2007).Consciousness:The radical plasticity thesis.Progress in Brain Research,168,19–33.
Cleeremans,A.(2011).The radical plasticity thesis:How the brain learns to be conscious.Frontiers in Psychology,2,86.
Cleeremans,A.,&Jiménez,L.(2002).Implicit learning and consciousness:A graded,dynamic perspective.In R.M.French &A.Cleeremans (Eds.),Implicit learning an d consciousness:An empirical,computational and philosophical consensus in the making?(pp.1–40).Hove,UK:Psychology Press.
Cole,J.,&Paillard,J.(1995).Living without touch and peripheral information about body position and movement:Studies upon deafferented subjects.In J.Bermúdez,A.Marce,&N.Eilan (Eds.),The body and the self(pp.245–266).Cambridge,Mass:MIT.
Costa,P.T.,&McCrae,R.R.(1992).Revised N EO Personality Inventory (NEO-PI-R) and NEO Five-Factor Inventory (NEO-FFI).Professional Manual.Odessa,FL:Psychological Assessment Resources.
Crone,J.S.,Ladurner,G.,Höller,Y.,Golaszewski,S.,Trinka,E.,&Kronbichler,M.(2011).Deactivation of the default mode network as a marker of impaired consciousness:An fMRI study.Plos One,6(10),26373.
Davidson,R.J.,&McEwen,B.S.(2012).Social influences on neuroplasticity:Stress and interventions to promote well-being.Nature Neuroscience,15,689–695.
Decety,J.,&Ickes,W.(2009).The social neuroscience of empathy.Cambridge:MIT Press.
Dehaene,S.,&Changeux,J.P.(2011).Experimental and theoretical approaches to conscious processing.Neuron,70,200–227.
Dehaene,S.,Changeux,J.P.,&Naccache,L.(2011).The global neuronal workspace model of conscious access:From neuronal architectures to clinical applications.In S.Dehaene &Y.Christen (Eds.),Characterizing consciousness:From cognition to the clinic? Research and perspectives in neurosciences(pp.55–84).Berlin Heidelberg:Springer-Verlag.
Dehaene,S.,Changeux,J.P.,Naccache,L.,Sackur,J.,&Sergent,C.(2006).Conscious,preconscious,and subliminal processing:A testable taxonomy.Trends in Cognitive Sciences,10(5),204–211.
Dehaene,S.,Charles,L.,King,J.R.,&Marti,S.(2014).Toward a computational theory of conscious processing.Current Opinion in Neurobiology,25,76–84.
Del Cul,A.,Baillet,S.,&Dehaene,S.(2007).Brain dynamics underlying the nonlinear threshold for access to consciousness.PLoS Biology,5,e260.
Destrebecqz,A.,&Cleeremans,A.(2001).Can sequence learning be implicit? New evidence with the process dissociation procedure.Psychonomic Bulletin &Review,8(2),343–350.
Destrebecqz,A.,&Cleeremans,A.(2003).Temporal effects in sequence learning.In L.Jiménez (Ed.),Attention and implicit learning(pp.181–213).Amsterdam:John Benjamins Publishing Company.
Destrebecqz,A.,&Peigneux,P.(2005).Methods for studying unconscious learning.Progress in Brain Research,150,69–80.
Dienes,Z.,&Scott,R.(2005).Measuring unconscious knowledge:Distinguishing structural knowledge and judgment knowledge.Psychological Research,69,338–351.
Fogassi,L.,Ferrari,P.F.,Gesierich,B.,Rozzi,S.,Chersi,F.,&Rizzolatti,G.(2005).Parietal lobe:from action organization to intention understanding.Science,308,662–667.
Fogel,A.(2011).Embodied awareness:Neither implicit nor explicit,and not necessarily nonverbal.Child Development Perspectives,5(3),183–186.
Frith,C.D.(2011).What brain plasticity reveals about the nature of consciousness:Commentary.Frontiers i n Psychology,2,87.
Fu,Q.F.,Bin,G.Y.,Dienes,Z.,Fu,X.L.,&Gao,X.R.(2013).Learning without consciously knowing:Evidence from event-related potentials in sequence learning.Consciousness and Cognition,22(1),22–34.
Fu,Q.F.,Dienes,Z.,&Fu,X.L.(2010).Can unconscious knowledge allow control in sequence learning?Consciousness and Cognition,19(1), 462–474.
Furman,D.J.,Waugh,C.E.,Bhattacharjee,K.,Thompson,R.J.,&Gotlib,L.H.(2013).Interoceptive awareness,positive affect,and decision making in Major Depressive Disorder.Journal of Affective Disorders,151(2),780–785.
Gaillard,R.,Dehaene,S.,Adam,C.,Clémenceau,S.,Hasboun,D.,Baulac,M.,… Naccache,L.(2009).Converging intracranial markers of conscious access.PLoS Biology,7,e1000061.
Gallagher,S.(2005).How the body shapes the mind.Oxford:Oxford University Press.
Gallagher,S.(1995). Body schema and intentionality.In:J.L.Bermúdez,A.Marcel,&N.Eila (Eds.),The body and the self(pp.225–244).Cambridge:The MIT Press.
Gallagher,S.(2003).Bodily self-awareness and object-perception.Theoria et Historia Scientiarum:International Journal for Interdisciplinary Studies,7(1),53–68.
Gu,X.S.,Gao,Z.X.,Wang,X.C.,Liu,X.,Knight,R.T.,Hof,P.R.,&Fan J.(2012).Anterior insular cortex is necessary for empathetic pain perception.Brain,135,2726–2735.
Gu,X.S.,Hof,P.R.,Friston,K.J.,&Fan,J.(2013).Anterior insular cortex and emotional awareness.TheJournal of Comparative Neurology,521,3371–3388.
Gu,X.S.,Liu,X.,van Dam,N.T.,Hof,P.R.,&Fan,J.(2013).Cognition–emotion integration in the anterior insular cortex.Cerebral Cortex,23,20–27.
Hardwick,R.M.,Rottschy,C.,Miall,R.C.,&Eickhoff,S.B.(2013).A quantitative meta-analysis and review of motor learning in the human brain.NeuroImage,67,283–297.
Howell,R.J.(2006).Self-knowledge and self-reference.Philosophy and Phenomenological Research,72,44–70.
Imamoglu,F.,Kahnt,T.,Koch,C.,&Haynes,J.D.(2012).Changes in functional connectivity support conscious object recognition.NeuroImage,63(4),1909–1917.
Kaufman,S.B.,DeYoung,C.G.,Gray,J.R.,Jiménez,L.,Brown,J.,&Mackintosh,N.(2010).Implicit learning as an ability.Cognition,116,321–340.
King,J.R.,Faugeras,F.,Gramfort,A.,Schurger,A.,Karoui,I.,Sitt,J.D.,… Dehaene,S.(2013).Single-trial decoding of auditory novelty responses facilitates the detection of residual consciousness.NeuroImage,83,726–738.
Koziol,L.F.,Budding,D.E.,&Chidekel,D.(2012).From movement to thought:Executive function,embodied cognition,and the cerebellum.The Cerebellum,11,505–525.
Kupers,R.,Pietrini,P.,Ricciardi,E.,&Ptito,M.(2011).The nature of consciousness in the visually deprived brain.Frontiers in Psychology,2,19.
Lawrence,R.L.(2012).Intuitive knowing and embodied consciousness.New Directions for Adult and Continuing Education,134,5–13.
Legrand,D.(2006).The bodily self:The sensori-motor roots of pre-reflective self-consciousness.Phenomenology and the Cognitive Sciences,5,89–118.
Li,Q.,Hill,Z.,&He,B.J.(2014).Spatiotemporal dissociation of brain activity underlying subjective awareness,objective performance and confidence.The Journal of Neuroscience,34(12),4382–4395.
Li,W.,Howard,J.D.,Parrish,T.B.,&Gottfried,J.A.(2008).Aversive learning enhances perceptual and cortical discrimination of indiscriminable odor cues.Science,319,1842–1845.
Lindquist,K.A.,Wager,T.D.,Bliss-Moreau,E.,Kober,H.,&Barrett,L.F.(2012).What are emotions and how are they created in the brain?Behavioral and Brain Sciences,35(3),172–202.
Molnar-Szakacs,I.,&Uddin,L.Q.(2013).Self-processing and the default mode network:Interactions with the mirror neuron system.Frontiers in Human Neuroscience,7,571.
Noë,A.(2004).Action in perception.Cambridge,MA:MIT Press.
Norman,E.(2010).“The Unconscious” in current psychology.European Psychologist,15(3),193–201.
Norman,E.,Price,M.C.,&Duff,S.C.(2006).Fringe consciousness in sequence learning:The influence of individual differences.Consciousness and Cognition,15(4),723–760.
Norman,E.,Price,M.C.,Duff,S.C.,&Mentzoni,R.A.(2007).Gradations of awareness in a modified sequence learning task.Consciousness and Cognition,16,809–837.
Penhune,V.B.&Steele,C.J.(2012).Parallel contributions of cerebellar,striatal and M1 mechanisms to motor sequence learning.Behavioural Brain Research,226(2),579–591.
Picard,F.(2013).State of belief,subjective certainty and bliss as a product of cortical dysfunction.Cortex,49(9),2494–2500.
Ray,S.,&Maunsell,J.H.R.(2010).Differences in gamma frequencies across visual cortex restrict their possible use in computation.Neuron,67,885–896.
Rose,M.,Haider,H.,&Büchel,C.(2010).The emergence of explicit memory during learning.Cerebral Cortex,20(12),2787–2797.
Rünger,D.(2012).How sequence learning creates explicit knowledge:The role of response–stimulus interval.Psychological Research,76,579–590.
Rünger,D.,&Frensch,P.A.(2008).How incidental sequence learning creates reportable knowledge:The role of unexpected events.Journal of Experimental Psychology:Learning,Memory,and Cognition,34(5),1011–1026.
Rünger,D.,&Frensch,P.A.(2010).Defining consciousness in the context of incidental sequence learning:Theoretical considerations and empirical implications.Psychological Research PRPF,74(2),121–137.
Sadaghiani,S.,Hesselmann,G.,&Kleinschmidt,A.(2009).Distributed and antagonistic contributions of ongoing activity fluctuations to auditory stimulus detection.The Journal of Neuroscience,29,13410–13417.
Sandman,C.A.(2006).Cardiac afferent influences on consciousness.In R.J.Davison (Ed.),Consciousness and self-regulation:Advances in research and theory(pp.55–85). US:Springer.
Schröter,M.S.,Spoormaker,V.I.,Schorer,A.,Wohlschläger,A.,Czisch,M.,Kochs,E.F.,… Rüdiger,I.(2012).Spatiotemporal reconfiguration of large-scale brain functional networks during Propofol-induced loss of consciousness.The Journal of Neuroscience,32(37),12832–12840.
Schwager,S.,Rünger,D.,Gaschler,R.,&Frensch,P.A.(2012).Data-driven sequence learning or search:What are the prerequisites for the generation of explicit sequence knowledge?Advances in Cognitive Psychology,8(2),132–143.
Schwiedrzik,C.M.,Singer,W.,&Melloni,L.(2009).Sensitivity and perceptual awareness increase with practice in metacontrast masking.Journal of Vision,9,18.1–18.18.
Shapiro,L.K.(2014).The Routledge handbook of embodied cognition(pp.9–17).London:Routledge.
Simon,J.R.,Stollstorff,M.,Westbay,L.C.,Vaidya,C.J.,Howard,J.H.,Jr.,&Howard,D.V.(2011).Dopamine transporter genotype predicts implicit sequence learning.Behavioural Brain Research,216,452–457.
Simonetti,N.(2014).Neurosciences and philosophy of mind:A reductive interpretation of the “Mirror Neurons System”(MNS).Research in Psychology and Behavioral Sciences,2(2),24–42.
Sridharan,D.,Levitin,D.J.,&Menon,V.(2008).A critical role for the right fronto-insular cortex in switching between central-executive and default-mode networks.Proceedings of the National Academy of Sciences of the United States of America,105,12569–12574.
Stawarczyk,D.,Majerus,S.,Maquet,P.,&D’Argembeau,A.(2011).Neural correlates of ongoing conscious experience:Both task-unrelatedness and stimulus-independence are related to default network Activity.PLoS One,6(2),e16997.
Sutin,A.R.,Beason-Held,L.L.,Resnick,S.M.,&Costa,P.T.(2009).Sex differences in resting-state neural correlates of openness to experience among older adults.Cerebral Cortex,19,2797–2802.
Thompson,E.(2007).Mind in life:Biology,phenomenology,and the sciences of mind.Cambridge,MA:Harvard University Press.
Thompson,E.,&Varela,F.J.(2001).Radical embodiment:Neural dynamics and consciousness.Trends in Cognitive Sciences,5(10),418–425.
Tononi,G.(2008).Consciousness as integrated information:A provisional manifesto.Biological Bulletin,215,216–242.
van den Heuvel,M.P.,Kahn,R.S.,Goñi,J.,&Sporns,O.(2012).High-cost,high-capacity backbone for global brain communication.Proceedings of the National Academy of Sciences of the United States of America,109(28),11372–11377.
van den Heuvel,M.P.,&Sporns,O.(2011).Rich-club organization of the human connectome.The Journal of Neuroscience,31,15775–15786.
Wan,X.H.,Nakatani,H.,Ueno,K.,Asamizuya,T.,Cheng,K.,&Tanaka,K.(2011).The neural basis of intuitive best next-move generation in board game experts.Science,331,341–346.
Wessel,J.R.,Haider,H.,&Rose,M.(2012).The transition from implicit to explicit representations in incidental learning situations:More evidence from high-frequency EEG coupling.Experimental Brain Research,217,153–162.
Wyart,V.,Dehaene,S.,&Tallon-Baudry,C.(2012).Early dissociation between neural signatures of endogenous spatial attention and perceptual awareness during visual masking.Frontiers in Human Neuroscience,6,16.
Yang,J.,&Li,P.(2012).Brain networks of explicit and implicit learning.PLoS One,7(8),e42993.
