心理干预的优先获益者
——来自发展可塑性的研究证据*
2016-02-01闫玉朋江光荣
闫玉朋 夏 勉 江光荣
(华中师范大学心理学院暨青少年网络心理与行为教育部重点实验室,湖北省人的发展与心理健康重点实验室,武汉 430079)
进化生态学、行为发展、生活史理论、分子生物学和流行病学在内等许多领域的研究发现,同一个基因型的个体因环境差异的影响可能发展出不同的表现型(Bateson et al.,2004),即个体具有调整其发展以应对环境条件的能力,这就是发展可塑性(Moczek et al.,2011)。例如,与处于有利条件下的女孩相比,童年早期处于不利环境中的女孩青春期的初次性行为的年龄会更早 (Belsky,2007),较早的初次性行为就是女孩应对童年不利条件而发展出的表现型,因为初次性行为提前可提高基因传播的可能性,避免个体在繁衍之前死于不利环境。
一些研究发现,个体受环境影响的程度不同,即发展可塑性存在个体差异(Ellis,Boyce,Belsky,Bakermans-Kranenburg,&van IJzendoorn,2011)。个体间发展可塑性的差异是指“不同的个体对于自身生活经验及环境的敏感性存在差异,从而导致不同的发展结果”(Belsky,1997,引自张卫,甄霜菊,2013)。在发展可塑性的个体差异的研究中,早期研究多集中于消极环境,并提出了个体受损于消极环境的素质-压力(diathesis-stress)模型(Monroe&Simons,1991)。后来,学者们不满于对消极环境的过于关注,加上受到进化推理思想的启发,先后提出两个涉及积极环境的新模型——差别易感性(differential susceptibility)(也有国内学者将之译为“不同易感性”(张文新,王美萍,曹丛,2012))模型(Belsky,Bakermans-Kranenburg,&van IJzendoorn,2007)和优势敏感性(vantage sensitivity)模型(Pluess&Belsky,2013;Manuck&McCaffery,2014)。这两个模型尤其关注个体在积极环境中的发展。在某种程度上,心理干预属于积极环境,而且因其作为实验条件的便利性,逐渐成为研究积极环境下个体发展可塑性的热点。其中一些干预研究表明具备某些特征的特定当事人更容易从心理干预中获益,这些特征涵盖人格和气质、基因以及生理方面的特征。
1 差别易感性模型与优势敏感性模型
1.1 差别易感性模型
根据进化论的观点,在不能预测下一代环境如何的情况下,为了使繁殖适合度(reproductive fitness)最大化,即提高基因传播的可能性,亲代采取了“两头下注(bets-hedging)”的策略,“下注”的方式为,使一些子代具备某种易感特征从而使其易感于未来的积极环境,使另一些子代不具备易感特征从而避免其易感于未来的消极环境(Ellis et al.,2011)。这种策略导致同一亲代的不同子代对环境具备不同的易感性,表现为同一个家庭内的兄弟姐妹对积极环境和消极环境有不同的反应性(Belsky et al.,2007;张卫,甄霜菊,2013)。值得注意的是,未来的环境未必正好与个体的易感特征相匹配。那些被“下注”为易感特征的个体,未来若生存在消极环境中,则其可能会受损最大;那些被“下注”为不具备易感特征的个体,未来若生活在积极环境中,则难以有所获益。但这样仍使得同一亲代的子代生活在与之相匹配的环境中的几率被最大化了,由此看来,“两头下注”的策略是传播基因的最佳策略。进化思想的这些理论催生了差别易感性模型。
差别易感性模型指具备某些特征的个体在积极环境中更易获益,在消极环境中则更易受损,积极环境中易获益的一面和消极环境中易受损的一面分别被称为差别易感性的“光明面(light side)”和“黑暗面(dark side)”,而不具备这些特征的个体在积极环境中并不更易获益,在消极环境中也并不更易受损(Belsky&Pluess,2009)。而后的很多研究也验证了这个模型(Bakermans-Kranenburg&van IJzendoorn,2011;Belsky&Pluess,2009,2013),即差别易感性特征可调节个体所处的环境与其发展结果之间的关系。例如,在富裕、温暖、和睦的家庭中成长的高敏感性儿童,相对于同等条件下的低敏感性儿童,能够发展出更高水平的社交技能;若其生活在混乱、暴力、贫困的家庭中,则有可能比同等条件下的低敏感性儿童社交技能更差。也就是说,敏感性调节了儿童成长的家庭环境与其社交技能之间的关系。
1.2 优势敏感性模型
同样基于进化推理,有研究者认为,有了聚焦于个体在消极环境中受损的素质-压力模型,也有了同时关注个体在消极环境中受损和在积极环境中获益的差别易感性模型,理应提出一个聚焦于个体在积极环境中获益的概念,表示个体在积极环境中的获益程度的差异性和多样性(Manuck&McCaffery,2014)。于是,Manuck及其合作者提出了优势敏感性模型,但他们最初并未对优势敏感性作出过多说明,只是指出它与素质-压力模型相对,表明在基因与环境的交互作用中,遗传变量调节了个体在积极环境中的获益(Sweitzer et al.,2013)。
Pluess和Belsky(2013)在与Manuck谈话的基础上,扩展了优势敏感性的含义,“某些个体对其所接触的积极环境反应更为敏感、积极”,在消极环境中也并不更易受损。例如,高智商的儿童在良好的教育条件下会有更高的学业成就,在不良的教育条件下学业成就也不会更低。与此同时,Pluess和Belsky(2013)提及一个与优势敏感性相对应的概念——“优势抵抗(vantage resistance),指具备某些特征的个体不能从积极影响中获益的倾向。例如,低智商的儿童在良好的教育条件下也难以获得优秀的学业成就。
差别易感性模型和优势敏感性模型均表现了个体在积极环境中获益的差异性,二者在消极环境方面则有所区别,优势敏感性模型表示个体在消极环境中不受损,而差别易感性模型表示个体在消极环境中受损具有差异性,见图1。
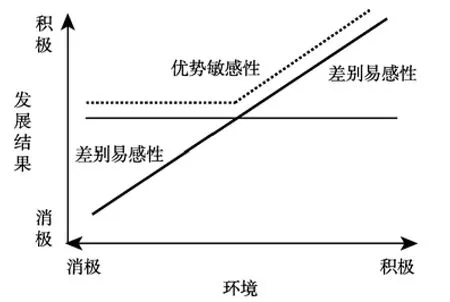
图1 差别易感性模型与优势敏感性模型
2 相关心理干预证据
为了一反素质-压力模型在心理健康领域的统治地位,差别易感性模型与优势敏感性模型对积极环境尤其关注。心理干预作为一种特殊的积极环境,便于操控,而且有一些作为实验研究所具备的特点,优于以往的相关研究:(1)个体特征与环境(心理干预)相互独立;(2)测量误差减少;(3)统计效力高(Bakermans-Kranenburg&van IJzendoorn,2015),因此心理干预研究越来越受到研究者的青睐。
2.1 差别易感性模型相关的心理干预研究
差别易感性模型相关的研究多集中于教养与儿童气质的交互作用(Temperament×Parenting)和遗传与环境交互作用(Gene×Environment)两个方面(Ellis et al.,2011),国内也有一些研究在遗传与环境交互作用方面检验了差别易感性模型(曹丛等,2014;王美萍,张文新,陈欣银,2015;张璠,陈杰,李新影,2015)。由于前述的原因,心理干预研究不断涌现。van IJzendoorn和Bakermans-Kranenburg(2012)曾对2011年12月前的差别易感性模型相关的干预研究进行评论,其中心理干预研究有7篇;2015年,期刊《Development and Psychopathology》为差别易感性模型引发的干预研究开辟了特辑(Belsky&van IJzendoorn,2015),其中心理干预研究有6篇。这些研究有的目的在于检验差别易感性模型,检验“黑暗面”意味着检验某个目标特征是否是问题行为或心理病理的成因之一,检验“光明面”意味着检验某一目标特征是否使个体更易于从心理干预中获益;有的并未做严格的检验。但这两类均致力于发现是否具备某个或某些特征的个体从心理干预中获益更多。差别易感性模型相关的这些心理干预研究按照差别易感性标记(marker)1标记(marker),来自于遗传学领域,遗传标记指在遗传分析中用作标记的基因,本文中出现的“标记”可理解为标志,通常为某一个体特征,如高感知加工敏感性这一个体特征就是优势敏感性的标记之一。(Belsky et al.,2007;Belsky&Pluess,2009)进行分类,可分为遗传类、气质类和生理类。下面分别从遗传类、气质类和生理类的特征介绍相关的研究。
2.1.1 遗传类特征的干预研究
有关差别易感性的遗传标记的研究较多,主要集中于对多巴胺受体D4(DRD4)基因区和5-羟色胺转运体(5-HTTLPR)基因区的检验。有一些研究验证了差别易感性模型。
现有的研究发现,DRD4-7重复等位基因是差别易感性的遗传标记。首先,较早的研究发现,该基因型被验证为易从消极环境中受损的特质,携带DRD4-7重复等位基因的婴幼儿若有一个不敏感的母亲,则表现出更多的外化问题行为(Bakermans-Kranenburg&van IJzendoorn,2006)。其后的研究将具有高水平外化问题行为的学步儿童随机分为两组,干预组为促进积极教养及敏感性惩戒的录像反馈干预,共6次;控制组则收到与干预组相同干预时间的6次电话援助。结果表明,对于携带DRD4-7重复等位基因的儿童,干预是有效的;对于非此基因型者,干预是无效的,即携带此基因的高水平外化问题行为的儿童从干预中有最大的获益(Bakermans-Kranenburg,van IJzendoorn,Pijlman,Mesman,&Juffer,2008)。因此可以证明携带DRD4-7重复等位基因的儿童易从消极环境中受损,同样易从积极环境中获益。这个结果证明了差别易感性模型的合理性。
Beach,Brody,Lei和Philibert(2010)将研究对象扩展到青少年,研究了有物质滥用问题的非裔美国青少年,同样证明DRD4-7重复等位基因为差别易感性标志。在干预组中,携带DRD4-7重复等位基因的青少年的物质使用行为更少,而控制组中携带DRD4-7重复等位基因的青少年报告了物质使用行为的增多。这一次在同一个研究中验证了差别易感性模型。Albert等人(2015)同样在一个研究中验证了差别易感性模型,他们追踪了270名欧裔美国人,从他们上幼儿园到25岁,干预为包括儿童社交技能训练、教养训练和亲子互动会谈等在内的快速轨道(Fast Track)干预,长达10年,结果发现,在小学和中学追踪时,参加干预的被试中携带糖皮质醇激素受体基因单核苷酸多态区rs10482672-A等位基因拷贝的儿童外化问题行为更少,而控制组的此基因型携带者有更多的外化问题行为,而且这个遗传与干预的交互作用对他们25岁时的遗传与干预的交互作用有中介作用。
然而,有一些研究由于没对个体在消极环境下的表现进行调查,因此没能明确验证差别易感性模型,但同样发现了携带某种基因型(促肾上腺素皮质激素释放激素受体1(CRHR1)TAT单体型0重复基因,以易感基因构建的多基因易感分数高,0或1个SCL6A3/DAT1-10重复等位基因)的特定群体(贫困年轻母亲,一年级低社会经济地位儿童,ADHD儿童)更易从心理干预(人际心理治疗,以教室为中心的干预,行为教养训练)中获益(抑郁症状更少,首次抽烟年龄延迟,症状减少)(Cicchetti,Toth,&Handley,2015;Musci et al.,2015;van den Hoofdakker et al.,2012)。也有一些研究没能验证差别易感性模型,也没发现特定基因区(多巴胺能(DRD2,DRD4,ANKK1)和γ-氨基丁酸能(GABRG1,GABRA2)基因区,5-HTTLPR基因区和DRD4基因区)调节了心理干预(援助非裔美国家庭项目,亲子心理治疗和心理教育性教养干预)对特定群体的效果(非裔美国青少年的酒精使用,受虐待儿童依恋安全性)(Brody,Chen,&Beach,2013;Cicchetti,Rogosch,&Toth,2011)。
以上研究大都关注遗传与干预两方面的交互作用,也有一些研究考虑了个体的生活环境,考察了三方面的交互作用。有的研究验证了差别易感性模型,如Schlomer等人(2015)发现对于生活在高水平母亲敌意下的11~16岁的高加索青少年,参与促进学校–社区–大学合作关系以提升心理弹性(PROmoting School-community-university Partnerships to Enhance Resilience,PROSPER)项目的干预后,DRD4-7+(7+表示大于等于7)重复等位基因携带者攻击行为减少,而此基因型个体在控制组则表现出最多的攻击行为。有的研究虽没有明确验证差别易感性模型,也发现生活在特定环境(高家庭风险,高母亲卷入)中携带特定基因型(DRD4-7+重复等位基因,至少1个DRD4-7重复等位基因)的青少年从心理干预(形成中的成年人干预项目,PROSPER干预项目)中获益更大(药物使用显著减少,饮酒风险降低)(Brody,Yu,&Beach,2015;Cleveland et al.,2015)。
基于差别易感性模型不断出现的随机对照实验,van IJzendoorn和Bakermans-Kranenburg(2015)进行了总的元分析和分类的元分析,其中的宏试验(macrotrials)所包含的研究与上述心理干预研究基本吻合,对11个宏试验的元分析结果表明易感基因携带者的干预有效性显著高于非易感基因携带者。
2.1.2 气质类特征的心理干预研究
检验差别易感性模型的最早的心理干预就来自气质方面的研究。Blair(2002)对婴幼儿健康和发展项目(Infant Health and Development Program,IHDP)的分析,检验了低出生体重早产婴儿在婴幼儿期的消极情绪性与儿童社会性发展和认知发展的关系。干预组提供的干预包括父母支持和高品质的教育性刺激;控制组只追踪而不做任何处理。结果发现,与干预组低水平消极情绪性儿童相比,干预组中高水平消极情绪性婴幼儿在行为方面和智力方面均表现出了最大的获益;在控制组中,高水平消极情绪性儿童则表现了更多的外化问题行为和内化问题行为。
Scott和 O’Connor(2012)采用 SPOKES 试验的数据,109名筛选出来的高对立水平的5~6岁儿童被随机分配到干预组或控制组,干预为时长12周的教养干预——顽劣年纪系列(Incredible Years,IY)学龄项目,结果表明,干预组中,以易激惹为显著特点的情绪紊乱型(Emotionally-Dysregulated type)儿童与控制组情绪紊乱型儿童相比,品行问题显著减少,处理效应量为0.84,远高于干预组顽固型(Headstrong type)儿童相应的效应量0.20,即表明情绪紊乱型儿童从干预中获益更大。干预前的数据表明顽固型儿童与情绪紊乱型儿童的行为问题与父母教养的相关系数差异虽不显著(p<0.05),但其关系模式表明情绪紊乱型儿童对积极教养和消极教养均有更强的敏感性。结合上述两方面的数据分析,研究者认为该研究验证了差别易感性模型。
气质方面也有研究没能验证差别易感性模型。Anzman-Frasca,Stifter,Paul和 Birch(2014)调查了婴幼儿消极性气质是否调节了抚慰/睡眠预防干预(Soothe/Sleep intervention)对婴幼儿情绪调节和肥胖的干预效应,结果未发现调节效应。
2.1.3 生理类特征的心理干预研究
到2015年4月为止,生理方面相关的心理干预研究仅有1篇(Gatzke-Kopp,Greenberg,&Bierman,2015),关注的目标特征是副交感神经反应性,以呼吸性窦性心律不齐为测量指标,干预为促进备择思维策略(PROmoting Alternative Thinking Strategies,PATHS)项目,干预对象为高水平攻击性和破坏性行为的儿童,结果发现对恐惧低反应性的儿童有更高的改善。但该研究并未对差别易感性模型做较为明确的检验。
2.2 优势敏感性的相关心理干预研究
优势敏感性模型的验证与差别易感性模型的意义不尽一致。与差别易感性模型的“光明面”得到验证的意义相似,优势敏感性模型的验证同样表明具备某个特征的个体更能从心理干预中获益;与差别易感性模型的“黑暗面”得到验证的意义相反,优势敏感性模型在消极环境中得到验证意味着某些特征对个体在消极环境中有保护性作用。
优势敏感性模型的提出较晚,因此检验研究较少(Brett et al.,2015;Carlson,Mendle,&Harden,2014;Nederhof,Belsky,Ormel,&Oldehinkel,2012;Pluess&Boniwell,2015)。到2015年4月为止,明确检验优势敏感性的心理干预研究只有一个。而有一个检验差别易感性模型的研究,其结果更符合优势敏感性模型(Cassidy,Woodhouse,Sherman,Stupica,&Lejuez,2011)。这两个研究检验的特征属于人格和气质类。
2.2.1 人格和气质类特征的心理干预研究
Pluess和Boniwell(2015)分析了英国伦敦一所女子学校实施的儿童运动、游戏和娱乐活动(Sport,Play,and Active Recreation for Kids,SPARK)心理弹性项目的数据。干预对象为166个11岁的女童,旨在预防这些高风险儿童(贫困地区的女孩)的抑郁。该项目以认知行为疗法和积极心理学的概念为基础,由经过专业心理学家训练的本校教师实施,总共12次,每次1个小时,跨越3~4个月。研究者对其进行了1年的追踪。结果表明,干预组整体与只进行了后测的控制组(n=197)在12个月后的抑郁无显著差异,然而干预组高感知加工敏感性(high Sensory-Processing Sensitivity)女童的抑郁分数显著低于控制组和低感知加工敏感性的女孩,表明高感知加工易感性女童从干预中获益最大。而且干预组前后测数据表明感知加工敏感性与抑郁分数无关,即高感知加工敏感性的儿童也并不更易从消极环境中受损,验证了优势敏感性模型。
Cassidy等人(2011)本来的目的在于检验差别易感性模型,他们的研究发现,接受安全圈家访干预(The Circle of Security-Home Visiting-4 Intervention,COS-HV4)的儿童中,高易激惹儿童的依恋安全性有显著提升,中度易激惹儿童群体则无明显变化;而在控制组中,高易激惹儿童组与中度易激惹儿童组安全型依恋的比例无显著差异。该研究结果验证了差别易感性的“光明面”,未能成功验证差别易感性的“黑暗面”。所以该研究结果更符合优势敏感性模型。
3 评论与展望
从检验模型的角度来说,上述研究质量不一,其中Blair(2002)和Beach等(2010)的研究较为严格地检验了差别易感性模型。从处理“什么对谁最有效(what works for whom)”问题的角度来说,则大多数研究结果表明具备某些特征的特定群体更易从特定的心理干预中获益。这些特征可分为3类:人格和气质类特征,包括高感知加工敏感性、高消极情绪性和高易激惹性;基因类特征,包括5-HTTLPR短等位基因、DRD4-7重复等位基因、糖皮质醇激素受体基因单核苷酸多态区rs10482672-A等位基因拷贝和CRHR1-TAT单体型0重复基因等;生理类,如对愤怒的低呼吸性窦性心律反应性。而且心理干预和干预效果都是多样化的,说明了易感特征有相对广泛的适用性。但是这些研究数量较少,有些结论还不一致。例如5-HTTLPR短等位基因和DRD4-7重复等位基因得到一些研究的支持(Bakermans-Kranenburg et al.,2008;Beach et al.,2010),却在其他干预研究中并未表现出调节效应(Cicchetti et al.,2011),甚至在有的研究中5-HTTLPR长等位基因携带者有最大的获益(Cicchetti et al.,2015)。
两个模型的发展历史较短,理论还不完备,心理干预研究证据更是处于初期阶段。要得出较为稳定的结论,需要经过大量证据积累和元分析的检验。理论模型的现状和研究结果为重新看待心理干预与当事人提供了一些启发,也为以后的研究提供了一些铺垫。
3.1 现有研究的启发
3.1.1 治疗遗传学的出现
差别易感性模型遗传标记的心理干预研究催生了一个新的研究领域——治疗遗传学(therapygenetics)(Eley,2014)。治疗遗传学,指用基因标记预测心理治疗的效果(Eley et al.,2012)。已有的研究关注的是5-HTTLPR基因区。Eley等(2012)研究了5-HTTLPR基因在认知行为疗法中的调节作用,对359名有焦虑障碍的6~13儿童实施认知行为疗法,结果发现在治疗结束后的6个月,2个5-HTTLPR短等位基因携带者的焦虑症状改善显著优于1或2个长等位基因携带者。与Eley等(2012)的研究结果不一致,Bockting,Mocking,Lok,Koeter和Schene(2012)对180个反复发作的抑郁病人研究则表明,5-HTTLPR基因与抑郁病人对认知行为疗法的反应无关。这个新领域与两个模型一样,面临着许多类似的问题,也提供了一些启示。
3.1.2 理论意义和应用意义
从理论意义上来说,这两个模型对临床与咨询心理学领域最大的启示是:当事人从心理咨询中获益的可能性和程度充满了多样性和差异性。而Bohart和Tallman(2010)可能不太认同当事人获益可能性有差异的说法,他们认为是当事人的自愈能力被激活的程度的差异,导致了当事人从心理干预中获益程度不一。前者强调当事人与心理干预的交互作用,后者强调当事人的主效应。因此在以后的研究中,需要进一步确证心理干预中当事人的主效应以及与干预的交互效应,为两种看法的争论提供事实依据。
在实际应用上,Pluess和Belsky(2013)提出一个大胆的设想,直接干预个体的优势敏感性使其具备高优势敏感性,使得个体能够持续性获益,必将极大提升心理干预的效果。当然,这个设想还有待于对优势敏感性和差别易感性的发生发展机制的进一步研究。
然而,差别易感性模型和优势敏感性模型也传达出这样一种信号:有些个体难以从心理干预中获益。一方面,这有助于对进入到心理干预中的当事人进行效果预测。Hudson等人(2013)的研究也支持这一信号,通过使用包括神经生长因子(Nerve Growth Factor,NGF)基因rs6330的一些因子构建风险指数,表明风险指数高的儿童持续焦虑障碍的可能性几乎是风险指数低的儿童的3倍。另一方面,这似乎悲观地意味着低易感性或无易感性的个体则难以得到改善,而且将面临一定的伦理风险——造成对非易感个体的歧视和污名(Brody,Beach,et al.,2013;Eley,2014;Ellis et al.,2011)。Pluess和Belsky(2013)态度比较乐观,他们认为鉴别出难以获益的群体以后,可以发展出针对该群体的特定干预方法。Frick,Ray,Thornton和Kahn(2014)的综述支持了这一观点,大部分研究中高冷酷无情特质(callous-unemotional traits)的品行障碍儿童治疗效果更差(较符合“优势抵抗”概念),但在一些加强的干预中,他们也同样出现了好的治疗效果。
总的来说,两个模型及其相关心理干预研究不仅提供了对当事人特征的新理解、对心理干预效应的重新评估,也提供了对效果的预测,这些都为发展个人化治疗提供了理论和证据基础(personalized treatment)(Klengel&Binder,2013;Landrø,2014),便于为不同的当事人提供更恰当的服务。然而,要达成这些目标,现有研究中存在的问题不得不引起重视。
3.2 现有研究存在的问题
3.2.1 发现了干预遗失的效率还是心理干预的优先获益者?
一些研究者认为两个模型的心理干预研究解释了为何以往的研究发现心理干预的效应比较小,因为干预对象中有相当一部分不具备易感性的个体,他们很难从心理干预中获益,而易感性的个体的心理干预效应则比较高,于是研究者认为发现了心理干预隐藏的效果(Bakermans-Kranenburg&van IJzendoorn,2015;Eley,2014;Klengel&Binder,2013),似乎提升了对心理干预的信心。然而这些结论都必须建立在这些心理干预是毋庸置疑积极有效的基础上。
那么,如何界定一项心理干预是否是“积极的”呢?这些心理干预都有深厚的理论基础和实证支持,但这种支持一般都来自于对某一特殊群体的干预效果检验,而不是面对全体,因此有效性的确证过程有可能是一个偏向形成的过程,使得这种干预只适用于某种特殊群体,而不适用于其他群体。当然,从来也不存在一种适用于所有个体的干预方法。那么,在研究中就遇到一个难题,能不能将只对部分个体有效的心理干预称为“积极”呢?能不能以这种干预来衡量个体是否具备易获益的特征呢?
实际上,上述研究默认心理干预的积极属性,是权衡的结果。因为发展可塑性的两个模型强调的是个体特征与环境的交互作用,在心理干预领域中,研究需要检验个体特征是否调节了干预与干预效果之间的关系,也需要检验干预对个体特征与干预效果的关系的调节。但在两个模型提出之初,大部分证据来自于对以往大型研究项目的数据分析,一般只包括一种心理干预,因此一次只检验一种调节作用,而且干预是“积极的”比个人特征是“有助于获益的”似乎更有说服力,因此现有研究均是对前一种调节关系的检验。将这两个模型应用到心理干预领域中,研究者若沉浸于对心理干预的信心中,则只能回答“什么样的个体是心理干预的优先获益者”,而不能获知“什么样的个体更易从什么样的心理干预中获益”。在以后的研究中,如何确定心理干预的积极属性,如何完善对两种调节关系的检验,都将是研究者需要考虑的重要问题。
3.2.2 模型的验证问题
虽然模型的验证并不是本文关注重点,但两个模型的区别(见图1),即个体是否具备在消极环境中受损的特质,是确定某种使个体获益的特征是否同时属于致病因子的关键。但目前整体研究趋势与以往的素质-压力模型只关注消极环境一样,只要研究从积极环境中得到显著的结果,研究者就急于得出符合差别易感性的结论,对消极环境的结果的探究明显不够。这导致两个模型在实际的研究中界限模糊。然而,如果将有困扰的当事人置于消极环境中,则可能面临严峻的伦理问题,这也是干预研究一直没有被试间设计的重要原因。
另外一个与模型检验有关的问题是,模型未验证说明了什么?一种解释是说明模型验证失败,当然一个模型的建立或推倒都不能由一个研究决定;另一种解释是模型未验证说明了模型的边界(Bakermans-Kranenburg&van IJzendoorn,2015),即模型的适用范围。显然后一种解释有造成对该模型不可证伪的风险。
3.2.3 两个模型自身存在的问题
目前尚不能确定差别易感性和优势敏感性是连续变量还是离散变量,是跨领域的特征还是特定领域的特征(Belsky&Pluess,2009,2013;Ellis et al.,2011;Pluess&Belsky,2013)。前者关系着个体从积极环境中的获益程度与可能性是“全或无”还是“多或少”的问题。后者则容易造成对研究结果的不同解释。当具备某些优势敏感性或差别易感性标志的个体在积极环境中并未出现获益时,究竟是否定了模型的合理性,还是由于这些特征只与特定的积极环境有交互作用,还是由于环境不够积极,则难以确定。因此在解释两个模型在心理干预领域的发现时,必须考虑这两个问题。
3.2.4 其他问题和局限
现有研究的心理干预以干预个体所处的环境居多。上述声称验证了差别易感性模型的干预研究(Bakermans-Kranenburg et al.,2008;Beach et al.,2010;Blair,2002;Cassidy et al.,2011;Scott&O’Connor,2012),主要聚焦于对父母的教养训练,即干预儿童所生存的环境,而不是直接干预儿童本身。这与发展可塑性研究长期以来集中于教养与环境的交互作用与基因与环境交互作用有关(Manuck&McCaffery 2014)。直接针对目标个体进行的心理干预能否为个体营造一个有影响力的环境呢?少数的一些直接干预研究似乎对此问题作出了肯定的回答。一个新的问题就出现了,从心理干预中获益的易感个体回到其消极且未发生变化的生活环境后,是会重新受损于其消极的生活环境呢,还是足以应对其生活环境?其结果和结果的解释现在都不得而知。
现有研究以基因与干预的交互作用研究最多,而且以候选基因关联研究(candidate gene association study)为主。候选基因关联研究是“根据既有遗传相关信息(基因位置)、生物相关信息(SNP功能类和通路信息)或者实证研究结果来直接选定可能与个体心理或行为表型变异有关的基因,并通过病例对照设计(case-control study)或基于随机人群的关联分析(population-based association analysis)来确定候选基因与该表型是否存在关联的一种研究策略”(引自张文新等,2012)。这与遗传与环境交互作用领域的研究趋势一致,但存在效应小、难复制的问题(Eley,2014;Manuck&McCaffery,2014)。只有少数研究考虑了相对广泛的多基因研究(Musci et al.,2015),大部分研究没有同时考虑多种基因。至于将3种易感标记同时纳入研究范围,则更是一个都没有。
现有心理干预研究样本多为婴幼儿与青少年,成年人样本较少。这可能是因为青少年及婴幼儿的发展可塑性高于成年人,因应心理干预而发生变化的能力更强,干预效果会更为显著。但进入到心理咨询中的有困扰的成人为数不少,有必要检验成人在心理干预这种新的积极环境中获益程度的差异。
除了特定的年龄特征外,上述研究被试群体也具备某些特定的病理性特征或风险特征,如出生体重低、焦虑障碍、行为问题等,因此,在这些特殊被试群体身上得到的研究发现是否能直接推广到一般群体中,还有待商榷。此外,从测量上来说,针对基因或生理标志的研究评定较为客观,但在人格和气质研究中,对气质的评估往往是通过儿童的养育者的报告或者儿童的自我报告,可能会有偏差。而现有研究被试以高加索人种为主,唯一的干预研究元分析发现种族可能是一个重要的调节变量(van IJzendoorn&Bakermans-Kranenburg,2015),但目前还缺乏对其他种族的研究。
3.3 研究展望
3.3.1 探索易感特征对心理干预效果差异的解释
虽然现有证据表明,具备某些特征的个体比其他个体更易从积极环境中获益,但并没有研究表明心理干预的获益差异有多少可归于差别易感性和优势敏感性的这些特征,即这些易感特征对心理干预效果的解释量有多少,尚是未知之数。只有在证明这个解释量较大的前提下,这个领域的研究才有意义。在证明两个模型成立及发现更多易感特征的同时,以后的研究应着手回答这个问题,为该领域的研究价值提供依据。
3.3.2 探索差别易感性和优势敏感性的机制
若想将差别易感性和优势敏感性的发现应用到心理干预的实践中,必须对其作用机制有深刻的理解。现有研究以基因方面的研究居多,而基因方面的研究以DRD4和5-HTTLPR基因区居多,提示多巴胺、5-羟色胺等神经递质可能在个体与环境的交互过程中起作用。此外,一些研究者指出治疗过程中基因表达的变化和DNA的甲基化以及特定的信息加工或大脑功能是未来机制的研究方向(Bakermans-Kranenburg&van IJzendoorn,2015;Eley,2014;Klengel&Binder,2013)。
3.3.3 扩展易感特征范围、环境范围及被试范围
现有易感特征的研究集中于基因方面,基因自有其客观、稳定的优势,而基因较难应用于临床实践中,有必要进一步增加生理、气质和人格方面的易感特征的研究。为了更严格地检验与区分两个模型,首先要做的是扩展研究的环境范围。根据Pluess和Belsky(2013)的建议,以后的研究应需要对个体在相对广泛的环境中的表现型进行检验,包括从消极环境到无消极环境,再从无积极环境到积极环境。Manuck和McCaffery(2014)也建议先建立起各种基因型、基因组合的反应常模(reaction norm)。反应常模是指同一基因型的个体在不同环境条件下产生的表型变异的范围(Manuck,2010)。而且个体气质与生理反应特征也应有其反应常模。在建立完好的反应常模的基础之上,两个模型则能被证实或证伪。就心理干预这一特殊的积极环境来说,首先需要增加直接心理干预的研究,来说明两个模型是否同样适用于未改变个体生存环境的直接心理干预。其次,需要在多种心理干预中检验各种潜在的易感标记是否具备易感性,在同一个研究中检验多种心理干预则有助于检验心理干预对个体特征与干预效果之间关系的调节作用。此外,为了扩展两个模型的解释范围,有必要在不同障碍、不同种族、不同年龄的被试群体中检验两个模型(Eley,2014;van IJzendoorn&Bakermans-Kranenburg,2015)。
3.3.4 多种易感特征的关系及综合分析
由于候选基因关联研究的局限,未来研究将朝向全基因组关联研究(genome-wide association study,GWAS),考察更广阔的基因组合。这是不同基因的综合分析,而人格、气质类和生理类也需要综合分析。同样属于易感标记,这些不同类别的易感特征间是否有关联呢?这是一个需要回答的问题。而且,已有研究表明依恋风格、应对风格、心理感受性(psychological mindedness)、进入情绪体验的能力以及完美主义等人格变量与个体从心理治疗中获益的程度有关(Bohart&Wade,2013),可探讨生理类和基因类易感特征与这些人格变量的关系,促进对这些人格变量更深层次机制的认识。在此基础之上,可同时考虑多种类别的易感标记的作用(Bakermans-Kranenburg&van IJzendoorn,2015)。
致谢:感谢孙启武老师对本文提出的宝贵意见;感谢应梦婷、朱文臻博士对英文摘要的修改和润色;感谢林秀彬博士为本文最后把关;感谢周忠英、黄俊同学在本文书写上所提供的宝贵建议。
曹丛,王美萍,张文新,纪林芹,陈亮,陈欣银.(2014).COMT基因rs6267多态性与母亲教养行为对青少年身体攻击和关系攻击的交互作用.心理学报,46(10),1486–1497.
王美萍,张文新,陈欣银.(2015).5-HTR1A基因rs6295多态性与父母教养行为对青少年早期抑郁的交互作用:不同易感性模型的验证.心理学报,47(5),600–610.
张璠,陈杰,李新影.(2015).BDNF Val66Met基因多态性和温暖的教养方式对青少年抑郁的影响.中国临床心理学杂志,23(1),52–55.
张卫,甄霜菊.(2013).发展可塑性的个体差异——基因与环境的交互作用.华南师范大学学报:社会科学版,(5),106–111.
张文新,王美萍,曹丛.(2012).发展行为遗传学简介.心理科学进展,20(9),1329–1336.
Albert,D.,Belsky,D.W.,Crowley,D.M.,Bates,J.E.,Pettit,G.S.,Lansford,J.E.,...Dodge,K.A.(2015).Developmental mediation of genetic variation in response to the Fast Track prevention program.Development andPsychopathology,27(1),81–95.
Anzman-Frasca,S.,Stifter,C.A.,Paul,I.M.,&Birch,L.L.(2014).Negative temperament as a moderator of intervention effects in infancy:Testing a differential susceptibility model.Prevention Science,15(5),643–653.
Bakermans-Kranenburg,M.J.,&van IJzendoorn,M.H.(2006).Gene-environment interaction of the dopamine D4 receptor(DRD4)and observed maternal insensitivity predicting externalizing behavior in preschoolers.Developmental Psychobiology,48(5),406–409.
Bakermans-Kranenburg,M.J.,&van IJzendoorn,M.H.(2011).Differential susceptibility to rearing environment depending on dopamine-related genes:New evidence and a meta-analysis.Development and Psychopathology,23(1),39–52.
Bakermans-Kranenburg,M.J.,&van IJzendoorn,M.H.(2015).The hidden efficacy of interventions:Gene×environment experiments from a differential susceptibility perspective.Annual Review of Psychology,66,381–409.
Bakermans-Kranenburg,M.J.,van IJzendoorn,M.H.,Pijlman,F.T.A.,Mesman,J.,& Juffer,F.(2008).Experimental evidence for differential susceptibility:Dopamine D4 receptor polymorphism (DRD4 VNTR)moderates intervention effects on toddlers' externalizing behavior in a randomized controlled trial.Developmental Psychology,44(1),293–300.
Bateson,P.,Barker,D.,Clutton-Brock,T.,Deb,D.,D'Udine,B.,Foley,R.A.,...Lahr,M.M.(2004).Developmental plasticity and human health.Nature,430(6998),419–421.
Beach,S.R.H.,Brody,G.H.,Lei,M.-K.,&Philibert,R.A.(2010).Differential susceptibility to parenting among African American youths:Testing the DRD4 hypothesis.Journal of Family Psychology,24(5),513–521.
Belsky,J.(2007).Experience in childhood and the development of reproductive strategies.Acta Psychologica Sinica,39(3),454–468.
Belsky,J.,Bakermans-Kranenburg,M.J.,&van IJzendoorn,M.H.(2007).For better and for worse:Differential susceptibility to environmental influences.Current Directions in Psychological Science,16(6),300–304.
Belsky,J.,&Pluess,M.(2009).Beyond diathesis stress:Differential susceptibility to environmental influences.Psychological Bulletin,135(6),885–908.
Belsky,J.,&Pluess,M.(2013).Beyond risk,resilience,and dysregulation:Phenotypic plasticity and human development.Development and Psychopathology,25(4 Pt 2),1243–1261.
Belsky,J.,&van IJzendoorn,M.H.(2015).What works for whom? Genetic moderation of intervention efficacy.Development and Psychopathology,27(1),1–6.
Blair,C.(2002).Early intervention for low birth weight,preterm infants:The role of negative emotionality in the specification of effects.Development and Psychopathology,14(2),311–332.
Bockting,C.L.H.,Mocking,R.J.,Lok,A.,Koeter,M.W.J.,&Schene,A.H.(2012).Therapygenetics:The 5HTTLPR as a biomarker for response to psychological therapy?Molecular Psychiatry,18(7),744–745.
Bohart,A.C.,&Tallman,K.(2010).Clients:The neglected common factor in psychotherapy.In B.L.Duncan,S.D.Miller,B.E.Wampold,&M.A.Hubble(Eds.),The heart and soul of change:Delivering what works in therapy(2th ed.,pp.83–111).Washington,DC,US:American Psychological Association.
Bohart,A.C.,& Wade,A.G.(2013).Theclientin psychotherapy.In M.J.Lambert(Ed.),Bergin and Garfield’s handbook of psychotherapy and behavior change(6th ed.,pp.219–257).Hoboken,New Jersey:John Wiley&Sons Inc.
Brett,Z.H.,Sheridan,M.,Humphreys,K.,Smyke,A.,Gleason,M.M.,Fox,N.,...Drury,S.(2015).A neurogenetics approach to defining differential susceptibility to institutional care.International Journal of Behavioral Development,39,150–160.
Brody,G.H.,Beach,S.R.H.,Hill,K.G.,Howe,G.W.,Prado,G.,&Fullerton,S.M.(2013).Using genetically informed,randomized prevention trials to test etiological hypotheses about child and adolescent drug use and psychopathology.American Journal of Public Health,103(S1),S19–S24.
Brody,G.H.,Chen,Y.F.,&Beach,S.R.H.(2013).Differential susceptibility to prevention:GABAergic,dopaminergic,and multilocus effects.Journal of Child Psychology and Psychiatry,54(8),863–871.
Brody,G.H.,Yu,T.Y.,&Beach,S.R.H.(2015).A differential susceptibility analysis reveals the“who and how”about adolescents' responses to preventive interventions:Tests of first-and second-generation gene×intervention hypotheses.Development and Psychopathology,27(1),37–49.
Carlson,M.D.,Mendle,J.,&Harden,K.P.(2014).Early adverse environments and genetic influences on age at first sex:Evidence for gene×environment interaction.Developmental Psychology,50(5),1532–1542.
Cassidy,J.,Woodhouse,S.S.,Sherman,L.J.,Stupica,B.,&Lejuez,C.W.(2011).Enhancing infant attachment security:An examination of treatment efficacy and differential susceptibility.Development and Psychopathology,23(1),131–148.
Cicchetti,D.,Rogosch,F.A.,&Toth,S.L.(2011).The effects of child maltreatment and polymorphisms of the serotonin transporter and dopamine D4 receptor genes on infant attachment and intervention efficacy.Development and Psychopathology,23(2),357–372.
Cicchetti,D.,Toth,S.L.,&Handley,E.D.(2015).Genetic moderation of interpersonal psychotherapy efficacy for low-income mothers with major depressive disorder:Implications for differential susceptibility.Development and Psychopathology,27(1),19–35.
Cleveland,H.H.,Schlomer,G.L.,Vandenbergh,D.J.,Feinberg,M.,Greenberg,M.,Spoth,R.,...Hair,K.L.(2015).The conditioning of intervention effects on early adolescent alcohol use by maternal involvement and dopamine receptor D4(DRD4)and serotonin transporter linked polymorphic region(5-HTTLPR)genetic variants.Development and Psychopathology,27(1),51–67.
Eley,T.C.(2014).The future of therapygenetics:Where will studies predicting psychological treatment response from genomic markers lead?Depression and Anxiety,31(8),617–620.
Eley,T.C.,Hudson,J.L.,Creswell,C.,Tropeano,M.,Lester,K.J.,Cooper,P.,...Rapee,R.M.(2012).Therapygenetics:The 5HTTLPR and response to psychological therapy.Molecular Psychiatry,17(3),236–237.
Ellis,B.J.,Boyce,W.T.,Belsky,J.,Bakermans-Kranenburg,M.J.,& van IJzendoorn,M.H.(2011).Differential susceptibility to the environment:An evolutionary–neurodevelopmental theory.Development and Psychopathology,23(1),7–28.
Frick,P.J.,Ray,J.V.,Thornton,L.C.,&Kahn,R.E.(2014).Can callous-unemotional traits enhance the understanding,diagnosis,and treatment of serious conduct problems in children and adolescents? A comprehensive review.Psychological Bulletin,140(1),1–57.
Gatzke-Kopp,L.M.,Greenberg,M.,&Bierman,K.(2015).Children's parasympathetic reactivity to specific emotions moderates response to intervention for early-onset aggression.Journal of Clinical Child&Adolescent Psychology,44(2),291–304.
Hudson,J.L.,Lester,K.J.,Lewis,C.M.,Tropeano,M.,Creswell,C.,Collier,D.A.,...Rapee,R.M.(2013).Predicting outcomes following cognitive behaviour therapy in child anxiety disorders:The influence of genetic,demographic and clinical information.Journal of Child Psychology and Psychiatry,54(10),1086–1094.
Klengel,T.,&Binder,E.B.(2013).Gene×environment interactions in the prediction of response to antidepressant treatment.International Journal of Neuropsychopharmacology,16(3),701–711.
Landrø,N.I.(2014).Towards personalized treatment of depression:A candidate gene approach.Scandinavian Journal of Psychology,55(3),219–224.
Manuck,S.B.(2010).The reaction norm in gene×environment interaction.Molecular Psychiatry,15(9),881–882.
Manuck,S.B.,&McCaffery,J.M.(2014).Gene-environment interaction.Annual Review of Psychology,65,41–70.
Monroe,S.M.,&Simons,A.D.(1991).Diathesis-stress theories in the context of life stress research:Implications for the depressive disorders.Psychological Bulletin,110(3),406–425.
Moczek,A.P.,Sultan,S.,Foster,S.,Ledón-Rettig,C.,Dworkin,I.,Nijhout,H.F.,...Pfennig,D.W.(2011).The role of developmental plasticity in evolutionary innovation.Proceedings of the Royal Society B:Biological Sciences,278,2705–2713.
Musci,R.J.,Masyn,K.E.,Uhl,G.,Maher,B.,Kellam,S.G.,&Ialongo,N.S.(2015).Polygenic Score×Intervention Moderation:An application of discrete-time survival analysis to modeling the timing of first tobacco use among urban youth.Development and Psychopathology,27(1),111–122.
Nederhof,E.,Belsky,J.,Ormel,J.,&Oldehinkel,A.J.(2012).Effects of divorce on Dutch boys' and girls' externalizing behavior in Gene×Environment perspective:Diathesis stress or differential susceptibility in the Dutch Tracking Adolescents' Individual Lives Survey study?Development and Psychopathology,24(3),929–939.
Pluess,M.,& Belsky,J.(2013).Vantage sensitivity:Individual differences in response to positive experiences.Psychological Bulletin,139(4),901–916.
Pluess,M.,& Boniwell,I.(2015).Sensory-Processing Sensitivity predicts treatment response to a school-based depression prevention program:Evidence of vantage sensitivity.Personality and Individual Differences,82,40–45.
Schlomer,G.L.,Cleveland,H.H.,Vandenbergh,D.J.,Feinberg,M.E.,Neiderhiser,J.M.,Greenberg,M.T.,...Redmond,C.(2015).Developmental differences in early adolescent aggression:A gene×environment×intervention analysis.Journal of Youth and Adolescence,44(3),581–597.
Scott,S.,&O’Connor,T.G.(2012).An experimental test of differential susceptibility to parenting among emotionallydysregulated children in a randomized controlled trial for oppositional behavior.Journal of Child Psychology and Psychiatry,53(11),1184–1193.
Sweitzer,M.M.,Halder,I.,Flory,J.D.,Craig,A.E.,Gianaros,P.J.,Ferrell,R.E.,&Manuck,S.B.(2013).Polymorphic variation in the dopamine D4 receptor predicts delay discounting as a function of childhood socioeconomic status:Evidence for differential susceptibility.Social Cognitive and Affective Neuroscience,8(5),499–508.
van den Hoofdakker,B.J.,Nauta,M.H.,Dijck-Brouwer,D.A.J.,van der Veen-Mulders,L.,Sytema,S.,Emmelkamp,P.M.G.,...Hoekstra,P.J.(2012).Dopamine transporter gene moderates response to behavioral parent training in children withADHD:A pilotstudy.Developmental Psychology,48(2),567–574.
van IJzendoorn,M.H.,&Bakermans-Kranenburg,M.J.(2012).Differential susceptibility experiments:Going beyond correlational evidence:Comment on beyond mental health,differential susceptibility articles.Developmental Psychology,48(3),769–774.
van IJzendoorn,M.H.,&Bakermans-Kranenburg,M.J.(2015).Genetic differential susceptibility on trial:Metaanalytic support from randomized controlled experiments.Development and Psychopathology,27(1),151–162.
