稀有糖D-阿洛糖性质及生物法生产研究进展*
2015-11-20冯再平巩慧玲袁慧君沐万孟江波
冯再平,巩慧玲,袁慧君,沐万孟,江波
1(兰州理工大学生命科学与工程学院,甘肃兰州,730050)
2(江南大学食品科学与技术国家重点实验室,江苏无锡,214122)
稀有糖(rare sugar)是一类重要的碳水化合物,在膳食、保健、医药等领域中发挥着非常重要的功能。根据国际糖协会(ISRS)定义,稀有糖是“在自然界存在但含量极少的一类单糖及其衍生物”,一般具有低热量、低吸收等特点,并且具有多种生理功能,在膳食、保健、医药等领域发挥着重要作用。D-塔格糖(D-tagatose)具有热量低、机体不能代谢或者代谢很少、抗龋齿、降血糖、改善肠道菌群等优点[1],其甜味刺激和果糖类似,较蔗糖快,口感、甜度和蔗糖较为类似,是一种优秀的低能量甜味剂;而D-阿洛酮糖具有保护胰腺β岛细胞、改善胰岛素敏感度和葡萄糖耐受性、减少腹部脂肪积累、清除活性氧自由基等作用,是一种很有前途的无能量的功能性甜味剂[2];L-系列糖缺乏D-型糖类的风味,有许多特殊的生理功能,可用作化工、医药产品的中间体,如 L-核糖及衍生物[3]。其中,D-阿洛糖由于具有独特的生理功能,成为近年来稀有糖研究的热点。
1 D-阿洛糖概述
1.1 D-阿洛糖的结构、理化性质及安全性
D-阿洛糖,是一种具广泛生理功能的稀有顺式己醛糖,是稀有糖D-阿洛酮糖的醛糖异构体、D-葡萄糖C-3位置上的差向异构体,存在于一些天然植物的提取物及细菌代谢物中[4]。
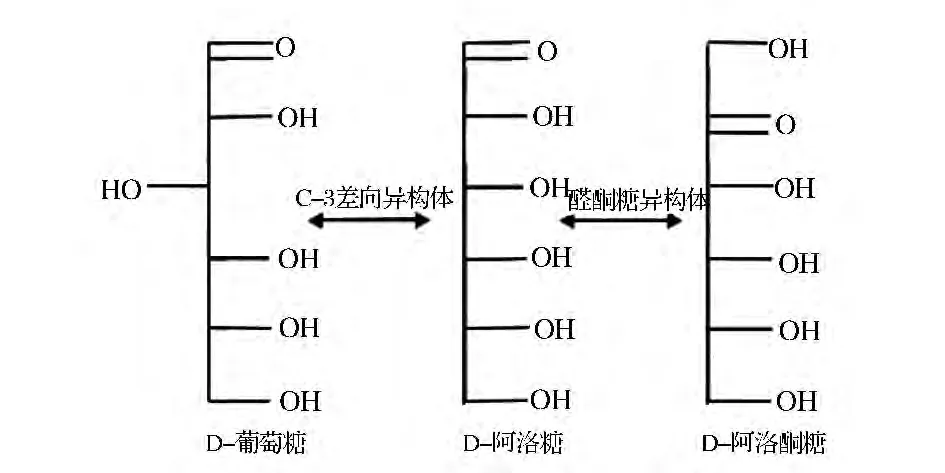
图1 D-葡萄糖、D-阿洛糖和D-阿洛酮糖的化学结构式Fig.1 Chemical structures of D-glucose,D-allose and D-psicose
D-阿洛糖是无味的白色晶体,分子式为C6H12O6,分子质量为180.16,熔点为128℃,旋光度为[α]20+14°(c=1.00%,H2O),易溶于水,不溶于乙醇。D-阿洛糖属于五羟基醛,可以以直链、环状形式存在,环状形式又分为呋喃型(D-阿洛糖的醛基与C-4上的羟基缩合形成)、吡喃型(D-阿洛糖的醛基与C-5上的羟基缩合形成)两种形式。由于D-阿洛糖分子中既有醛基,又有羟基,能够作用形成半缩醛从而形成一对非对映旋光异构体(α-与β-异头体)。天然化合物中,D-阿洛糖多以β-吡喃型的形式存在,在D-阿洛糖的水溶液中,含β-D-吡喃阿洛糖(77.5%),α-D-吡喃阿洛糖(14%),β-D-呋喃阿洛糖(5%),和 α-D-呋喃阿洛糖(3.5%)[5]。
D-阿洛糖的急性、亚慢性毒性试验表明D-阿洛糖是无毒的;大鼠的经口LD50为20.5 g/kg,在亚慢性实验中,对照与试验组的血清化学和血液学检验结果绝大多数没有明显差异,而营养学方面的特性与D-葡萄糖、D-果糖等单糖是不同的[6]。

表1 D-阿洛糖的理化性质Table 1 Physical and chemical properties of D-allose
1.2 D-阿洛糖在大肠杆菌中的代谢途径
D-阿洛糖可以通过细胞膜上的转运系统进入细胞质,参与分解代谢的糖酵解途径(图2)。3种蛋白(AlsA,AlsB,and AlsC)参与D-阿洛糖运输。D-阿洛糖先与D-阿洛糖-结合蛋白(AlsB)结合,经ABC转运体(包括ABC转运蛋白AlsA和转运元件AlsC)转运进入细胞质。在细胞质中,EMP途径的D-阿洛糖激酶(AlsK),D-阿洛糖-6-磷酸异构酶(AlsI),和D-阿洛酮糖-6-磷酸3-差向异构酶(AlsE)经过顺序酶反应,经由两个中间产物D-阿洛糖6-磷酸和D-阿洛酮糖6-磷酸,实现将 D-阿洛糖转化成 D-果糖-6-磷酸[7]。
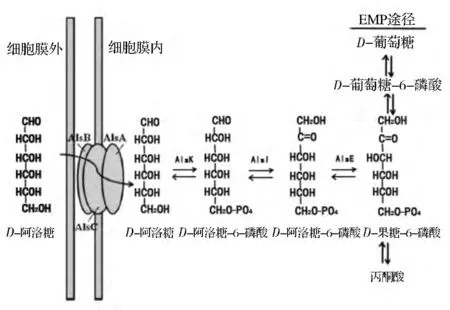
图2 D-阿洛糖膜转运和细胞内代谢示意图[8]Fig.2 Schematic illustration for the proposed membrane transport and intracellular metablolism of D-allose
1.3 D-阿洛糖功能性质
在稀有糖中,D-阿洛糖由于具有广泛的生理功能,而成为近年来的研究热点。其生理功能主要包括:
1.3.1 抑制致癌作用及抑制各种癌细胞系的增殖[8]
D-阿洛糖的众多生理功能中,最引人注目的就是其显著的抑癌作用。D-阿洛糖可抑制氧化应激条件下的致癌作用发生,并且能够诱导肿瘤抑制基因——硫氧还蛋白相互作用蛋白(thioredoxin interacting protein,TXNIP)表达,以剂量依赖形式抑制前列腺、头颈、肝、卵巢、子宫颈及皮肤等不同来源的癌细胞系以及白血病。
1.3.2 癌症放、化疗协同增效作用
2011年,Hoshikawa等[9]报道了 D-阿洛糖对头颈癌细胞HSC-3的放射增敏作用,发现合并D-阿洛糖和放射治疗比两者单独使用的效果要好。在37%的存活水平上,10和25 mmol/L D-阿洛糖处理组的辐射增强率分别达到1.61和2.11。并且两者的联合使用还可以减少3D培养实验中癌细胞增殖。单独放射处理时不能增加TXNIP的mRNA表达水平,而D-阿洛糖合并放射处理可以显著地提高TXNIP表达。两者合并使用时,可以显著地诱导细胞内活性氧自由基的产生和细胞凋亡的发生。
通常的抗肿瘤药物5-氟尿嘧啶,虽然可以诱导TXNIP表达,抑制胸苷酸合成酶合成而阻碍细胞周期,但具有非特异性细胞毒性,可使患者表现出多种副作用。Yamaguchi等对人肝细胞癌细胞HuH-7的实验表明,D-阿洛糖与5-氟尿嘧啶有协同增效作用,而D-阿洛糖对普通细胞没有已知的副作用,因此D-阿洛糖与5-氟尿嘧啶合并使用可能成为癌症治疗的新方案[10]。这些报道都说明 D-阿洛糖作为癌症治疗药物的有效性和可行性。
1.3.3 抑制高钠诱导型高血压[11]
Kimura等研究了2种高血压动物模型——盐敏感型高血压DS大鼠和自发性高血压大鼠(摄入D-阿洛糖后对高血压发生过程及氧化状态的影响),结果表明,DS大鼠喂食高盐(4%)饲料4周以后,血压明显升高;而在喂高盐饲料同时以2 g/(kg·d)的剂量给予D-阿洛糖时,可以抑制血压的升高,同时伴有减少主动脉过氧化物的生成。而在自发性高血压模型动物中,D-阿洛糖对高血压发生早期的血压或大动脉过氧化物的生成没有明显影响。这个实验说明D-阿洛糖可以应用于抑制高钠诱导型高血压。
1.3.4 抗氧化性及相关作用
2011年,为了阐明D-阿洛糖的抗氧化特性,Ishihara等[12]研究了D-阿洛糖的活性氧自由基清除能力和在线粒体活性氧自由基产生过程中所起的作用。研究结果显示,D-阿洛糖不能清除过氧化氢和超氧阴离子,其清除羟基自由基的能力与D-葡萄糖相当;而在线粒体中,D-阿洛糖可以减弱由鱼藤酮诱导的D-葡萄糖-依赖性活性氧自由基的产生(说明D-阿洛糖在线粒体内抑制活性氧自由基产生,可能与其在细胞水平和D-葡萄糖竞争相关)。Sun等研究表明,D-阿洛糖和α-乳白蛋白之间的美拉德反应产物也具有抗氧化能力[13]。
D-阿洛糖的抗氧化性使其在存在氧化应激的急性局部缺血/再灌注损伤、器官移植手术等相关临床应用中,起到相应的抗炎、保护作用[14]。
1.3.5 免疫抑制、冷冻保护等其他功能
D-阿洛糖的其他功能的报道还包括:Sui等[15]发现,哺乳类动物细胞系包括OVCAR-3、HeLa、HaCaT、HDF、NIH3T3等在-80℃冷冻时,在培养基中添加不同浓度的D-阿洛糖时细胞存活率提高,说明D-阿洛糖可以用作冷冻保护剂;2012年,Harada等[16]发现,D-阿洛糖可以加强甲硝唑对寄生虫毛滴虫的效果,从而减少甲硝唑用量,避免毛滴虫对药物产生抗性;Tanaka等[17]报道,D-阿洛糖与内吞作用以及树突细胞激励T细胞相关,可能起到免疫抑制作用。
1.4 D-阿洛糖的合成策略
由于D-阿洛糖在自然界存在稀少,要开展D-阿洛糖的营养特性、生理功能的深入研究,以及推广D-阿洛糖在功能性食品领域、医疗卫生领域的应用,靠从自然界资源中提取该稀有糖是不现实的,需要实现D-阿洛糖的大规模生产。D-阿洛糖的化学合成法种类繁多,而生物合成法条件温和、前景诱人。下面具体说明D-阿洛糖获得的各种可能途径。
1.4.1 化学合成法
化学合成方法通过各种羟基的选择性保护及水解方法的应用,经氧化还原途径实现各种糖或糖衍生物原料构型改变来合成阿洛糖。如天津大学的张卫红[18]以 α-D-葡萄糖为原料,经氧化还原反应,改变C-3 位羟基的构型,合成 1,2:5,6-氧-二异丙叉基-α-D-呋喃阿洛糖;再经完全水解得到D-阿洛糖。可以看出,化学合成法涉及异丙叉化、氧化、还原、水解等多步反应,步骤繁琐,过程复杂,副产物多。
1.4.2 立体选择性合成法
立体选择性合成法(手性合成法),是利用手性非糖类物质做底物或是潜手性环境,进行立体选择性和立体专一性反应。Mheta等[19]以环辛烷为原料反应得到了(DL)-β-阿洛糖。该方法同样反应步骤多,操作复杂,其立体选择性对由非糖化合物合成稀有糖类的研究具有理论意义。
1.4.3 催化立体异构化法
在合成糖的方法中,催化剂作用下的立体异构化法是较为成熟实用的糖合成方法。其原理是:以钼酸等为催化剂,糖类化合物发生C-2或C-3位的立体异构化反应,使单糖分子羟基构型发生变化。一项D-阿洛糖制备的美国专利[20]就是依据此原理,在酸性条件下以D-葡萄糖为原料,经钼酸盐催化,发生C-3立体异构化反应,从而得到含量7%的产物D-阿洛糖。然后,产物混合液经浓缩、脱色、蒸汽化,流经以离子交换树脂为吸附剂的分离区域,D-阿洛糖被吸附住而得到分离。催化立体异构化法直接简便,解决转化率与收率不高、能耗大、分离方法复杂等问题后,其市场潜力是显而易见的。
1.4.4 生物合成法[8]
如图3所示,目前D-阿洛糖的生物合成需要两步生物转化反应实现。第一步反应以D-果糖为原料,在D-塔格糖3-差向异构酶(D-tagatose 3-epimerase,DTE)家族成员的作用下,C3位发生差向异构化,获得D-阿洛酮糖。该反应的催化机制、酶分子结构、生物转化等都有较多报道。在此基础上可大量制备D-阿洛酮糖,用作附加值更高的D-阿洛糖制备的原料。第二步反应以D-阿洛酮糖为原料,在酮醛糖异构酶的作用下,转化为对应的己醛糖D-阿洛糖。

图3 D-阿洛糖的两步法生物合成Fig.3 Two-step biosynthesis of D-allose
2 生物法产D-阿洛糖研究进展
2.1 L-鼠李糖异构酶
国际上最早开始研究报道生物催化与生物转化法生产D-阿洛糖的是日本香川大学稀有糖研究中心Ken Izumori研究组。1997年,该研究组在研究Pseudomonas stutzeri的L-鼠李糖异构酶(L-Rhamnose Isomerase,L-RhI)时发现该酶具有较宽的底物谱,除了能够可逆催化L-鼠李糖与L-鼠李酮糖之间的醛酮异构反应外,还可以催化包括D-阿洛糖与D-阿洛酮糖在内的多对五元、六元酮醛糖之间的异构化反应[21]。可以说对P.stutzeri L-鼠李糖异构酶的研究开启了生物转化、生物催化法生产稀有糖D-阿洛糖的序幕,但是P.stutzeri L-鼠李糖异构酶不一定就是最适合产D-阿洛糖的生物催化剂来源,用P.stutzeri L-鼠李糖异构酶催化D-阿洛糖的转化反应存在一些问题。首先,该酶催化D-阿洛酮糖和D-阿洛糖之间的异构化反应时,反应平衡体系中并不仅仅只有这两种糖,还存在第三种糖——D-阿卓糖,即该酶催化的是D-阿洛糖、D-阿卓糖和D-阿洛酮糖三者之间的酮醛异构化反应,反应达到平衡以后,体系中三者所占比例分别为25%、8%和67%,这就给产物的分离增加了难度;其次,该酶的耐热性也不是很理想,50℃时的半衰期仅为0.2 h,不能满足工业生产需要;第三,该酶催化反应最适pH值为9.0,而在偏碱性环境下高温反应时,单糖易发生各种副反应导致颜色加深,影响产物质量;最后,该酶需在Mn2+存在下酶活最高,而添加金属离子会增加反应成本及增加产物纯化的工作量。
继发现P.stutzeri L-鼠李糖异构酶产D-阿洛糖的活性之后,寻找产D-阿洛糖酶的新微生物来源,研究酶结构和性质的工作大量开展起来。来自Bacillus pallidus Y25的大肠杆菌重组L-鼠李糖异构酶,在pH 7.0,65℃时有最大酶活,转化D-阿洛酮糖为D-阿洛糖时,平衡比例为65∶35,60℃时半衰期为1 h,没有副产物产生[22]。 重 组 Thermoanaerobacterium saccharolyticum NTOU1 L-鼠李糖异构酶最适pH为7.0,最适温度为75℃,70℃处理2 h后仍有90%酶活,没有副产物生成,平衡比例为 71∶29[23]。重组 Caldicellulosiruptor saccharolyticus ATCC 43494 L-鼠李糖异构酶,非常耐热,其反应最适温度为90℃,这样的温度显然在工业化应用时会增加能源的消耗,最适pH为7.0,80℃处理6 h,酶活仅下降10%,平衡比例为67∶33,没有副产物生成[24]。这些酶在 pH值、耐热性方面比P.stutzeri L-鼠李糖异构酶有了改善,而作用时都需要二价金属离子存在保证最大酶活。
2.2 核糖-5-磷酸异构酶
另一类能够催化D-阿洛酮糖异构化生成D-阿洛糖的酶来自核糖/半乳糖异构酶家族(RpiB/LacA/LacB,蛋白家族IPR003500)。这些酶包括核糖-5-磷酸异构酶 B(ribose-5-phosphate isomerase,RPI)、半乳糖-6-磷酸异构酶 (galactose-6-phosphate isomerase,GPI)等,都属于磷酸糖异构酶,催化异构化反应时一般不需添加金属离子。由于糖磷酸异构酶较宽泛的底物特异性,不仅能够催化含糖磷酸的酮醛糖异构化反应,还可以催化与磷酸糖具有相同骨架构象的单糖的酮醛糖异构化反应,从而可以应用于稀有糖的生物转化。
对核糖/半乳糖异构酶家族产D-阿洛糖酶活性的研究比L-鼠李糖异构酶晚了10年。2007年,韩国建国大学的 Deok-Kun Oh研究组发现,Clostridium thermocellum来源的核糖-5-磷酸异构酶 B[25]可以催化C2、C3和C4羟基取向为同一个方向的酮醛糖之间的可逆异构化反应。用该酶催化D-阿洛酮糖和D-阿洛糖之间可逆的酮醛异构化反应,不需要添加金属离子,不生成D-阿卓糖副产物,最适反应温度为65℃,50℃时的半衰期是P.stutzeri L-鼠李糖异构酶的480倍,最适pH与L-鼠李糖异构酶的9.0相比更接近中性,为7.5。由于没有生成D-阿卓糖副产物,反应达到平衡后,33%的D-阿洛酮糖转化成了D-阿洛糖,转化率比P.stutzeri L-鼠李糖异构酶要高了8%[26]。虽然C.thermocellum核糖-5-磷酸异构酶作用于D-阿洛糖的催化效率kcat/Km为1.5 mmol/(L·min),较P.stutzeri L-鼠李糖异构酶的7.1 mmol/(L·min)要低,但是C.thermocellum核糖-5-磷酸异构酶的酶学性质显然更适应与工业化生产的需要。且经过定点突变C.thermocellum Arg132Glu的催化效率提高到了2.2 mmol/(L·min),减少了与P.stutzeri L-鼠李糖异构酶酶活差距的同时,耐热性更佳,65℃下酶的半衰期是野生型C.thermocellum核糖-5-磷酸异构酶半衰期的1.74倍[27]。不同微生物来源的产D-阿洛糖酶的性质比较见表2。

表2 不同微生物来源的产D-阿洛糖酶的性质Table 2 Biochemical properties of D-allose production enzymes from different microorganism origins

续表2
核糖/半乳糖异构酶家族蛋白作用的最适底物是磷酸糖,除了C.thermocellum核糖-5-磷酸异构酶,其他有产D-阿洛糖活性的核糖/半乳糖异构酶家族成员的报道并不多,且催化效率都不太高,以D-阿洛糖为底物转化为D-阿洛酮糖时,Clostridium difficile核糖-5-磷酸异构酶[29]为 0.03 mmol/(L·min),Thermotoga maritime核糖-5-磷酸异构酶[29]为0.06 mmol/(L·min),Lactococcus lactis半乳糖-6-磷酸异构酶[30]为 0.17 mmol/(L·min)。
2.3 生物法产D-阿洛糖
生物法合成D-阿洛糖,理论上最经济直接的线路就是利用差向异构酶作用于底物D-葡萄糖,直接获得D-阿洛糖。可惜的是,目前为止,自然界还没有发现能实现此反应的酶类。因此,利用生物法合成D-阿洛糖较为实际的路线是经过两步酶或生物转化反应,先将D-果糖差向异构化为D-阿洛酮糖,再将D-阿洛酮糖酮醛异构化为D-阿洛糖,目前报道的生物法合成D-阿洛糖多是按照此路线,以D-阿洛酮糖为底物实现的(表3)。在产物的分离纯化方面,利用钙型阳离子树脂分离 D-阿洛酮糖与 D-果糖混合物[31]以及 D-阿洛酮糖与 D-阿洛糖的混合物[32]的研究已有报道。

表3 应用微生物酶以D-阿洛酮糖为底物生产D-阿洛糖Table 3 D-Allose production from D-psicose by microbial enzymes
而韩文佳等报道[33],利用 D-阿洛酮糖3-差向异构酶 (D-psicose 3-epimerase,DPE)和L-鼠李糖异构酶双酶偶联反应,以2%的D-果糖为底物,经10 h反应达到平衡,得到5.12和2.04 g/L的D-阿洛酮糖和D-阿洛糖,较2种酶单个反应的转化率要高。虽然由于起始酶的催化效率不高,导致产物浓度较低,仍不失为利用廉价原料获得2种稀有糖的有益尝试。
3 小结
生物法产D-阿洛糖的大规模应用,需要解决两个问题。首先是适合工业化生产的异构酶的来源问题,目前还没有作用范围偏酸性的酶催化D-阿洛酮糖转化为D-阿洛糖的报道。另外一方面是底物成本的问题,直接以D-阿洛酮糖为底物的生产成本过高。因为生物法产D-阿洛糖的底物——D-阿洛酮糖本身是一种具有降血糖、抑制肥胖细胞生成、低热量等功能的稀有糖,被FDA认定为“GRAS”物质,是一种非常有前途的新型甜味剂,本身也是价格不菲。如果能够找到直接将D-葡萄糖异构化为D-阿洛糖的差向异构酶,有望实现从D-葡萄糖一步法获得D-阿洛糖;而目前没有相应酶发现的前提下,利用D-阿洛酮糖-3差向异构酶和酮醛糖异构酶的偶联作用,以D-果糖为底物反应获得2种稀有糖(D-阿洛酮糖和D-阿洛糖)是可行的降低底物成本的方案。
[1]Oh D K.Tagatose:properties,applications,and biotechnological processes[J].Applied Microbiology and Biotechnology,2007,76(1):1 -8.
[2]MU WM,ZHANG WL,FENG YH,et al.Recent advances on applications and biotechnological production of D-psicose[J].Applied Microbiology and Biotechnology,2012,94(6):1 461-1 467.
[3]林清泉,刘有才,李丽峰,等.稀有糖的制备及应用最新进展[J].食品与发酵工业,2013,39(6):146-151.
[4]Ragupathi R,Kannan R,Arumugam R,et al.Chemical composition and antibacterial activity of Indian seagrasses against urinary tract pathogen [J].Food Chemistry,2012,135(4):2 470-2 473.
[5]Kopper S,Freimund S.The composition of keto aldoses in aqueous solution as determined by NMR spectroscopy[J].Helvetica Chimica Acta,2003,86(3):827 -843.
[6]Iga Y,Nakamichi K,Shirai Y,et al.Acute and sub-chronic toxicity of D-allose in rats[J].Bioscience Biotechnology and Biochemistry,2010,74(7):1 476 -1 478.
[7]Poulsen T S,Chang Y Y,Hove-Jensen B.D-Allose catabolism of Escherichia coli:involvement of alsI and regulation of als regulon expression by allose and ribose[J].Applied Microbiology and Biotechnology,1999,181(22):7 126 -7 130.
[8]Lim Y R,Oh D K.Microbial metabolism and biotechnological production of D-allose [J].Applied Microbiology and Biotechnology,2011,91(2):229 -235.
[9]Hoshikawa H,Indo K,Mori T,et al Enhancement of the radiation effects by D-allose in head and neck cancer cells[J].Cancer Letters,2011,306(1):60 -66.
[10]Yamaguchi F,Kamitori K,Sanada K,et al.Rare sugar DAllose enhances anti-tumor effect of 5-fluorouracil on the human hepatocellular carcinoma cell line HuH-7[J].Journal of Bioscience and Bioengineering,2008,106(3):248-252.
[11]Kimura S,ZHANG GX,Nishiyama A,et al.D-allose,an all-cis aldo-hexose,suppresses development of salt-induced hypertension in Dahl rats[J].Journal of Hypertension,2005,23(10):1 887 -1 894.
[12]Ishihara Y,Katayama K,Sakabe M,et al.Antioxidant properties of rare sugar D-allose:Effects on mitochondrial reactive oxygen species production in Neuro2A cells[J].J Biosci Bioeng,2011,112(6):638 -642.
[13]SUN YX,Hayakawa S,Chuamanochan M,et al.Antioxidant effects of Maillard reaction products obtained from ovalbumin and different D-aldohexoses[J].Bioscience Biotechnology and Biochemistry,2006,70(3):598 -605.
[14]GAO D,Kawai N,Nakamura T,et al.Anti-inflammatory effect of D-allose in cerebral ischemia/reperfusion injury in rats[J].Neurologia Medico-Chirurgica,2013,53(6):365-374.
[15]SUI L,Nomura R,DONG Y,et al.Cryoprotective effects of D-allose on mammalian cells[J].Cryo-biology,2007,55(2):87-92.
[16]Harada M,Kondo E,Hayashi H,et al.D-Allose and D-psicose reinforce the action of metronidazole on trichomonad[J].Parasitology Research,2012,110(4):1 565 -1 567.
[17]Tanaka S,Sakamoto H.Effects of D-allose on the endocytic activity of dendritic cells and the subsequent stimulation of T cells[J].Cellular Immunology,2011,271(1):141-146.
[18]张卫红.D-阿洛糖及衍生物的合成与应用研究[D].天津:天津大学化工学院,2004.
[19]Mehta G,Pallavi K.The hexose sugar bets-allose and its 2C-brnached homologue from cyclotetraene [J].Tetrahedron Letter,2004,45(20):3 865 - 3 867.
[20]Ryamond R,Herber.Preparation of high purity D-allose from D-glucose[P].U.S.P,5433793,1995.
[21]Hossain B S,Itami Y,Izumori K.Isolation of an L-rhamnose isomerase-constitutive mutant of Pseudomonas sp.strain LL172:Purification and characterization of the enzyme[J].Journal of Fermentation and Bioengineering,1997,84(4):319 -323.
[22]Poonperm W,Takata G,Okada H,et al.Cloning,sequencing,overexpression and characterization of L-rhamnose isomerase from Bacillus pallidus Y25 for rare sugar production [J].Applied Microbiology and Biotechnology,2007,76(6):1 297 -1 307.
[23]LIN CJ,Tseng W C,LIN TH,et al.Characterization of a thermophilic L-Rhamnose isomerase from Thermoanaerobacterium saccharolyticum NTOU1[J].Journal of Agricultural and Food Chemistry,2010,58(19):10 431 -10 436.
[24]LIN CJ,Tseng W C,FANG TY.Characterization of a thermophilic L-rhamnose isomerase from Caldicellulosiruptor saccharolyticus ATCC 43494[J].Journal of Agricultural and Food Chemistry,2011,59(16):8 702 -8 708.
[25]Yoon R Y,Yeom S J,Kim H J,et al.Novel substrates of a ribose-5-phosphate isomerase from Clostridium thermocellum[J].Journal of Biotechnology,2009,139(1):26 -32.
[26]Park C S,Yeom S J,Kim H J,et al.Characterization of ribose-5-phosphate isomerase of Clostridium thermocellum producing D-allose from D-psicose[J].Biotechnology Letters,2007,29(9):1 387 -1 391.
[27]Yeom S J,Seo E S,Kim Y S,et al.Increased D-allose production by the R132E mutant of ribose-5-phosphate isomerase from Clostridium thermocellum[J].Applied Microbiology and Bio-technology,2011,89(6):1 859 -1 866.
[28]FENG ZP,MU WM,JIANG B.Characterization of ribose-5-phosphate isomerase converting D-psicose to D-allose from Thermotoga lettingae TMO[J].Biotechnology Letters,2013,35(5):719 -724.
[29]Yeom S J,Kim B N,Park C S,et al.Substrate specificity of ribose-5-phosphate isomerases from Clostridium difficile and Thermotoga maritim [J].Biotechnology Letters,2010,32(6):829 -835.
[30]Park H Y,Park C S,Kim H J,et al.Substrate specificity of a galactose 6-phosphate isomerase from Lactococcus lactis that produces D-allose from D-psicose[J].Journal of Biotechnology,2007,132(1):88 -95.
[31]邢庆超,沐万孟,江波,等.D-阿洛酮糖的分离纯化[J].食品工业科技,2011,32(9):236 -238.
[32]冯再平,沐万孟,江波,等.生物转化法产D-阿洛糖的分离纯化[J].食品工业科技,2014,35(22):304-307;316.
[33]韩文佳,朱玥明,柏玮,等.双酶偶联转化果糖制备含有稀少糖的混合糖液[J].生物工程学报,2014,30(1):90-97.
