云南省普洱市传统发酵豆豉微生物的群落多样性*
2015-11-20柳陈坚刘晓峰张海燕罗义勇李晓然
柳陈坚,刘晓峰,张海燕,罗义勇,李晓然
(昆明理工大学生命科学与技术学院,云南 昆明,650500)
豆豉以大豆或黄豆为主要原料,利用毛霉、曲霉或者细菌蛋白酶的作用,分解大豆蛋白质,达到一定程度时,加盐、加酒、干燥等方法,抑制酶的活力,延缓发酵过程而制成。豆豉不仅含有大豆中原有的丰富营养素,而且通过微生物发酵作用又产生很多种对人体有极高保健作用的功能性物质,因此其具有较高的营养价值,易于消化吸收[1]。
目前,云南地区的豆豉是以家庭作坊式为主,是开放式自然制曲,因此在发酵过程中不仅有主要参与发酵过程的微生物,还伴随着其他次要微生物的生长。而得益于云南独特的地理位置以及气候环境,使得豆豉中具有许多独特而宝贵的微生物资源。最初研究豆豉中微生物群落多样性主要采用传统微生物培养法,这也是实验室最常用的一种方法。但是自然界中存在许多独特而难培养的微生物,利用微生物培养法只能分离到自然界中极少数的微生物,并不能真实地反映环境中微生物的群落结构,这成为微生物资源开发利用的一个限制性因素,使得微生物的多样性资源难以得到全面开发和利用。近年来,利用不依赖培养的方法研究微生物群落多样性的实验越来越普遍。基于16S rDNA的分子生物学技术的兴起对依赖培养的技术进行了补充,并得到长足的发展。在微生物群落多样性的研究方法不断改进的同时,测序技术也取得了巨大进展。随着测序技术的发展,被称为“第二代测序(next-generation sequencing,NGS)”技术在过去几年中不断涌现,主要包括罗氏454公司的GS-FLX测序平台、Illumina公司的Solexa Genome Analyzer测序平台和ABI公司的Solid测序平台[2]。第二代测序技术高通量,低成本,可以同时对多个单独的DNA分子进行测序分析的特点为研究微生物群落多样性提供了极大的便利。现在高通量测序技术越来越广泛地应用到微生物群落多样性的分析中,尤其以焦磷酸测序(pyrosequencing)的应用最为普遍。
本研究通过不依赖培养的分子生物学方法并结合高通量测序研究了云南省普洱市的豆豉微生物群落多样性,为进一步利用和开发豆豉中微生物资源及发酵食品的质量与安全奠定基础。
1 材料与方法
1.1 材料与试剂
豆豉购于云南省普洱市菜市场,为当地百姓自行发酵制成。购买后置于冰盒中运送到实验室,-80℃保存。
DNA提取裂解缓冲液:0.1 mol/L Tris/HCl,0.1 mol/L EDTA,0.75 mol/L蔗糖;TE缓冲液:10 mmol/L Tris-HCl,pH 8.0,1 mmol/L EDTA ,pH 8.0;Premix Ex TaqTM大连TaKaRa公司;QIAquick Gel Extraction Kit德国Qiangen公司;有机溶剂均为国产分析纯。
1.2 仪器与设备
Sigma 3-18K离心机,美国Sigma公司;ABI 7200PCR仪,美国ABI公司;罗氏GS-FLX测序仪,瑞士Roche公司;Nanodrop ND-1000分光光度计,美国Thermo公司。
1.3 实验方法
1.3.1 DNA提取
DNA提取参考Schmidt等[3]的方法并做了部分改变。取约0.2 g豆豉置于无菌的1.5 mL离心管中,加入450 μL裂解缓冲液,加入10 μL 50 mg/mL 溶菌酶(Lysozyme)和 10 μL 20 mg/mL 溶壁酶(Lyticase),充分混合均匀后,37℃水浴30 min。加入25 μL 20%SDS和5 μL 20 mg/mL蛋白酶K,充分混合均匀后,55℃水浴2 h以上。加入80 μL 5mol/L NaCl和60 μL 10%CTAB/NaCl,小心混匀后,65℃水浴20 min。加入700 μL苯酚、氯仿与异戊醇混合液[V(苯酚)∶V(氯仿)∶V(异戊醇)=25∶24∶1],颠倒混匀后于 12 000 r/min离心10 min,将上层水相转移至新的1.5 mL离心管中,加入等体积的氯仿与异戊醇混合液[V(氯仿)∶V(异戊醇)=24∶1],颠倒混匀后于12 000 r/min离心10 min。将上层水相转移至新的1.5 mL离心管中,加入0.6倍体积的异丙醇,-20℃沉淀过夜后于12 000 r/min离心15 min,倒掉液体,用500 μL预冷的70%乙醇洗沉淀,去掉液体后,打开离心管盖子,放置于60℃烘干箱干燥30 min,每10 min轻弹管壁,促进液体挥发,待液体全部挥发后取出,加入50 μL TE缓冲液溶解DNA。提取好的DNA存放于-20℃冰箱。
1.3.2 细菌16S rDNA和真菌ITS区域扩增及焦磷酸测序
进行DNA扩增的细菌16S rDNA通用引物为338F(5’-ACH YCT ACG GGA GGC HGC-3’)和907R(5’-CCG TCA ATT CMT TTG AGT TT-3’),扩增真菌ITS区域的引物为NSI1(5’-GATTG AATGGCTTAGTGAGG-3’)和58A2r(5’-CTGCGTTCTT CATCGAT-3’)。为了减少PCR产物中非特异性扩增的异源双链DNA的含量,将PCR产物稀释10倍作为模版后按照原来的条件再做5个循环[4]。PCR扩增采用 Premix Ex TaqTM。具体体系及程序:在50 μL的PCR反应体系中:ddH2O 18 μL,Forward 引物(5 μmol/L)2 μL,Reverse 引物(5 μmol/L)2 μL,Premix Ex TaqTM25 μL,模板15 ng。PCR扩增程序为:95℃预变性5 min;94℃变性1 min,56℃复性1 min,72℃延伸1 min,共30个循环;最后72℃延伸7 min,4℃保存。
将PCR产物在10 g/L的琼脂糖凝胶中电泳,电泳后将凝胶浸泡于含有20 μg/mL EB的TAE缓冲液中10 min后在紫外灯(216 nm)照射下呈现橙色条带,目标条带割下后使用QIAquick Gel Extraction Kit对PCR产物进行回收。使用nanodrop ND-1000分光光度计(Thermo,美国)对完成纯化的PCR扩增产物定量。使用GS-FLX(Roche,瑞士)测序系统对DNA进行测序。
1.3.3 测序结果分类鉴定
对于焦磷酸测序的测序结果分析质量文件,去掉测序质量不好的序列以及细菌和真菌序列长度分别小于400 bp和300 bp的序列。根据barcode将序列分配所属的样品中,同时去除barcode和引物序列。对于细菌序列使用 Mothur v1.25.1[5]的 classify.seqs 命令,采用Silva的核糖体小亚基(SSU)rRNA序列数据库V102[6]的分类信息,得到序列的分类信息。对于真菌序列则从每个OTU中选择代表性序列,与NCBI的GenBank进行Blast比对,确定真菌序列的分类信息。
1.3.4 多样性指数和系统发育分析
对所有序列使用Mothur v1.25.1[5]进行比对,以0.03为Cutoff值划分可操作分类单元(operational taxonomic unit,OTU)。计算Cutoff为0.03时的ACE和Chao1值,并绘制稀释曲线。细菌的16S rDNA和真菌ITS区域的在样品中覆盖度用公式C=[1-(n1/N)])×100来计算。其中,n1代表Singleton(只有1条序列的OTU)的数量,N则表示总序列数[7]。
从细菌16S rDNA序列中每个OTU选取代表序列,将其在NCBI的GenBank进行Blast比对,将结果中具有确定分类信息的序列用ClustalX 1.83[8]进行比对,将比对的结果用 Mega v4.0[9]中的 Neighbourjoining方法构建系统发育树。
2 结果与分析
2.1 序列信息和多样性指数
焦磷酸测序后的细菌和真菌序列经过质量和长度删选后,分别得到1 008和3 733条高质量的序列。以0.03为cutoff对这些序列进行分析,共得到细菌OTU 25个,其中,Singleton 14个;真菌OTU 12个,有6个Singleton。样品中细菌和真菌的稀释曲线如图1所示,在cutoff为0.05时,细菌和真菌的稀释曲线均呈现出了接近平台的趋势,而在cutoff为0.03时,曲线的上升趋势还很明显,说明在种的水平上,测序量可能还不能完全反映样品中的微生物群落多样性(图1)。
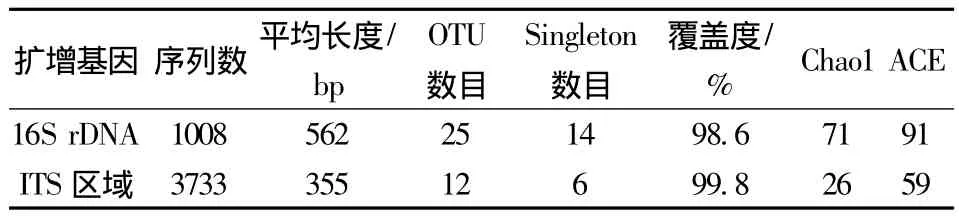
表1 豆豉中细菌和真菌的丰度(cutoff 0.03)Table 1 Bacterial and fungal richness with cutoff of 0.03
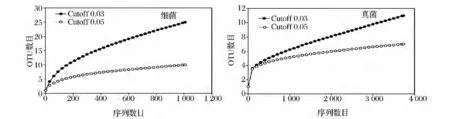
图1 根据Rarefaction方法估计豆豉样品细菌和真菌的多样性Fig.1 Estimated OTU numbers,according to the rarefaction method,depending on OTUs identified with a 3%cut-off
2.2 细菌群落多样性分析
根据Silva的SSUrRNA序列数据库对筛选出的1 008条细菌序列进行分类。经分析,1 008条细菌序列全部属于厚壁菌门(Firmicutes),共检测到厚壁菌门的5个细菌科,分别是:肠球菌科(Enterococcaceae)、葡萄球菌科(Staphylococcaceae)、明串珠菌科(Leuconostocaceae)、肉杆菌科(Carnobacteriaceae)和乳杆菌科(Lactobacillaceae)。其中,占总序列数93%的序列属于肠球菌科,6%的序列属于葡萄球菌科,而属于明串珠菌科、肉杆菌科和乳杆菌科的序列均不足1%,其中以属于乳杆菌科的序列最少,仅有1条。在属的分类水平上,92.5%的序列属于四联球菌属(Tetragenococcus),6%的序列属于葡萄球菌属(Staphylococcus),而属于乳杆菌属(Lactobacillus)、颗粒链菌属(Granulicatella)、蜜蜂球菌属(Melissococcus)和Fructobacillus均不足1%,见图2。

图2 豆豉中细菌在不同分类水平的分布Fig.2 The major families and genera in fermented soybean
2.3 真菌群落多样性分析
以0.03为cutoff,共得到12个OTU,从除Singleton和Doubleton(含有2条序列的OTU)之外的6个OTU中,各选取1条代表性序列,在 NCBI的 Gen-Bank进行Blast比对,确定每个OTU的分类信息。所分析的3 733条真菌序列均属于假丝酵母属(Candida),其中有3个OTU,占总序列数85.5%(3 192条)的序列属于 C.etchellsii,OTU PE_ITS_3(8%)属于C.apicola,还有占总序列数6%(244条)的2个OTU属于皱状假丝酵母(C.versatilis)(表2)。
2.4 细菌的系统发育分析
以0.03为cutoff,确定了样品1 008条细菌序列的25个OTU。除去Singleton和Doubleton后,对剩余的10个OTU与其在GenBank数据库中的相近序列共同构建系统发育树,如图3所示。丰度最高的OTU是P1,占总序列数的86.7%,具有绝对的优势,经比对,OTU P1的序列与嗜盐四联球菌(T.halophilus),具有97%的相似度。有43条序列属于OTU P9,这也是丰度第二高的OTU,属于这个OTU的序列与葡萄球菌属Staphylococcus sp.具有较高的相似度。OTU P7有19条序列,也属于葡萄球菌属,与肉葡萄球菌(S.carnosus)具有较高的相似度。有16条序列划分在OTU P3中,在系统进化树上并未与已知分类信息的细菌呈现较高的相似度。OTU P6有15条序列,与嗜盐四联球菌也具有97%的相似度。另外,其余的五个 OTU占总序列数的比例均 <1%。其中,OTUP2、P5、P8、P10,均与嗜盐四联球菌具有较高的相似度,而OTUP4则与Leuconostoc fallax具有99%的相似度。从系统进化树上可看出,所有序列均属于厚壁菌门。

表2 真菌OTU及其分类信息Table 2 Fungal OTUs and the closest taxonomic annotations
3 讨论
不同类型的发酵食品,其中的微生物群落结构也各有差异,例如,发酵玉米食品的优势细菌属于乳杆菌属,优势真菌是热带假丝酵母(C.tropicalis)[10];发酵可可豆中的优势细菌是乳杆菌属和醋菌属(Acetobacter),优势真菌类型为酿酒酵母(Saccharomyces cerevisiae)和汉森氏酵母属(Hanseniaspora)[11];发酵海产品的优势细菌是乳杆菌属和魏斯氏菌属,还检测到嗜盐古菌的存在[12]。
在本研究中,豆豉的细菌群落多样性明显高于真菌群落的多样性。在豆豉的细菌群落中,丰度相对较高的10个OTU分别属于厚壁菌门的5个菌科。丰度最高的OTU P1及另外4个丰度较低的OTU,都与嗜盐四联球菌具有较高的相似度。嗜盐四联球菌是一种能够适应高盐等极端环境[13]的乳酸菌,嗜盐乳酸菌通常存在于发酵的鱿鱼肝酱[14]、豆瓣酱[15]及咸鱼[16]等食品中。丰度第二高的OTU是P9,经比对属于葡萄球菌,但是并没有具体到种的分类信息。葡萄球菌属中至少有40个不同的菌种,除了有从发酵食品中分离到的,也有一部分潜在致病菌,因此,确定其是否为致病菌需要进一步分离鉴定。这也暗示了豆豉中可能存在潜在的致病风险。与OTUP7相似度最高的是肉葡萄球菌,最初分离自干香肠中,与肉制品的生产也具有一定的关系[17]。2009年德国科学家Rosenstein等[18]从肉品发酵剂中分离到肉葡萄球菌,说明肉葡萄球菌在发酵过程中可能具有某种作用。
豆豉的真菌群落中丰度最高的OTU属于C.etchellsii,另外的2个OTU分别属于C.apicola和C.versatilis,这3株酵母菌都是耐盐菌[19]分离自大豆酱等高盐环境中,对于食品发酵中的香气物质产生具有重要作用[20]。这表明云南省普洱市的豆豉含盐量极高,检测到了极高比例的耐盐菌,因此只有能够适应高盐环境的微生物才能大量繁殖。
普洱市地处热带,不论是豆豉的细菌群落还是真菌群落,其优势菌都是耐盐菌,除了表明豆豉极高的含盐量之外,也暗示了人们在食用此类豆豉时要注意食盐摄入量,避免引起高血压等疾病。多数乳酸菌能够产生细菌素等抗菌物质,可以考虑在制作豆豉过程中通过人为添加产细菌素乳酸菌的方法来减少制作过程中食盐的添加。葡萄球菌属中虽然有的细菌能够参与豆豉的发酵过程,但是并未做具体实验证明它不存在潜在致病性,需要做进一步的研究来确定。
[1]卢露,郑晓莹.豆豉发酵中微生物及其功能研究进展[J]. 粮食与食品工业,2011,18(1):42-45.
[2]贺纪正,袁超磊,沈菊培,等.土壤基因组学研究方法与进展[J].土壤学报,2012,49(1):155-164.
[3]Schmidt T M,DeLong E F,Pace N R.Analysis of a marine picoplankton community by 16S rRNA gene cloning and sequencing[J].Journal of Bacteriology,1991,173(14):4 371 -4 378.
[4]Thompson J R,Marcelino L A,Polz M F.Heteroduplexes in mixed-template amplifications:formation,consequence and elimination by 'reconditioning PCR '[J].Nucleic Acids Research,2002,30(9):2 083 -2 088.
[5]Schloss P D,Westcott S L,Ryabin T,et al.Introducing mothur:open-source,platform-independent, communitysupported software for describing and comparing microbial communities[J].Applied and Environmental Microbiology,2009,75(23):7 537 -7 541.
[6]Pruesse E,Quast C,Knittel K,et al.SILVA:a comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB[J].Nucleic Acids Research,2007,35(21):7 188 -7 196.
[7]Good I J.The population frequencies of species and the estimation of population parameters[J].Biometrika,1953,40(3-4):237-264.
[8]Thompson J,Gibson T,Plewniak F,et al.The CLUSTAL_X windows interface:flexible strategies for multiple sequence alignment aided by quality analysis tools[J].Nucleic Acids Research,1997,25(24):4 876 -4 882.
[9]Tamura K,Dudley J,Nei M,et al.MEGA4:Molecular evolutionary genetics analysis(MEGA)software version 4.0[J].Molecular Biology and Evolution,2007,24(8):1 596 -1 599.
[10]Obinna-Echem P C,Kuri V,Beal J.Evaluation of the microbial community,acidity and proximate composition of akamu,a fermented maize food[J].Science of Food and Agriculture,2013,94(2):331 -340.
[11]de Melo Pereira G V,Magalh~aes-Guedes K T,Schwan R F.rDNA-based DGGE analysis and electron microscopic observation of cocoa beans to monitor microbial diversity and distribution during the fermentation process[J].Food Research International,2013,53(1):482 -486.
[12]RohS W,KimK H,Nam Y D,et al.Investigation of archaeal and bacterial diversity in fermented seafood using barcoded pyrosequencing[J].The ISME Journal,2010,4(1):1 -16.
[13]Juste A,Lievens B,Frans I,et al.Genetic and physiological diversity of Tetragenococcus halophilus strains isolated from sugar-and salt-rich environments[J].Microbiology-Sgm,2008,154(9):2 600-2 610.
[14]Satomi M,Kimura B,Mizoi M,et al.Tetragenococcus muriaticus sp.nov.,a new moderately halophilic lactic acid bacterium isolated from fermented squid liver sauce[J].I nternational Journal of Systematic Bacteriology,1997,47(3):832-836.
[15]Tanasupawat S,Thongsanit J,Okada S,et al.Lactic acid bacteria isolated from soy sauce mash in Thailand[J].Journal of General and Applied Microbiology,2002,48(4):201-209.
[16]Villar M,de Ruiz Holgado A P,Sanchez J J,et al.Isolation and characterization of Pediococcus halophilus from salted anchovies(Engraulis anchoita)[J].Applied and Environmental Microbiology,1985,49(3):664 -666.
[17]Schleifer K H,Fischer U.Description of a new species of the genus Staphylococcus:Staphylococcus carnosus[J].International Journal of Systematic Bacteriology,1982,32(2):153-156.
[18]Rosenstein R,Nerz C,Biswas L,et al.Genome analysis of the meat starter culture bacterium Staphylococcus carnosus TM300[J].Applied and Environmental Microbiology,2009,75(3):811 -822.
[19]Ozaki S,Fukuda S,Fujita T,et al.RAPD analysis of salttolerant yeasts from contaminated seasoned pickled plums and their growth inhibition using food additives[J].Biocontrol Science,2008,13(4):125 -130.
[20]Suezawa Y,Kimura I,noue M,et al.Identification and typing of miso and soy sauce fermentation yeasts,Candida etchellsii and C.versatilis,based on sequence analyses of the D1D2 domain of the 26S ribosomal RNA gene,and the region of internal transcribed spacer 1,5.8S ribosomal RNA gene and internal transcribed spacer 2[J].Bioscience Biotechnology and Biochemistry,2006,70(2):348 -354.
