甘油发酵生产3-羟基丙酸的代谢改造工程菌研究进展*
2015-11-19牛坤秦海彬柳志强郑裕国
牛坤,秦海彬,柳志强,郑裕国
(浙江工业大学生物与环境工程学院,浙江 杭州,310014)
随着煤炭石油等不可再生资源的日益减少,潜力巨大的燃油生物炼制技术受到了越来越多的关注,其中,生物柴油产业更是在近几年内迅速发展。然而,每生产10 t生物柴油就会产生1 t甘油[1],随着全球各国生物柴油产业不断发展,必将对国际甘油市场形成极大的冲击。为了解决这一问题,业界兴起了以甘油为原料生产高附加值化学品的热潮。
甘油具有特殊的物理性质和化学结构,广泛应用于医药、食品、化妆品以及工业化工产品等领域[2]。而研发甘油的革新用途,建立其下游化学品开发利用的有效途径,也成为近年来科学家不断探索的新问题,当前利用甘油的有效途径包括:生产乙醇、制备1,3-丙二醇、合成环氧氯丙烷、生产乙二醇、生产乳酸、合成二羟基丙酮及3-羟基丙酸等[3]。
3-羟基丙酸(3-hydroxypropionic acid,3-HP)是一种无色无味的油状液体,可与水、醇、醚等多种溶剂互溶。3-HP为三碳无手性有机化合物,具有羧基和羟基两种官能团,这使得3-HP具有优良的化学性质,如脱水生成丙烯酸,氧化生成有机酸等。同时3-HP也是一种重要的化学中间体,可以用来合成很多重要的工业化工产品,如涂料、胶黏剂、水处理化学品和个人护理用品等。由于3-HP显著的市场价值及工业应用前景,2004年美国能源部报告将3-HP列为当今世界上12种最具有开发潜力的化工产品之一[4]。
1 以甘油为底物合成3-HP的主要途径及关键酶
目前,市售的3-HP均由化学合成法制得[5-6]。具体方法包括:由β-羟基丙腈与NaCN水解反应制备3-HP;丙烯酸在高温或液酸催化下生成3-HP;金属铂催化3-羟基丙醛生成3-HP等。但是,以上途径中原材料多为非可再生资源,且易造成环境不兼容、成本高、产品分离纯化复杂等问题,因此依靠上述途径进行3-HP的大规模生产存在一定的局限性。
随着基因工程技术的发展,生物法替代化学法合成大宗化学品已经得到广泛的研究,在此基础上,研究学者尝试采用生物法合成3-HP。目前生物法合成3-HP主要有两种途径,一是以葡萄糖为底物;二是以甘油为底物。甘油在微生物体内可以通过氧化反应和还原反应2种方式进行代谢。如图1所示,在氧化途径中,甘油首先经过甘油脱氢酶 (GDH)催化生成二羟基丙酮(DHA),然后进一步代谢生成磷酸二羟丙酮(DHAP),最终进入糖酵解途径生成丙酮酸,并为菌体的生长提供所需要的能量ATP和还原力NAD(P)H。在还原途径中,甘油首先经过甘油脱水酶(DhaB)与辅酶 VB12的作用,脱水生成3-羟基丙醛(3-HPA),再由 1,3-丙二醇氧化还原酶 (DhaT/YqhD)还原为产物1,3-丙二醇 (1,3-PDO),同时消耗了氧化途径中所生成的NAD(P)H。而在甘油还原生成3-HPA后,可进一步在醛脱氢酶 (AldH)的催化下以NAD+为辅因子生成3-HP。
甘油脱水酶(DhaB)是生物合成3-HP的第一个关键酶。该酶为辅酶VB12依赖型,在厌氧或微氧条件下具有较高的活性。醛脱氢酶(AldH)是影响3-HP合成的第二个关键酶,该酶为辅酶NAD+依赖型,在氧气充足的条件下具有较高活性,其活性的高低将显著影响3-HP的产量。因此,目前在醛脱氢酶的筛选上做了大量的研究工作 (表1),例如来源于Klebsiella pneumoniaeDSM 2026的醛脱氢酶PuuC[8]、 来源于巴西固氮螺菌 (Azospirillum brasilense)的 α-酮戊二酸半醛脱氢酶 KGSADH[9]、来源于罗伊氏乳杆菌(Lactobacillus reuteri)的丙醛脱氢酶 PduP 等[10]。
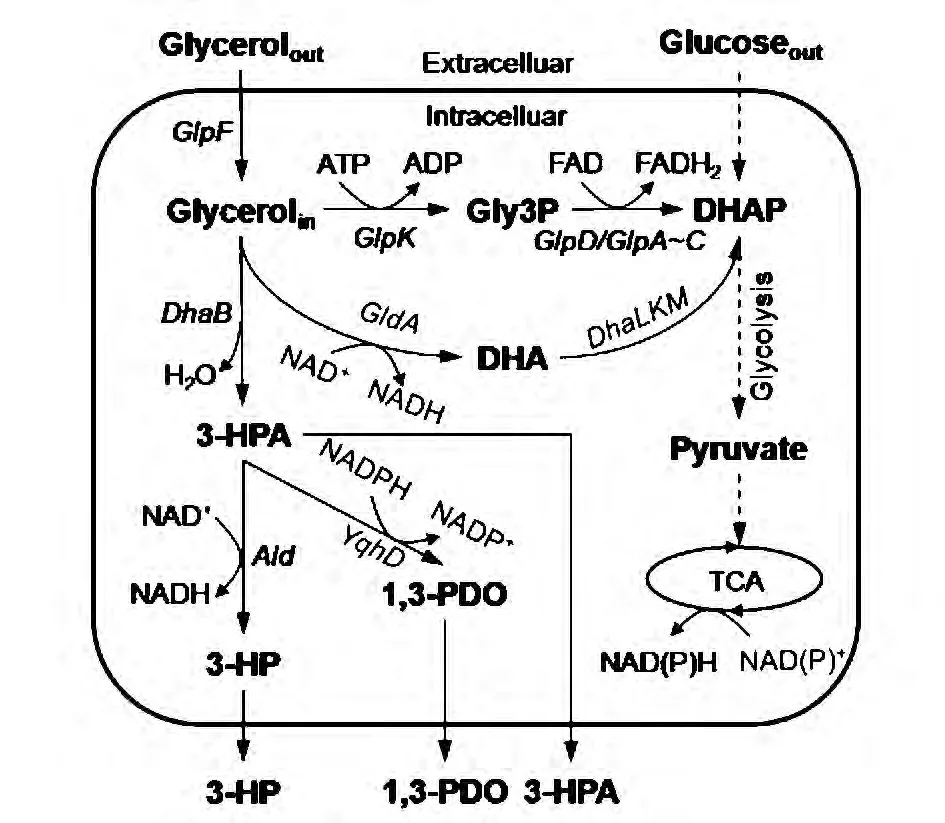
图1 甘油在微生物体内的主要代谢路径[7]Fig.1 The main metabolic pathway of glycerol in microorganisms
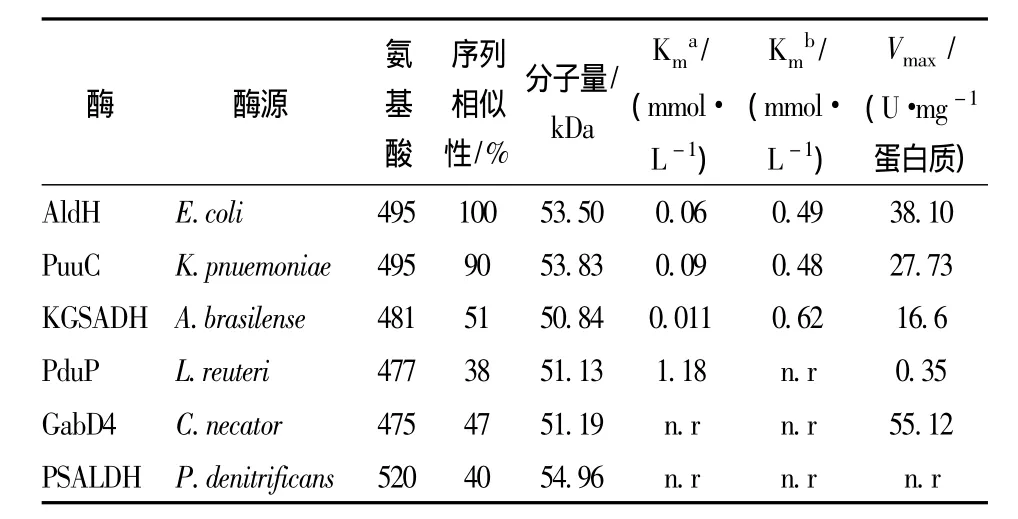
表1 已报道的醛脱氢酶Table 1 Some reported aldehyde dehydrogenase
2 3-羟基丙酸生物合成的代谢工程改造
早在20世纪60年代研究人员就开始了微生物发酵生产3-HP的研究。2001年,Suther等人首次报道了以甘油为底物,采用基因工程菌两步法合成3-HP的研究[11]。他们克隆了来源于克雷伯氏肺炎杆菌的dhaB基因,以及分别来源于大肠杆菌、人类和酿酒酵母的4种醛脱氢酶基因。通过实验发现酿酒酵母的醛脱氢酶基因ald4效果最好,虽然最终3-HP产量仅为0.17 g/L,但为以甘油为底物生产3-HP的研究拉开了序幕。随着甘油脱水酶和醛脱氢酶的不断发现,愈来愈多的重组菌株被报道,其中尤以E.coli和K.pneumoniae为宿主菌研究最多。
2.1 以E.coli作为宿主合成3-HP
E.coli是开发最早的一种模式生物,它具有清楚的遗传背景,易于改造,且生长条件简单,对营养物质要求低,因此被广泛用于发酵生产许多高价值化工产品。就3-HP的合成来说,因其自身含有醛脱氢酶基因,因此采用E.coli作为宿主备受青睐。
表2所示为近年来以E.coli为宿主合成3-HP的主要研究,其中韩国Park团队在此方面的工作尤为突出[8-9,12-15]。2008 年,Raj 等在 Suthers 的研究基础上克隆了来源于K.pneumoniaeDSM 2026的甘油脱水酶基因dhaB和E.coliK-12的醛脱氢酶基因aldH,构建了1株以甘油为底物生产3-HP的基因工程菌E.coliSH254。该菌以甘油 (100 mmol/L)为唯一碳源发酵30 h后,3-HP产量为0.58 g/L,甘油脱水酶活力为37 U/mg,醛脱氢酶活力为22.8 U/mg,得率为 0.48 mol/mol[12]。后续通过对摇瓶培养过程中pH、IPTG浓度、装液量、底物浓度等条件的优化,使3-HP的摇瓶产量提高到4.4 g/L;发酵罐中发酵72 h后,3-HP产量为31 g/L,得率为0.35 mol/mol,最大产率达 3.41 mmol/(g cell·h)[13]。
通过上述研究,Park团队发现3-HP产量较低主要是由于甘油脱水酶的不稳定性以及2种关键酶活性的不平衡造成的。为了稳定甘油脱水酶的活性,该团队克隆了甘油脱水酶再激活酶基因gdrAB,并将此基因与dhaB基因进行了共表达,同时筛选了新的醛脱氢酶α-酮戊二酸半醛脱氢酶KGSADH。实验结果表明含有dhaB-gdrAB和kgsadh基因的重组菌SHBGK1经过5 L发酵罐培养后,3-HP产量达到38.7 g/L。这一产量虽然较前人的研究有了较大的提高,但是过程中甘油转化率仍然过低,仅为35%,主要原因还是醛脱氢酶活性偏低,导致双酶表达不均衡,中间代谢产物3-羟基丙醛(3-HPA)积累产生细胞抑制毒性造成的[9]。

表2 以E.coli为宿主合成3-HPTable 2 3-HP production by recombinant E.coli
因此筛选高活力醛脱氢酶并对其进行分子改造,同时敲除3-HP的竞争途径也是近年来提高产量和甘油转化率的主要策略。Kim等克隆了来源于铜绿假单胞菌(Pseudomonas aeruginosa)的半醛脱氢酶PSALDH以及来源于短乳杆菌(Lactobacillus brevis)的甘油脱水酶DhaB-DhaR,构建了重组菌株E.coliBL21 star(DE3),同时敲除了甘油竞争途径中合成3-磷酸甘油的glpK基因和催化3-HPA合成1,3-PDO的yqhD基因,使3-HP产量大幅度提高达到57.3 g/L[7]。Chu等克隆了来源于钩虫贪铜菌(Cupriavidus necator)的新型醛脱氢酶GabD4,发现该酶对中间产物3-HPA的催化活性是目前所报道的最高值,而后续又对其进行了定点和饱和突变,使其活性提高到原始酶的1.4倍,敲除途径中乙酰辅酶A合成乙酸的关键基因ackA-pta以及yqhD基因后,3-HP的产量也提高至目前的最高水平71.9 g/L[16]。
目前,国内众多科研院校也相继开展了利用重组大肠杆菌合成3-HP的研究。江南大学张晓梅等构建了同时表达DhaB和AldH的基因工程菌E.coliJM109,该菌在含有50 g/L甘油的发酵液中培养48 h,3-HP产量为4.92 g/L,甘油利用率为50.12%,转化率为12.3%[18]。后续考察了甘油脱水酶激活因子(gdrAB)对3-HP合成量的影响,发现菌株中重组gdrAB后,3-HP合成量提高了6.4倍[19]。南京工业大学的胡南等以自身携带有乙醛脱氢酶的E.coliBL21(DE3)plysS作为宿主,异源表达了来源于K.pneumoniae的dhaB基因,重组菌E.coliHP获得的甘油脱水酶比活力在1 mmol/L IPTG的诱导下达到77.2 U/mg,摇瓶条件下3-HP的产量达到了5.44 g/L,摩尔转化率为53%[20]。
在发酵过程中,甘油除了合成3-HP外,还作为碳源进行同化作用合成细胞,并有部分甘油经过氧化最终生成ATP和还原力。因此,采用E.coli作为宿主最主要的问题还要提高甘油合成3-HP的转化效率。Jung等人除了敲除3-HP的竞争途径外,还研究了甘油的代谢速率对3-HP合成的影响,主要考察了甘油激酶GlpK(将甘油转化为3-磷酸甘油)、甘油促进因子GlpF(促进甘油由胞外到胞内的吸收),以及甘油途径调节因子GlpR(控制甘油的整体代谢途径)等的影响。发现甘油途径调节因子GlpR是影响甘油代谢的主要抑制因素,该途径的敲除可以激活甘油的分解代谢,提高3-HP的合成产量。敲除副产物乙酸和1,3-丙二醇的合成途径后3-HP产量为28.1 g/L,而在此基础上过量表达glpk基因并敲除glpR基因后,甘油的利用速率显著增加,3-HP的产量也提高至40.5g/L;而如果在敲除glpR基因的基础上过量表达glpF基因后,3-HP 的产量会提高至 42.1 g/L[17]。
2.2 以K.pneumoniae作为宿主合成3-HP
在3-HP合成过程中,甘油脱水酶发挥催化活性需要辅酶VB12的参与,而K.pneumoniae自身能够在微氧或厌氧的条件下合成VB12,这在很大程度上解决了昂贵的辅酶添加问题,降低了生产成本。并且K.pneumoniae自身含有甘油脱水酶基因,可以直接利用甘油合成3-HPA,因此,采用K.pneumoniae作为宿主合成3-HP的研究也越来越多。K.pneumoniae遗传背景与进化方向与大肠杆菌相近,可相互进行基因操作。另外,与E.coli相比,K.pneumoniae代谢利用甘油能力较强,同化率高,无需额外添加碳源,且甘油途径得到了详细的解析。
在K.pneumoniae以甘油为底物进行发酵的过程中会生成另一种重要的化工产品1,3-PDO,因此以K.pneumoniae作为宿主的另一个优势是可以实现两种重要化工产品的联产,且3-HP和1,3-PDO联产可实现辅因子NAD+的再生平衡[21]。国内华东理工大学在此方面的工作较为突出,HUANG等构建重组菌K.pneumoniae成功表达了E.coli的aldH基因,在5 L发酵罐中发酵24 h后,3-HP和1,3-PDO的产量分别达到24.4 g/L和49.3 g/L,甘油的总转化率达到0.61 mol/mol,实验证明在不添加VB12情况下可以成功合成3-HP和1,3-PDO[22]。如前所述,甘油脱水酶与醛脱氢酶对氧气的需求是不同的,为了平衡两者的活性,HUANG等人还考察了不同的通气量和搅拌转速对3-HP合成的影响,发现在微氧的条件下 (1.5 vvm)适当提高搅拌转速可以使3-HP产量增加至48.9g/L,而使 1,3-PDO 的产量降低至 25.3g/L[23]。后续该课题组对已构建的重组菌K.pneumoniae(pUC18kan-aldHec)以甘油为底物生产3-HP和1,3-PDO的过程进行发酵过程的优化,发现补料阶段控制甘油浓度在20~30 g/L,发酵26 h后3-HP和1,3-PDO的产量分别达到47.2 g/L和43.9 g/L;而间歇性补加甘油时,产物得率最高,发酵26 h后,甘油合成3-HP和1,3-PDO的总转化率可以达到0.73 mol/mol[21]。

表3 以K.pneumoniae作为宿主合成3-HPTable 3 3-HP production by recombinant K.pneumoniae
在以K.pneumoniae为宿主合成3-HP的过程中,敲除副产物的代谢途径依然是研究的重要方向。Luo等人通过灭活K.pneumoniae中的甘油脱氢酶使代谢过程中氧化途径缺失,同时将NAD+依赖型的醛脱氢酶AldHk克隆到宿主菌中,发现3-HP的产量可以提高3.6倍[24]。Ashok等人还考察了甘油脱水酶与醛脱氢酶的酶活对氧气需求不匹配的问题,他们将NAD+依赖型的新型醛脱氢酶 PuuC(γ-谷氨酰-γ-氨基丁醛脱氢酶)克隆于K.pneumoniae中,该重组菌可以在微氧条件下有效地将3-HPA转化为3-HP和1,3-PDO,而PuuC为NAD+辅酶依赖型,微氧条件会提高NADH的量而不利于NAD+的循环,并且在以氧气为电子受体的NADH/NAD+辅酶循环过程中会伴随大量的1,3-PDO和乳酸生成,因此在发酵过程中尝试采用硝酸盐作为NADH的电子受体来完成NAD+的循环,结果表明硝酸盐的加入可以提高NAD+的循环量,但是由甘油合成3-HP的碳流并没有增加,后续又通过对影响甘油代谢速率和1,3-PDO产量的相关基因glpK和dhaT(1,3-丙二醇氧化还原酶)进行敲除,使得甘油合成3-HP的转化率提高到30%,发酵罐中 3-HP 的产量提高到 22.5g/L[15,25]。而继续敲除yqhD基因后,增大了由3-HPA流向3-HP碳流,在发酵过程中优化通气量,以5%的溶氧量发酵48 h后3-HP产量提高至28 g/L[26]。除了寻找NADH的电子受体外,在体内进行多酶共表达进行辅因子NAD(P)+再生也是一个主要方向,曹宁等引入酿酒酵母(S.cerevisiae)的3-磷酸甘油脱氢酶基因GPD1,得到耦合3-HP合成与NAD+再生的重组菌K.pneumoniae(pET-pk-dhaB-puuC-GPD1),微氧条件下,摇瓶中发酵15 h后,3-HP产量达到1.45 g/L,是携带空载体菌的2.39倍[27]。
在以K.pneumoniae为宿主合成3-HP的研究中,还有学者尝试采用静息细胞对底物甘油进行转化,该方法的优点是可以在有氧条件下培养活细胞,然后在微氧或无氧条件下利用甘油进行转化合成3-HP。Kumar等以kgsadh基因作为醛脱氢酶基因构建重组菌K.pneumoniaeJ2B,采用静息细胞催化法在1.5 L反应器中厌氧分批转化12 h后,3-HP质量浓度为11.3 g/L,1,3-PDO质量浓度为15.2 g/L,总转化效率可以达到0.71 mol/mol。但是除了上述产物之外还有大量乳酸产生,因此在后续的工作中将该重组菌中的ldhA(乳酸脱氢酶)基因进行敲除,同样采用静息细胞转化后,3-HP和1,3-PDO产量分别提高到22.7 g/L 和 23.5 g/L[28-29]。

续表3
2.3 其他宿主
由甘油合成3-HPA的关键酶甘油脱水酶为辅酶VB12依赖型,因此,筛选能够自身合成VB12的菌株,减少辅酶添加,降低发酵成本,也是目前的一个研究方向。脱氮假单胞菌(Pseudomonas denitrificcans)是目前工业生产VB12的常用菌株,该菌株可以在有氧条件下自身合成辅酶VB12,同时有氧条件下辅酶NAD+也能够进行有效地再生,是3-HP合成的理想宿主。
Park团队将前述的甘油脱水酶及其再激活酶基因(dhaB/gdrAB)以及来源于K.pneumoniae的醛脱氢酶基因puuC共表达于P.denitrificans宿主细胞中构建了基因工程菌株,发现该重组菌可以葡萄糖作为碳源和能源进行生长,而在培养基中添加甘油后可以产生5 g/L左右的3-HP,虽然3-HP的产量不高,但是为以甘油为底物合成3-HP提供了新的研究方向[30]。后续该团队也在VB12产生菌的筛选方面做了一些工作,筛选到1株在好氧条件下合成VB12的菌株Pseudomonassp.SP2,该菌株也为后续3-HP的研究提供了基础[31]。
3 研究展望
目前,以甘油为底物发酵生产3-HP的研究中,3-HP的最高产量为71.9 g/L,而据文献报道有机酸的商业化生产需要达到100 g/L以上[32]。因此,为了尽快实现3-HP的工业化生产,需要深入研究甘油生产3-HP途径中的限制性因素,提高关键酶的活性及底物转化率。在以甘油为底物,利用甘油脱水酶将甘油转化为3-羟基丙醛,醛脱氢酶进一步将3-羟基丙醛氧化为3-羟基丙酸的途径中,存在着甘油脱水酶不稳定性和自杀性灭活、醛脱氢酶酶活力偏低、两种酶对氧气的需求不匹配、中间产物3-HPA的累积毒性、产物3-HP积累毒性以及VB12供给和辅酶循环再生等问题。其中,甘油脱水酶得到了深入的研究,表达甘油激活因子解决了它的不稳定性和自杀性灭活问题;甘油脱水酶DhaB蛋白质晶体结构的确定为它的分子改造提供了理论基础。而低活力的醛脱氢酶作为限速步骤,限制了3-HP的产量,导致中间产物3-HPA的累积,3-HPA对细胞具有抑制毒性,因此继续筛选高活力醛脱氢酶,使3-HPA迅速转化为3-HP还是今后研究的重要方向。获得适应性强、酶活高、稳定性好的醛脱氢酶,除了定向进化、理性设计和半理性设计等方法外,还可以采用宏基因组学进行寻找。內源辅酶NAD(P)+浓度和再生对醛脱氢酶活性和3-HP产量有着不可忽略的影响,在今后的研究中我们可以参考其他氧化还原酶的一些解决策略来提高NAD(P)+的循环量,如多酶共表达进行辅酶再生或者电化学等方法,而解决NAD(P)H/NAD(P)+高比例对酶活力抑制问题也需要进一步的研究,如寻找适当的NADH电子受体等。近几年的研究使3-HP的产量得到很大的提高,菌株能否耐受高3-HP浓度将成为3-HP工业化生产的关键,高耐受菌株的选育或基因缺陷型菌株的发现也必将成为今后研究的重点。
[1] Meesters PAEP,Huijberts GNM,Eggink G.High-cell-density cultivation of the lipid accumulating yeastCryptococcus curvatususing glycerol as a carbonsource[J].Applied Microbiology and Biotechnology,1996,45(5):575 - 579.
[2] Silva GPD,Mack M,Contiero J.Glycerol:A promising and abundant carbon source for industrial microbiology[J].Biotechnology Advances,2009,27(1):30 -39.
[3] Alba E D,Javier F,Beatriz L B,et al.Glycerol and derived solvents:new sustainable reaction media for organic synthesis[J].Chemical Communications,2011,22(47):6 208-6 227.
[4] Werty T,Petersen G.Top value added chemicals from biomass[C].Energy Efficiency and Renewable Energy,2004.
[5] 楼坚,裘娟萍.生物法合成3-羟基丙酸的研究进展[J].工业微生物,2006,36(4):56-60.
[6] 温丽瑗,陈小平,郎春燕.3-羟基丙酸合成途径研究进展[J].天然气化工,2012,37(4):62-67.
[7] Kim K,Kim S K,Park Y C,et al.Enhanced production of 3-hydroxypropionic acid from glycerol by modulation of glycerol metabolism in recombinantEscherichia coli[J].Bioresource Technology,2014,156:170 -175.
[8] Raj S M,Rathnasingh C,Jung W C,et al.A Noveldependent aldehyde dehydrogenase encoded by thepuuC gene ofKlebsiella pneumoniaeDSM 2026 that utilizes 3-hydroxypropionaldehyde as a substrate[J].Biotechnology and Bioprocess Engineering,2010,15(1):131 -138.
[9] Rathnasingh C,Raj S M,Jo J E,et al.Development and evaluation of efficient recombinantEscherichia colistrains for the production of 3-hydroxypropionic acid from glycerol[J].Biotechnologyand Bioengineering,2009,104(4):729-739.
[10] LUO LH,Seo J W,Baek J O,et al.Identification and characterization of the propanediol utilization protein PduP ofLactobacillus reuterifor 3-hydroxypropionic acid production from glycerol[J].Applied Microbiology and Biotechnology,2011,89(3):697 -703.
[11] Suthers P F,Madisn W I,Cameron D C,et al.Production of 3-hydroxypropionic acid in recombinant organism[P].United States Patent,6852517.2005.
[12] Raj S M,Rathnasingh C,Jo J E,et al.Production of 3-hydroxypropionic acid from glycerol by a novel recombinantEscherichia coliBL21 strain[J].Process Biochemistry,2008,43(12):1 440 -1 446.
[13] Raj S M,Rathnasingh C,Jung W C,et al.Effect of process parameters on 3-hydroxypropionic acid production from glycerol using a recombinantEscherichia coli[J].Applied Microbiology and Biotechnology,2009,84(4):649 -657.
[14] Jo J E,Raj S M,Rathnasingh C,et al.Cloning,expression,and characterization of an aldehyde dehydrogenase fromEscherichia coliK-12 that utilizes 3-Hydroxypropionaldehyde as a substrate[J].Applied Microbiology and Biotechnology,2008,81(1):51 -60.
[15] Ashok S,Subramanian R M,Chelladurai R,et al.Development of recombinantKlebsiella pneumoniaeΔdhaT strain for the co-production of 3-hydroxypropionic acid and 1,3-propanediol from glycerol[J].Applied Microbiology and Biotechnology,2011,90(4):1 253-1 265.
[16] Chu HS,Young SK,Chan ML,et al.Metabolic engineering of 3-hydroxypropionic acid biosynthesis inEscherichia coli[J].Biotechnology and Bioengineering,2015,112(2):356-364.
[17] Minty J J,Lesnefsky A A,Lin F,et al.Elevated production of 3-hydroxypropionic acid by metabolic engineering of the glycerol metabolism inEscherichia coli[J].Metabolic Engineering,2014,23:116 -122.
[18] 张晓梅,诸葛斌,许正宏.产3-羟基丙酸重组菌的构建及其转化甘油的研究[J].生物技术通报,2009(8):104-108.
[19] 权国燕,方慧英,诸葛斌,等.甘油脱水酶再激活因子提高重组大肠杆菌3-羟基丙酸合成能力[J].中国生物工程杂志,2011,31(6):75 -80.
[20] 胡南,缪鑫昕,谈曙明,等.重组大肠杆菌生物转化甘油生产3-羟基丙酸[J].生物技术,2012,22(1):72-75.
[21] 李清,黄艳娜,李志敏,等.重组肺炎克雷伯菌发酵联产3-羟基丙酸和1,3-丙二醇[J].过程工程学报,2014,14(1):133-138.
[22] HUANG Y,LI Z,Shimizu K,et al.Simultaneous production of 3-hydroxypropionic acid and 1,3-propanediol from glycerol by a recombinant strain ofKlebsiella pneumoniae[J].Bioresource Technology,2012,103(1):351 -359.
[23] HUANG Y,LI Z,Shimizu K,et al.Co-production of 3-hydroxypropionic acid and 1,3-propanediol byKlebseilla pneumoniaeexpressingaldH under microaerobic conditions[J].Bioresource Technology,2013,128:505 - 512.
[24] LUO LH,Seo J W,Oh B R,et al.Stimulation of reductive glycerol metabolism by over expression of an aldehyde dehydrogenase in a recombinantKlebsiella pneumoniaestrain defective in the oxidative pathway[J].Journal of Industrial Microbiology and Biotechnology,2011,38(8):991-999.
[25] Ashok S,Mohan R S,Ko Y,et al.Effect of puuC over expression and nitrate addition on glycerol metabolism and anaerobic 3-hydroxypropionic acid production in recombinantKlebsiella pneumoniaeΔglpK ΔdhaT[J].Metabolic Engineering,2013,15:10 -24.
[26] Ashok S,Sankaranarayanan M,Ko Y,et al.Production of 3-hydroxypropionic acid from glycerol by recombinantKlebsiella pneumoniaeΔdhaT ΔyqhD which can produce vitamin B12 naturally[J].2013,110(2):511 - 524.
[27] 曹宁,李莎,葛喜珍,等.肺炎克雷伯氏菌3-羟基丙酸合成关键酶基因过表达与NAD+再生的耦合[J].北京化工大学学报:自然科学版,2014(1):74-77.
[28] Kumar V,Mugesh S,Jae K,et al.Co-production of 3-hydroxypropionic acid and 1,3-propanediol from glycerol using resting cells of recombinantKlebsiella pneumoniaeJ2B strain over expressing aldehyde dehydrogenase[J].Applied Microbiology and Biotechnology,2012,96(2):373 -383.
[29] Kumar V,Mugesh S,Durgapal M,et al.Simultaneous production of 3-hydroxypropionic acid and 1,3-propanediol fromglycerol using resting cell system ofD-lactic acid deficient mutant ofKlebsiella pneumoniaewith over expression of KGSADH [J].Bioresource Technology,2013,135:555-563.
[30] ZHOU S,Catherine C,Rathnasingh C,et al.Production of 3-hydroxypropionic acid from glycerol by recombinantPseudomonas denitrificans[J].Biotechnology and Bioengineering,2013,110(12):3 177 -3 187.
[31] Arasu M V,Sarkar R,Sekar B S,et al.Isolation of a novelPseudomonas speciesSP2 producing vitamin B12 under aerobic condition[J].Biotechnology and Bioprocess Engineering,2013,18(1):43 -51.
[32] Kumar V,Ashok S,Park S.Recent advances in biological production of 3-hydroxypropionic acid[J].Biotechnology Advances,2013,31(6):945 -961.
