鸡传染性支气管炎病毒ck/CH/LHLJ/090908株的基因组特性研究
2014-03-08陈玲凤韩宗玺邵昱昊孔宪刚刘胜旺
陈玲凤,韩宗玺,邵昱昊,孔宪刚,刘胜旺
(中国农业科学院哈尔滨兽医研究所 兽医生物技术国家重点实验室/禽呼吸道病创新团队,黑龙江 哈尔滨 150001)
鸡传染性支气管炎(Infectious bronchitis,IB)是由IB 病毒(IBV)引起的一种急性高度接触性的传染性呼吸道疾病。该病主要侵害鸡的呼吸系统和泌尿生殖系统,造成肉鸡增重减少、蛋鸡产蛋数量和质量下降,给养鸡业带来严重经济损失[1]。
IBV 基因组为单股不分节段的正链RNA,基因组全长约为27 600 nt[2],具有5' 端帽子结构和3' 端poly(A)结构。该病毒基因组含有6 个开放阅读框(ORF),从5'端开始的前2/3 的基因组序列仅编码具有活性的聚合酶,后1/3 的基因组序列编码病毒的4 个结构蛋白[3]。其中mRNA1 包含2 个ORF,即ORF1a 和ORF1b,分别编码450 ku 和300 ku,但因核糖体移码作用,这两个ORF 可合并产生一个大蛋白即PP1ab,分子量约为750 ku。该大蛋白可通过病毒自身编码的蛋白酶裂解为相应的功能蛋白[4]。ORF1a 和ORF1b 之间还存在特殊的发夹结构,称之为假结,研究表明该结构可通过调节核糖体延伸循环而改变重叠区的阅读框[5]。假结上游28 nt 处为滑脱序列,为保守的TTTAAAC[6]。而mRNA2,mRNA4,mRNA6 为单顺反子,分别编码结构蛋白:纤突蛋白(S)、膜蛋白(M)、核衣壳蛋白(N);mRNA3,mRNA5 为多顺反子,各含3 个和2 个ORF,编码3a、3b、E 及5a 和5b 蛋白[7]。
尽管H120 疫苗广泛应用国内大多数鸡场,但H120 样型病毒株在发病鸡场中仍有较高的分离率。本研究通过对病毒株ck/CH/LHLJ/090908 的基因组序列进行遗传进化分析,结果表明该分离株与H120疫苗株亲缘关系最近,但其E 蛋白缺失11 个氨基酸,显示该分离株为H120 疫苗株在鸡群中自然传代中形成的变异株。
1 材料和方法
1.1 病 毒 IBV ck/CH/LHLJ/090908 株为本实验室分离鉴定[8]。
1.2 主要实验材料 TRIzol Reagent RNA 提取试剂盒购自Invitrogen 公司;One Step RT-PCR Kit Ver.2、One Step PrimeScript®RT-PCR kit、3'-Full RACE Core Set Ver.2、Clontech SMARTerTMRACE cDNA Amplification Kit、DNA Maker、Ex Taq DNA 聚 合酶、pMD18-T 载体均购自宝生物工程(大连)有限公司;DNA 纯化回收试剂盒购自OMEGA 公司;受体菌TG1 由本实验室制备并保存;引物由本实验室设计并保存。
1.3 病毒基因组RT-PCR扩增及克隆测序 以含病毒鸡胚尿囊液中提取的总RNA 为模板,进行RT-PCR。反应程序为:50 ℃30 min,94 ℃2 min;94 ℃30 s、55 ℃30 s、72 ℃2 min,30 个循环;72 ℃10 min。基因组5' 端和3' 端的序列分别参照Clontech SMARTerTMRACE cDNA Amplification Kit和3'-Full RACE Core Set Ver.2 的使用说明书进行扩增。将扩增的片段分别克隆于pMD18-T 载体中,由北京六合华大基因科技股份有限公司进行测序。
1.4 ORF的确定和序列分析 应用DNAStar 中MEGALIGN 程序对病毒的全基因组序列进行拼接和比对,经Gene Runner3.0 软件对序列进行编辑和ORF 确定,并以MEGA6.0 软件对该病毒株的全基因组序列进行遗传进化分析。
2 结果
2.1 ck/CH/LHLJ/090908的全基因组序列扩增、克隆及测序 从ck/CH/LHLJ/090908 病毒RNA 中扩增出16 条与预计大小相符的片段(图1),将扩增片段分别克隆于pMD18-T 载体中,筛选重组阳性质粒,每个片段选择5 个以上独立克隆进行测序,保证病毒株序列测定的准确性。
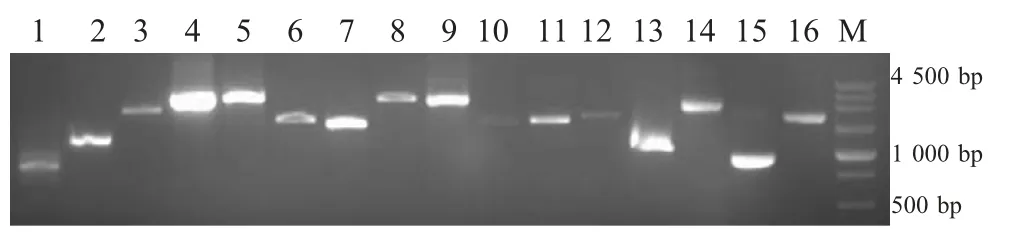
图1 IBV 全基因组的RT-PCR 扩增的16 个重叠片段电泳图Fig.1 RT-PCR amplifications of the 16 overlapping fragments covering the whole genome of IBV
2.2 ck/CH/LHLJ/090908的全基因组序列的确定将病毒基因组中获得的16 个首尾重叠的基因片段进行拼接,与参考病毒株Beaudette 进行比较,显示该分离株基因组的组成特征为:5'-1a-1b-S-3a-3b-EM-5a-5b-N-3',符合IBV 基因组组成特征(表1)。测定其基因组全长为27 609 nt,与H120 疫苗株具有较高的核苷酸同源性(GenBank No.KJ425503)。
2.3 ck/CH/LHLJ/090908全基因组序列分析 将该分离株与20 株参考株进行比较分析,核苷酸同源性在91.3 %~99.8 %。通过构建遗传进化树(图2),结果表明该病毒株为H120 样型病毒株,与H120 同源性极高,达99.7 %。
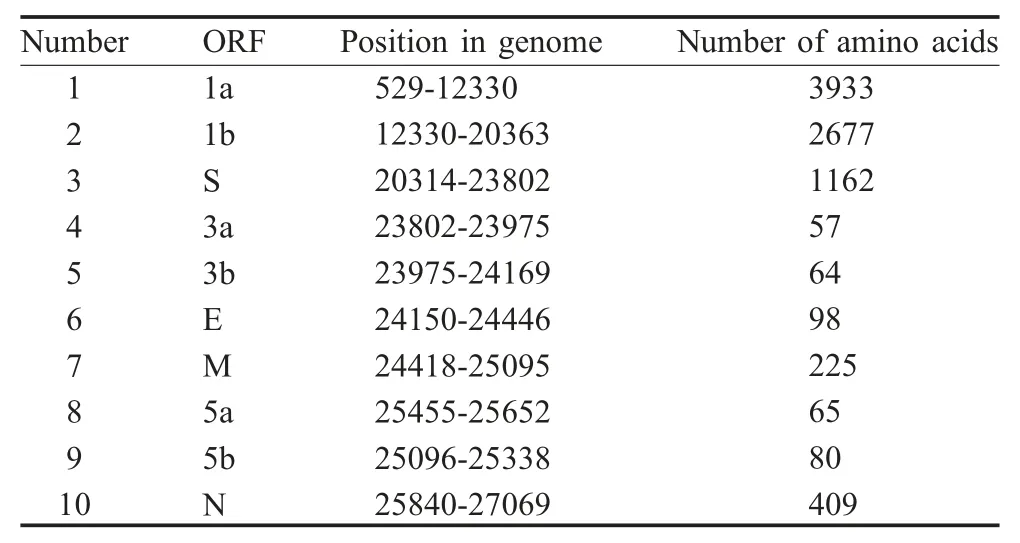
表1 ck/CH/LHLJ/090908 推测的ORFTable 1 ORF encoded in ck/CH/LHLJ/090908 genome
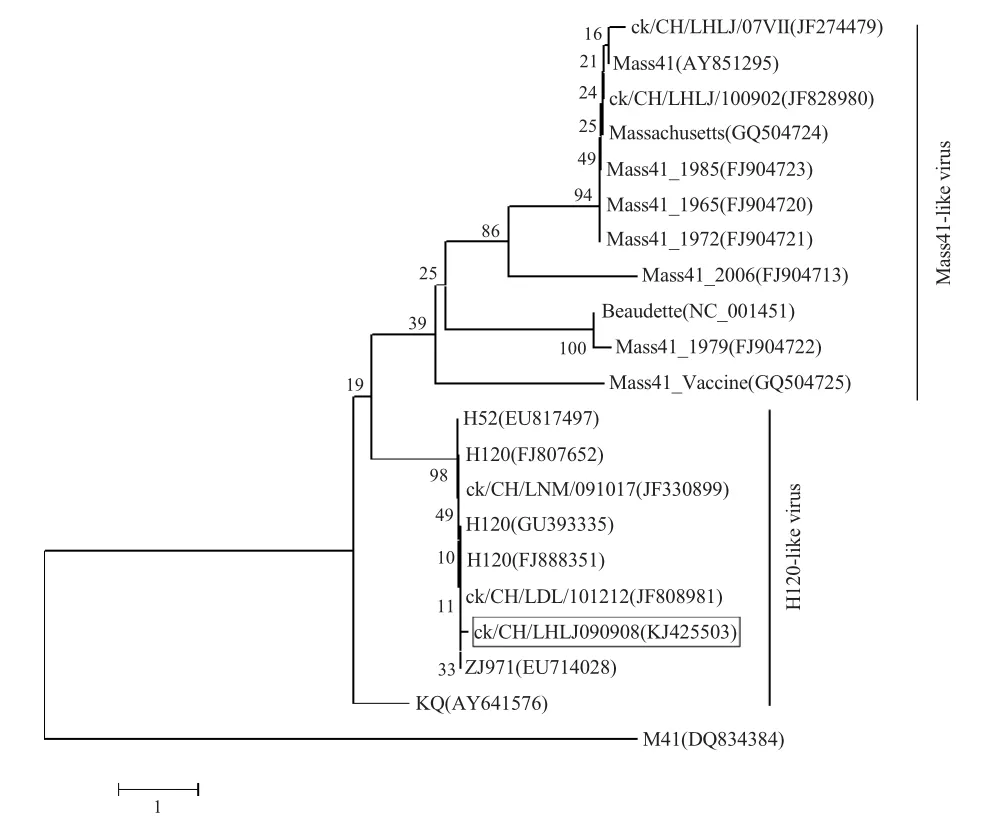
图2 IBV 全基因组核苷酸序列系统进化分析Fig.2 Phylogenetic tree constructed on the basis of the full-length genome of IBV
2.4 ck/CH/LHLJ/090908的ORF1遗传特征 分离株ck/CH/LHLJ/090908 的ORF1 全长19 835 nt,包含两个ORF,即ORF1a 和ORF1b。其中ORF1a 为11 802 nt,编码3 933 个氨基酸残基组成的多肽;ORF1b 为8 034 nt,编码2 677 个氨基酸残基组成的多肽。ck/CH/LHLJ/090908 与H120 的ORF1 核苷酸序列同源性较高,达99.9 %。基因组仅存在少量的点突变。
2.5 ck/CH/LHLJ/090908的S基因遗传特征 分离株ck/CH/LHLJ/090908 与H120 疫苗株的S 基因的转录相关序列(TAS)及其ORF 起始密码子ATG 之间的核苷酸数目相同,均为52 nt,而且TAS 序列也相同,为CTGAACAA。其S 基因为3 489 nt,编码1 162个氨基酸组成的S 蛋白。其中S1 基因为1 611 nt,编码537 个氨基酸组成的S1 蛋白,而S2 基因为1 878 nt,编码625 个氨基酸组成的S2 蛋白。其S蛋白的裂解识别位点为533RRFRR537,与H120 疫苗株相同。
分离株ck/CH/LHLJ/090908 的S 基因与H120 比较,核苷酸同源性为99.7 %。其S1 基因与H120 比较,同源性为99.8 %,仅出现4 个位点的突变,突变位点分别位于351、439、440、1 063 位;S2 基因与H120 比较,同源性为99.7 %,出现6 个位点的突变,突变位点分别位于174、1 046、1 290、1 529、1 722、1 796 位。
2.6 ck/CH/LHLJ/090908的ORF3遗传特征 IBV分离株基因3 的TAS 序列为CTGAACAA,与H120相同。其TAS 序列与ORF 起始密码子ATG 之间的核苷酸数目相同,均为23 nt。IBV 分离株ck/CH/LHLJ/090908 的基因3 由645 nt 组成,而H120 的基因3 由678 nt 组成。进一步比较分析发现ck/CH/LHLJ/090908 的ORF3c 基因在248 nt~280 nt 处缺失33 个碱基(图3),导致E 蛋白缺失11 个氨基酸,编码98 个氨基酸的E 蛋白,而H120 疫苗株由330 nt组成,编码109 个氨基酸的E 蛋白。
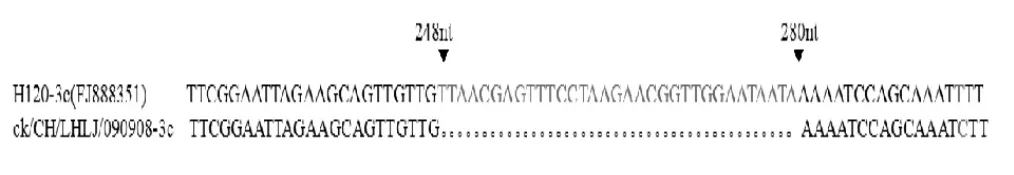
图3 ck/CH/LHLJ/090908 ORF3c 基因33 个碱基的缺失Fig.3 The deletion of 33 nucleotides in the ORF3c gene of ck/CH/LHLJ/090908
2.7 ck/CH/LHLJ/090908的M 基因遗传特征 IBV分离株的M 基因核苷酸序列由678 nt 组成,编码225 个氨基酸的M 蛋白。与H120 进行比较分析,其核苷酸同源性为99.9 %,变异仅发生在T189C 位。M 蛋白含有一个或两个潜在的糖基化位点,研究发现,M 蛋白至少有一个共同的潜在糖基化位点,即位于高度保守的NCT 序列上[9]。比较分析IBV 分离株M 蛋白的氨基酸序列,有两个潜在的糖基化位点,为NET 和NCT。
2.8 ck/CH/LHLJ/090908的ORF5遗传特征 分离株的基因5 由443 nt 组成,ORF5a 由198 nt 组成,编码65 个氨基酸的5a 蛋白;ORF5b 由249 nt 组成,编码82 个氨基酸的5b 蛋白。比较分析IBV 分离株与H120 参考株的基因5 序列,发现ORF5a 和ORF5b 基因核苷酸序列同源性均为100 %。
2.9 ck/CH/LHLJ/090908的N基因遗传特征 分离株的N 基因均由1 230 nt 组成,编码409 个氨基酸的N 蛋白。32 株IBV 分离株均含有30 个丝氨酸残基,其中在165~190 位约有9 个丝氨酸比较保守,为丝氨酸聚集区[10]。比较分析IBV 分离株与H120参考株的N 基因序列,其核苷酸同源性为99.8 %,仅存在两个点突变,即T64C 和C536T。
3 讨论
分离株ck/CH/LHLJ/090908 属于H120 样型病毒株,与H120 疫苗株的基因组序列进行比较分析,结果表明其核苷酸同源性达99.7 %,只有0.3 %的核苷酸差异。将其结构蛋白基因与H120 进行比较分析,其中M 蛋白的氨基酸同源性为100 %;S 蛋白及N 蛋白的氨基酸同源性均为99.5 %;而E 蛋白的氨基酸同源性仅为88 %,差异最大。进一步的分析表明该分离株的ORF3c 序列在248 nt~280 nt 处连续缺失33 nt,导致E 蛋白缺失11 个氨基酸。
在国内,H120 疫苗仍是应用最广泛的疫苗。本实验室分离株ck/CH/LHLJ/090908 核苷酸序列与H120 疫苗株高度同源,但其ORF3c 基因缺失了33 nt,导致E 蛋白缺失11 个氨基酸,基因组其他位置仅出现点突变现象。结果表明,该分离株的出现可能与弱毒疫苗的使用有关,由于其E 蛋白缺失11 个氨基酸,表明该病毒株可能是疫苗毒在鸡体中传代进化形成。Wang 等的研究表明基因组的累积突变能引起抗原的变异,从而导致免疫失败的情况出现[11]。疫苗的广泛使用使得疫苗病毒株长期存在于鸡场中,部分鸡群由于免疫力低下,使得病毒在免疫压力下部分氨基酸发生改变以逃避宿主的免疫清除机制,从而造成新病毒株的分离。
随着分子生物学技术的快速发展,开发出免疫保护率高、安全性好、成本低廉并且适于商业化生产的活载体疫苗或亚单位疫苗是未来IB 疫苗研究的趋势[12-13]。本研究了解了IBV 的遗传变异规律,为制定有效的免疫预防措施提供了实验依据。
[1]马会杰,刘孝珍,韩宗玺,等.鸡传染性支气管炎病毒ck/CH/LHB/100801 株的分离鉴定及分子特征研究[J].中国预防兽医学报,2011,33(5):348-353.
[2]Boursnell M E G,Brown T D K,Foulds I J,et al.Completion of the sequence of the genome of the coronavirus avian infectious bronchitis virus[J].J Gen Virol,1987,68(Pt1):57-77.
[3]刘胜旺.我国鸡传染性支气管炎流行现状及原因分析[J].中国家禽,2010,32(16):5-9.
[4]Ziebuhr J,Thiel V,Gorbalenya A E.The autocatalytic release of a putative RNA virus transcription factor from its polyprotein precursor involves two paralogous papain-like proteases that cleave the same peptide bond[J].J Biol Chem,2001,276(35):33220-33232.
[5]Brierley I,Rolley N J,Jenner A J,et al.Mutational analysis of the RNA pseudoknot component of a coronavirus ribosomal frameshifting signal[J].J Mol Biol,1991,220(4):889-902.
[6]刘胜旺,刘玉芬,孔宪刚,等.鸡传染性支气管炎中国地方分离毒株LX4 mRNA1 ORF1 的遗传变异特征[J].病毒学报,2005,20(4):347-352.
[7]Mardani K,Noormohammadi A H,Hooper P,et al.Infectious bronchitis viruses with a novel genomic organization[J].J Virol,2008,82(4):2013-2024.
[8]Sun Chu-yang,Han Zong-xi,Ma Hui-jie,et al.Phylogenetic analysis of infectious bronchitis coronaviruses newly isolated in China,and pathogenicity and evaluation of protection induced by Massachusetts serotype H120 vaccine against QX-like strains[J].Avian Pathol,2011,40(1):43-54.
[9]Cavanagh D,Davis P J.Evolution of avian coronavirus IBV:sequence of the matrix glycoprotein gene and intergenic region of several serotypes[J].J Gen Virol,1988,69:621-629.
[10]Sapats S I,Ashton F,Wright P J,et al.Novel variation in the N protein of avian infectious bronchitis virus[J].Virology,1996,226(2):412-417.
[11]Wang Li,Junker D,Hock L,et al.Evolutionary implications of genetic variations in the S1 gene of infectious bronchitis virus[J].Virus Res,1994,34(3):327-338.
[12]Tomley F M,Mockett A P A,Boursnell M E G,et al.Expression of the infectious bronchitis virus spike protein by recombinant vaccinia virus and induction of neutralizing antibodies in vaccinated mice[J].J Gen Virol,1987,68(9):2291-2298.
[13]Johnson M A,Pooley C,Ignjatovic J,et al.A recombinant fowl adenovirus expressing the S1 gene of infectious bronchitis virus protects against challenge with infectious bronchitis virus[J].Vaccine,2003,21(21-22):2730-2736.
