动物眼睛的起源与进化研究进展
2013-12-21马冬梅朱华平桂建芳
马冬梅, 朱华平, 桂建芳
(1.中国水产科学研究院珠江水产研究所 农业部热带亚热带水产资源利用与养殖重点实验室, 广州 510380; 2.中国科学院水生生物研究所 淡水生态与生物技术国家重点实验室, 武汉430072)
脊椎动物的眼睛是高度特化、结构复杂、功能完善的精密器官,仅视网膜内就至少有7种高度分化的细胞类型,其中神经细胞相互联系,将接收到的光信号转化为化学信号,传入大脑,产生视觉。解释像眼睛这样精细器官的进化,对于进化生物学家是一种挑战[1-2]。在“物种起源”理论中,达尔文提出了一个假说,现在进化较为完善的眼睛是由简单而不完善的原始眼睛进化而成的。达尔文推测原始的眼睛至少由两种细胞组成:视神经(光感受器细胞)和在一侧保护它的色素细胞,并被半透明的表皮所覆盖,但没有晶体和任何起折射作用的组织。涡虫的眼睛就是这种原始的眼睛。比较解剖学家已经发现了大量的介于原始眼睛和脊椎动物眼睛的中间类型,例如:杯状眼(eye cups)、小孔眼(pinhole eye)、具有晶体的照相机似的眼睛(camera-type eye)、反光镜眼(reflecting mirror eye)和由许多小眼组成的复眼(compound eye)等,这为达尔文的理论提供了依据。但是达尔文承认自然选择理论不能解释原始眼睛的起源,因为选择只能在眼睛已经有一定功能,能够感受光线后才能作用于眼睛的进化。选择不能解释原始眼的起源,就好像不能解释生命的起源一样,眼睛和生命的出现都可以称之为奇迹[3-4]。随着科学的发展,人们对眼睛的进化已经有了非常深入的了解,达尔文的解释,在今天看来,也是无可挑剔的。
1 眼睛的出现
在生物进化过程中,光线可能是一种巨大的推动力量,而动物的视觉又是一个非常重要的选择优势。生物在地球上出现后的1015个日出与日落,使生物体学会了利用光来感受这个世界,进化出了眼睛[5]。在寒武纪(Cambrain)以前眼睛是以一个简单的视杯形式而存在,仅用来感受光线,不能加工成直接的信息。虽然现在科学上还无法解释,但已证实在寒武纪的确发生了一次“物种形成”或者说“物种进化”的大爆炸[6],这时随着捕食动物和捕食作用的出现,眼睛的类型发生了根本性的变化,光作为一种动作信号而加速了眼睛的进化。1998年,Parker研究了加拿大Rocky Mountains的Yoho国家公园内的伯吉斯页岩(Burgess Shale)中古老物种(Wiwaxiacorrugata,Canadiaspinosa和Marrellasplendens生存在大约5亿5千万年前的中寒武纪时期)的化石,分析结果表明,它们整齐而粗糙的体表可能在当时起到了衍射光栅的作用。这些小动物身体上具有刺状附肢,附肢上整齐的凹槽在阳光的照射下可以反射光线,并衍射出有金属光泽的彩虹色的亮光,以吓跑捕食者,这可能是后生动物的第一个视觉信号,而这种视觉信号加速了眼睛的进化[7]。目前人们对此十分感兴趣,还在做进一步的研究。
眼睛收集光线,由晶体来聚焦,投影到感光细胞上,感光细胞将光信息转化为化学神经信号。有些眼睛没有瞳孔,有些甚至没有晶体,如鹦鹉螺(Nautilus)的眼睛。但根据定义,特化的具有光转换功能细胞的器官就可能称之为眼。从寒武纪开始,软体动物门(Mollusca)、节肢动物门(Arthropoda)和脊索动物门(Chordata)的动物就形成了具有功能的眼[5]。
2 眼睛进化所需的时间
关于眼睛进化所需要的时间问题也是创造论和进化论争论的焦点之一。1994年,两位瑞典生物学家Nilsson和 Pelger认为,估计眼睛结构的进化所需要的代数比估计其它复杂的器官要简单。为了确定眼睛结构进化到现在所需要的代数,Nilsson设计了一个简单的计算模型,描绘了从眼点(light sensitive spot)到具有晶体的眼睛的进化模式。他们假设眼睛的进化存在无数微小的变异,把每一个微小变异设置为1%的改进(比如长度改变了1%),他们算出从眼点到具有晶体的眼睛共需要1829个1%的改进。同时,这些微小的变异将会对生物的生存有影响,使捕食者更好地发觉猎物或使猎物更好地躲避捕食者。尼尔森和佩尔格将眼睛的遗传率设为50%。代入群体遗传学专门计算一个有优势的遗传变异在生物群体中的传播时间方程式,结果表明从眼点进化到照相机似的眼睛需要363992代,以一代为一年计算,大概364000年,这在地质年代上只是一瞬间,但这一估计并没有把巩膜和光感受器等的进化时间估计在内。但是,这已充分说明眼睛的进化速度,要比以前人们设想的快得多。生物在地球上至少已有6亿年的历史,这足够让眼睛进化出很多次[8]。
3 眼睛的单起源论
一些科学家认为眼睛是单起源(monophyletic)的,即由一个祖先进化来的,而另一些科学家认为眼睛是多起源(polyphyletic)的,即由多个祖先进化来的。1977年,Salvini-Plawen 和Mayr比较了许多物种眼睛的结构、光感受器的类型、眼的发生过程,以及感光神经细胞轴突的位置和其它解剖学特征。根据分析的结果,他们认为眼睛的进化不是单起源的,而是至少有四十种以上的起源方式[9]。
最近基于形态学证据的“多起源论”受到了分子生物学实验结果的挑战。有研究表明,一个非常保守的“决定”基因——Pax6 (pairedhomeobox6)可以在不同的物种中诱导眼睛的形成,说明Pax6 基因对于眼睛的形态发生是一个关键基因,进而证明后生动物眼睛是从一个原始类型进化而来的,这种进化机制叫做居间进化(intercalary evolution)[10-11]。
Pax6 基因首先是在小鼠和人中克隆得到的,它与小鼠的小眼突变(smalleye)[12-13]和人的无虹膜病(Aniridia)有关[14]。杂合的Pax6 基因突变导致人和小鼠的眼睛有缺陷,纯合的Pax6 突变对于小鼠来说是致死的,突变小鼠没有眼睛和鼻子,脑也发育不全[12-14]。在果蝇中发现了一个Pax6 基因同源物eyeless基因,这两个基因序列十分相似,paired domain的同源性达到94%,homeodomain的同源性达到90%。小鼠的Pax6 基因在果蝇中的异位表达诱导出了异位的复眼,说明Pax6 基因在调节眼睛形态建成中有通用的功能[11]。这使人们改变了一直以来认为昆虫的复眼与脊椎动物的照相机似的眼睛是两个不同起源的观点。
4 眼睛的类型与进化
自然界中动物的眼睛被分为两个最基本的类型,即单眼(single eye)和复眼(compound eye)。单眼主要有三种,一种是照相机似的眼睛(正透镜眼)(positive lens or camera eye),存在于脊椎动物和一些非脊椎动物,例如人和头足类的眼睛。第二种是凹透镜眼(concave mirror eye),蛤和介形亚纲甲壳类的眼,能产生明亮但却十分模糊的图案。第三种是小孔眼(pinhole eye),小孔眼和杯状眼主要存在于软体动物中,它只能观察到物体的位置。复眼由若干个小眼(ommatidia)组成,分为两种类型,一种是联立眼(apposition eye),小眼具有独立的功能,另一种是重叠象眼(superposition eye),小眼一起合作在视网膜上产生明亮而有层理的图像[15]。
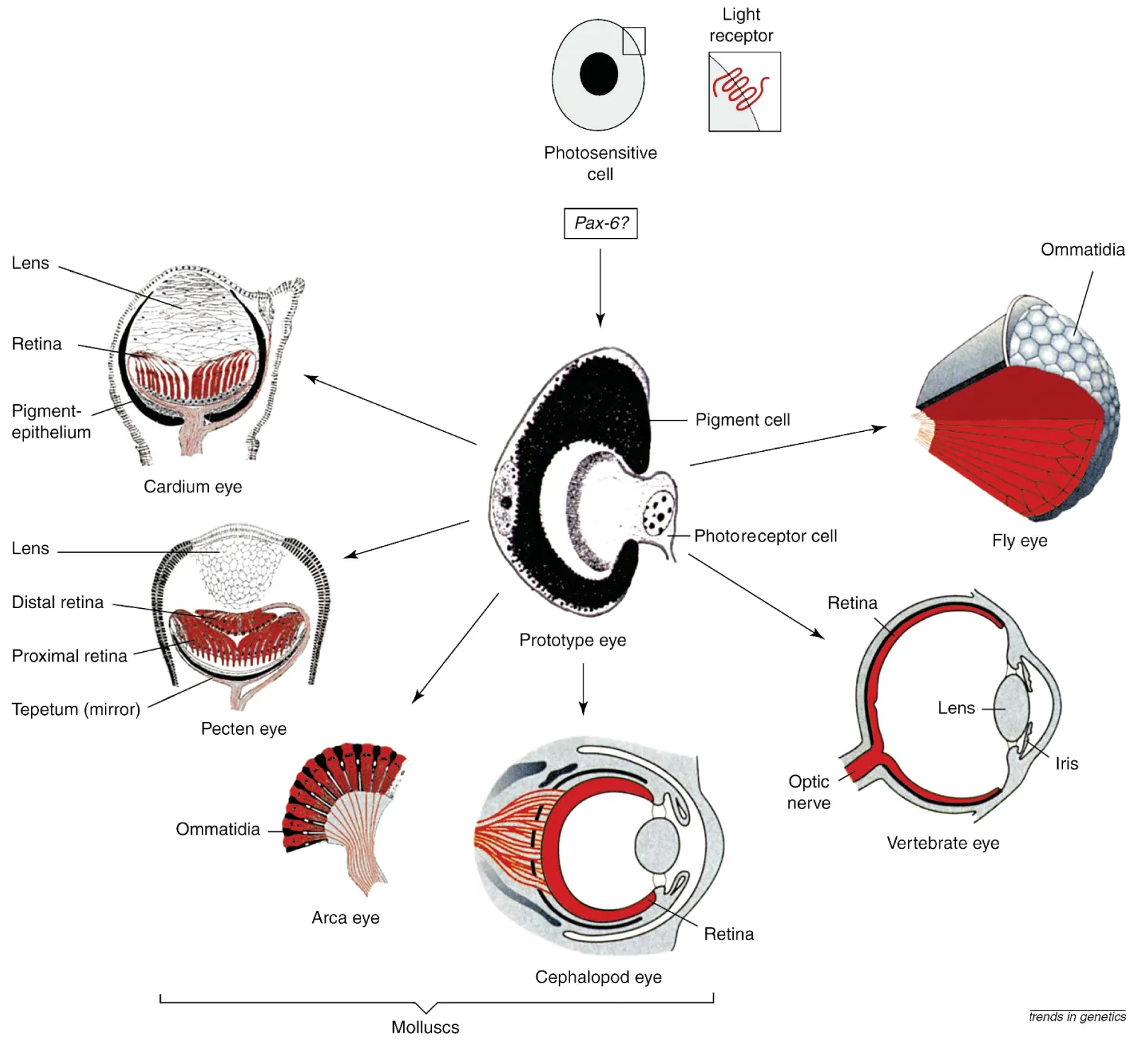
图1 从一个原始类型进化为各种不同类型眼睛的示意图(引自参考文献[10])
如图1所示,根据Gehring和Ikeo的假设,眼睛起源于一种古老的原始类型,原始眼。首先,具有视蛋白(有光感受功能的蛋白)的光敏感细胞进化出现,它在Pax6基因的作用下与色素细胞组成一个简单的器官——原始眼。然后,原始眼通过平行进化和趋同进化,进化出各种不同类型的眼睛:有昆虫的复眼;脊椎动物的照相机似的眼睛;软体动物蛤类(Cardium)的原始照相机似的眼;扇贝(Pecten)的反射镜透镜眼;舟贝(Arca)的复眼和高度进化的头足类的照相机似的眼睛[10]。
5 光感受器与眼睛进化的关系
根据Fernald假设,眼睛的起源是开始于一种对光敏感的视蛋白或视网膜分子[15]。如图2所示,它在Pax6或eya的作用下首先进化成两种光感受器,一种是睫纤毛光感受器(Ciliary photoreceptors);另一种是微绒毛光感受器(Microvillar phototreceptors)[15-16]。
睫纤毛光感受器和微绒毛光感受器再各自进化成一种照相机似的单眼和一种复眼。睫纤毛光感受器进化成的照相机似的单眼,是在Rx、Sox2、PITX2.3、FOXC1、OTX1、MAF和PAX2等基因的作用下完成的,脊椎动物的眼睛就是这种眼睛。睫纤毛光感受器另外进化成一种复眼,如名叫 arc clams的软体动物就具有这种复眼。从微绒毛光感受器也进化出了照相机似的单眼,如头足类的眼睛就是这种单眼。而微绒毛光感受器在dac、toy、so、optix、ey、eyg、ato、notch、dpp、Egfr和hh等基因的作用下进化成了昆虫所具有的复眼[16-17]。
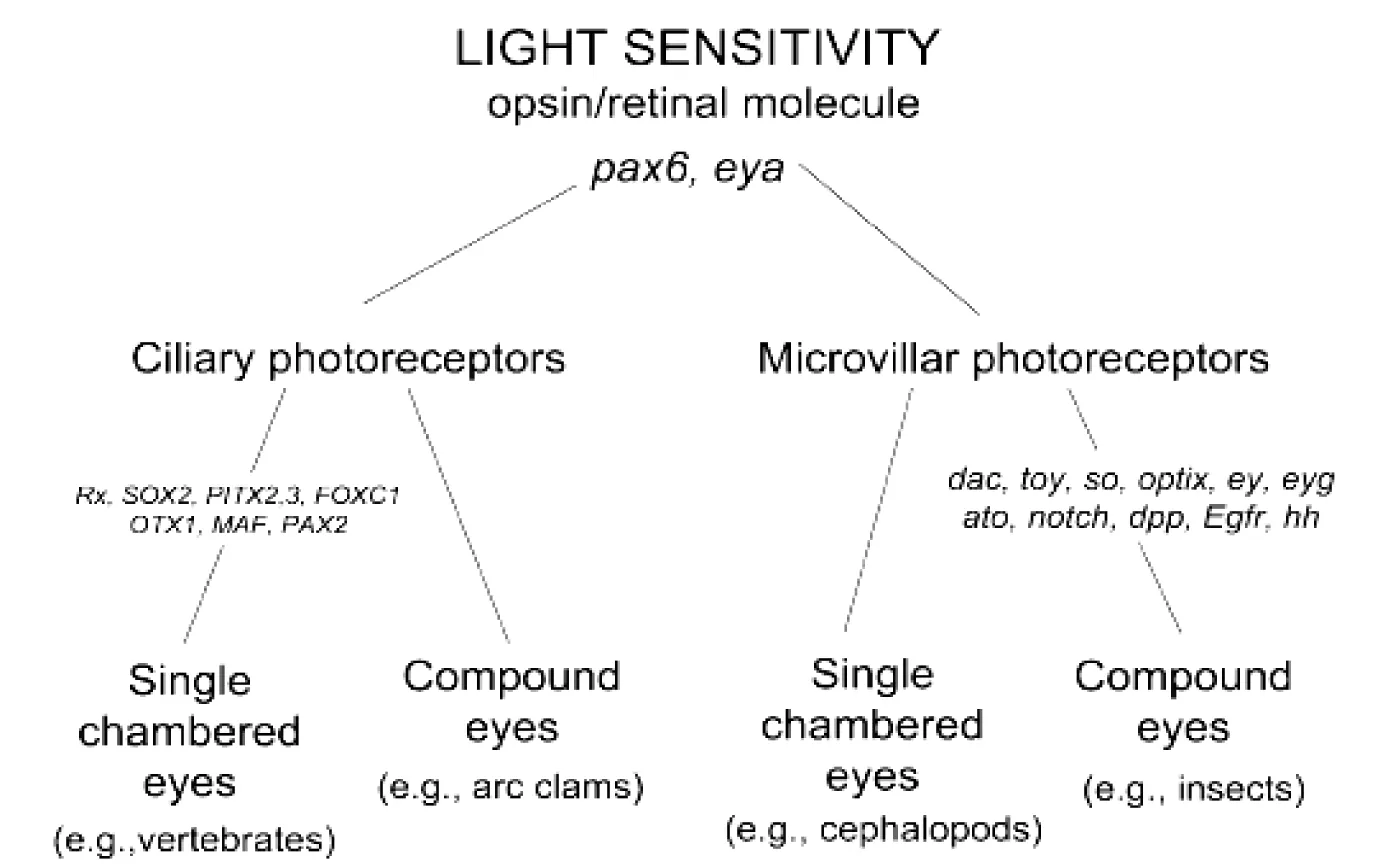
图2 眼睛可能的进化方式示意图(引自参考文献[16])
6 光感受器和视蛋白的类型与进化
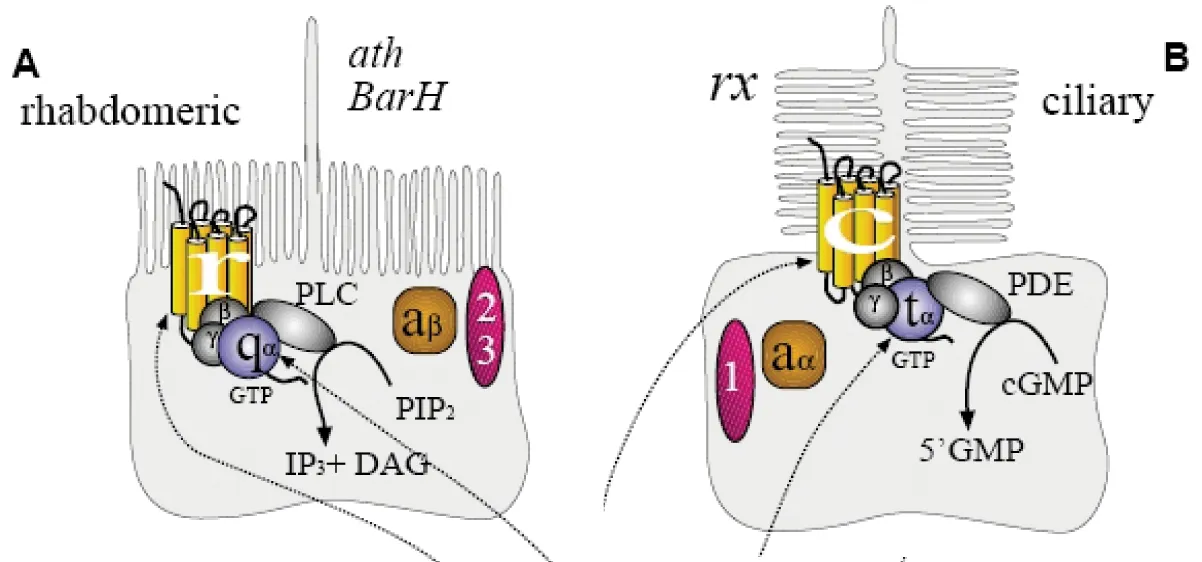
图3 微绒毛光感受器细胞和睫纤毛光感受器
如上所述,光感受器细胞分为两个基本的类型:一种具有微绒毛(microvilli),另一种具有睫纤毛(cilia),它们都有很大的膜表面积以容纳光转换器分子——视蛋白。微绒毛光感受器也叫弹状细胞(rhabdomeric)光感受器,主要存在于无脊椎动物中,而所有脊椎动物的光感受器都是睫纤毛光感受器也叫睫状细胞(ciliary)光感受器。这两种光感受器的形态有区别,微绒毛光感受器上表面垂直折叠以增加它的表面积(图3A),而睫纤毛光感受器增加表面积则通过对睫纤毛的膜水平折叠,使它看起来象一叠盘子(图3B)[18]。它们的生化反应过程也是不同的,节肢动物和软体动物的微绒毛受体在光的刺激下极化,吸收光子,打开Na+离子通道。而脊椎动物的睫纤毛光感受器在光的刺激下关闭Na+离子通道而超极化。脊椎动物和无脊椎动物的光信号转换机制也是不同的。脊椎动物光感受器利用环GMP(cyclic GMP)作为二级传导系统,无脊椎动物则是利用三磷酸肌醇(inositol trisphosphate)[5]。视锥细胞和视杆细胞是由古老的睫纤毛光感受器原体进化而来的,视神经节细胞、无长突细胞和极细胞是从微绒毛光感受器原体进化而来的[18]。
现在所有的眼睛感光都是利用以维生素A为基础的视色素,它是由一个发色团和一个脱辅基蛋白——视蛋白组成。视蛋白存在的历史比眼睛存在的历史还要长。遗传分析表明,视蛋白都是同源的,是从相同的祖先进化而来[19]。在硬骨鱼类、爬行类和鸟类的研究中发现,古老的光吸收蛋白分子在3亿5千万到4亿年前就进化成4个锥体家族,以分辨色彩,后来杆状视蛋白家族出现,辅助暗视觉[20]。但是在哺乳类中,杆状视蛋白出现的较早,而以3种视色素分子为基础的能区别色彩的视觉是在约3千5百万年前才通过基因重复而获得[5,21],进化到现在,使人类有能力感受到这个绚丽多彩的世界,享受五彩缤纷的世界给我们带来的视觉震撼。
以上的研究结果,简要阐述了目前科学对眼睛的进化的认识。眼睛这个最为复杂、精密、神奇的器官的进化过程还有许多不解之迷,进一步研究眼睛基因发育的特征,破解“相似的眼睛发育程序是如何将单起源的原始眼诱导成如此多样的眼睛种类?”,对于我们来说还是一个极大的挑战。
[1]楼允东, 郑德崇.组织胚胎学 [M].北京: 农业出版社, 1981.
[2]Arendt D, Hausen H, Purschke G.The “division of labour” model of eye evolution[J].Phil Trans R Soc B, 2009, 364: 2809-2817.
[3]Darwin C R.The origin of species by means of natural selection, or the preservation of favoured races in the struggle for life [M].6th ed.London: John Murray, 1872.
[4]Gehring W J.Chance and necessity in eye evolution[J].Genome Biol Evol, 2011, 3: 1053-1066.
[5]Conway-morris S.The crucible of creation [M].Oxford: Oxford University Press, 1998.
[6]Parker A R.Colour in burgess shale animals and the effect of light on evolution in the Cambrian [J].Proc R Soc Lond B Biol Sci, 1998, 265: 967-972.
[7]Fernald R D.Evolution of eyes [J].Current Opinion in Neurobiology, 2000, 10: 444-450.
[8]Nillson D E.A pessemistic estimate of the time required for an eye to evolve [J].Proc Biol Sci, 1994, 256: 53-58.
[9]Salvini-plawen S V, Mayr E.On the evolution of photoreceptors and eyes [J].Evolutionary Biology, 1977, 10:207-263.
[10]Gehring W J, Ikeo K.Pax 6: mastering eye morphogenesis and eye evolution [J].Trends Genet, 1999, 15:371-377.
[11]Halder G, Callaerts P, Gehring W J.Induction of ectopic eyes by targeted expression of the eyeless gene inDrosophila[J].Science, 1995, 267:1788-1792.
[12]Hogan B L, Horsburgh G, Cohen J, et al.Small eyes (Sey): a homozygous lethal mutation on chromosome 2 which affects the differentiation of both lens and nasal placodes in the mouse [J].J Embryol Exp Morphol, 1986, 97: 95-110.
[13]Hill R E, Favor J, Hogan B L, et al.Mouse small eye results from mutations in a paired-like homeobox-containing gene [J].Nature, 1991, 354: 522-525.
[14]Glaser T D, Walton S, Cai J, et al.Pax6 gene mutations in aniridia.In Molecular Genetics of Ocular Diseases [M].Edited by Wiggs JL.New York: J Wiley Inc, 1995, 51-82.
[15]Fernald R D.Eyes: variety, development and evolution [J].Brain Behav Evol, 2004, 64:141-147.
[16]Land M F, Nilsson D E.Animal eyes [M].Ox-ford UK: Oxford University Press, 2002.
[17]Fernald R D.Evolving eyes [J].Int J Dev Biol, 2004, 48: 701-705.
[18]Arendt D.Evolution of eyes and photoreceptor cell types [J].Int J Dev Biol, 2003, 47: 563-571.
[19]Cashmore A R, Jarillo J A, Wu Y J, et al.Cryptochromes: blue light receptors for plants and animals [J].Science, 1999, 284: 760-765.
[20]Bowmaker J K.Evolution of colour vision in vertebrates [J].Eye, 1998, 12: 541-547.
[21]Nathans J.The evolution and physiology of human color vision: insights from molecular genetic studies of visual pigments [J].Neuron, 1999, 24: 299-312.
