临产后子宫体部平滑肌高表达HSP27和 αB-crystallin*
2013-11-07周昌菊张卫社
马 薇, 周昌菊, 张卫社
分娩发动的特征性表现是子宫平滑肌的规律收缩和宫颈口的进行性扩张[1]。这一特征目前公认是由多因素作用、多途径调节、多分子参与、多阶段变化的交互作用过程。交互作用的最终靶器官为子宫平滑肌。在宫缩诱导因素的刺激下,子宫平滑肌产生协调、规律、有力的收缩,引起宫腔内压力升高,启动分娩。研究临产前后人子宫体部平滑肌蛋白质表达的变化,既能反映分娩启动后基因组被激活时蛋白的表达情况,又能反映子宫平滑肌从静息状态到收缩状态的分子转变结果,对于探讨分娩启动后子宫平滑肌的收缩机制和早产的诊治具有重要意义。
本研究拟以人子宫体部平滑肌为研究材料,应用比较蛋白质组学技术筛选出临产前后子宫体部平滑肌热休克蛋白27(heat-shock protein 27,HSP27)和α-晶体蛋白 B 链(α-crystallin B chain,αB-crystallin)的表达变化,探讨小热休克蛋白(small HSPs)与临产后子宫平滑肌的收缩关系及可能机制。
材料和方法
1 标本来源及分组
本研究通过医学伦理委员会认证、经受试对象书面同意,选择2005年11月至2006年11月在湘雅三医院及湘雅医院住院分娩初产妇12例,年龄22~34岁,平均年龄(28.0±5.7)岁。单胎,无妊娠合并症,孕龄38~41周,因胎位异常或胎儿窘迫在硬膜外麻醉下行剖宫产术。分为:(1)未临产(not-in-labor,NIL)组:胎儿电子监护仪示无宫缩,阴道检查宫口未开;(2)自然临产(in-labor,IL)组:自然临产、规律宫缩,宫口扩张≥5 cm。每组6例,所有经催产素或/和米索前列醇引产者一律排除。子宫体部组织取材于宫底前壁下1 cm,18号肾活检穿刺针抽取10 mm厚组织,局部注射缩宫素20 U,压迫片刻,如出血以3-0可吸收线8字缝合止血。标本经冷生理盐水漂洗,液氮冻存备用。
2 主要试剂
尿素、二硫苏糖醇(DTT)、硫脲、碘乙酰胺、固相pH梯度干胶条和考马斯亮蓝G250均为瑞典Amersham Biosciences产品;琼脂糖、过硫酸铵、丙烯酰胺、甲叉双丙烯酰胺、甘氨酸、硫代硫酸钠、碳酸氢铵、四甲基乙二胺、三羟甲基氨基甲烷(Tris)、十二烷基硫酸钠(SDS)和基质α-氰基-4-羟基肉桂酸(CCA)均为Sigma-Aldrich产品;Trizol试剂、Reverse Transcription Kit和 PCR Kit为 Promega产品;DEPC为 Sigma产品;HSP27羊抗人单克隆抗体和β-actin羊抗人单克隆抗体购自Santa Cruz;硝酸纤维素膜为Schleicher&Schuell产品;化学发光试剂为Pierce产品。RNeasy Mini试剂盒和OneStep RT-PCR试剂盒均为Qiagen产品;其它生化试剂均为进口分装或国产分析纯。所用引物由上海生工生物工程公司根据设计合成,见表1。
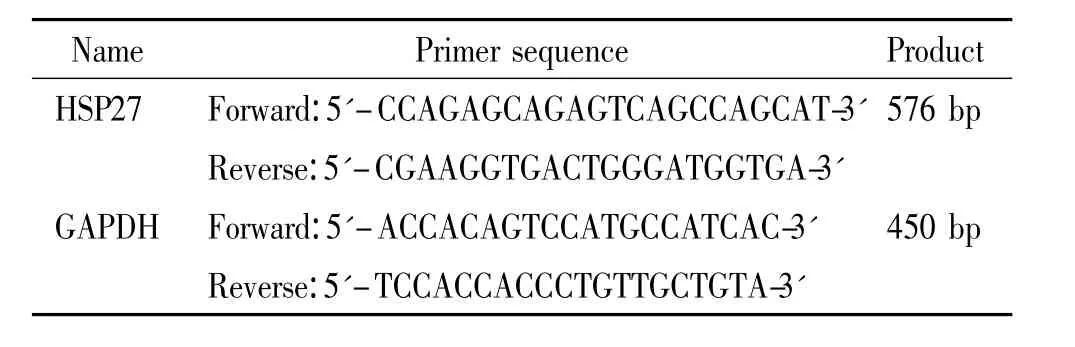
表1 引物序列Table 1.Sequences of the primers
3 生物信息学软件
Lab Scan扫描软件和Data Explorer质谱分析软件为Applied Biosystem产品;ImageMaster 2D Platinum双向凝胶图谱分析软件为Amersham Biosciences产品和Mascot Distiller质谱信号峰识别软件、Mascot肽质量指纹图数据库查询软件和Mascot MS/MS数据库查询软件均为Matrixscience产品。
4 主要方法
4.1 组织总蛋白质的提取 80 mg组织样品加入800 μL组织裂解液的比例加入组织裂解液(7 mol/L尿素、2 mol/L 硫脲、100 mmol/L DTT、4%CHAPS、0.5 mmol/L EDTA、40 mmol/L Tris、2% 两性载体蛋白质pH 3~10)混合,液氮充分研磨。室温放置1 h。离心,取上清液即得蛋白质样品。Bradford法[2]测定蛋白浓度后分装,-80℃冻存。
4.2 双向凝胶电泳(two-dimensional gel electrophoresis,2-DE) 第一向固相pH梯度等电聚焦:取900 μg组织总蛋白质(3个同组样本混合,每个样本300 μg)与水化液(8 mol/L 尿素、4%CHAPS、40 mmol/L Tris、40 mmol/L DTT、0.5%IPG buffer pH 3 ~10)充分混合,使上样总体积为450 μL,采用 pH 3~10,4 cm线性IPG预制胶条,水化和聚焦在20℃自动进行,30 V 水化 13 h后,依次经 100 V 0.5 h、500 V 1 h、1 000V 1 h、5 000 V 1 h、8 000 V 8.5 h 进行等电聚焦,总电压时间积为74 940 Vh。聚焦后先置于平衡 I液(50 mmol/L Tris-HCl、6 mol/L 尿素、30% 甘油、1%SDS、0.2%DTT)中,摇床上平衡 15 min。再在平衡II液(3%碘乙酰胺替代0.2%DTT,其余组分同平衡I液)中平衡15 min。然后进行第二向垂直SDS-PAGE电泳,分离胶浓度为12.5%。
4.3 考马斯亮蓝染色 双蒸水漂洗后,考马斯亮蓝G250染色,双蒸水漂洗至背景清楚。
4.4 凝胶图像分析 凝胶经Image Scanner扫描仪及LabScan扫描软件扫描,ImageMaster 2D Platinum图像分析软件依次进行点检测、背景消减、匹配、量化比较从而获取斑点的相关信息。差异点标准:以NIL组2-DE图谱为参考胶,与IL组子宫体部平滑肌组织的平均胶进行两两匹配比较。以蛋白质表达水平(相对体积)增加2倍及2倍以上为差异有统计学意义。
4.5 基质辅助激光解吸电离 -飞行时间质谱(MALDI-TOF-MS) 分析切割差异蛋白质点于1.5 mL EP管中,50%乙腈和50 mmol/L碳酸氢铵脱色,乙腈脱水,真空冷冻离心抽干,胰蛋白酶(0.1 mg/L)酶解,萃取,真空冷冻离心浓缩,然后与基质液CCA混合,点样于不锈钢板。最后经MALDI-TOF-MS质谱仪质谱分析,获得肽质量指纹图谱(peptide mass finger printing,PMF)。经 Mascot查询系统,检索SWISS-PROT和NCBI蛋白质数据库。搜索结果如果Mascot分数>64分(P<0.05)则认为得到阳性鉴定,否则视为未得到鉴定。
4.6 半定量RT-PCR检测 采用常规Trizol试剂提取临产和未临产子宫体部平滑肌组织总RNA。逆转录反应具体操作如下:总RNA与适量DEPC水和Oligo(dT)混合使其体积达12 μL,轻轻混匀70℃孵育5 min,立即置于冰上冷却5 min,然后配制以下混合物(体积为7 μL):10 ×反应缓冲液 4 μL,RNA 抑制酶 1 μL 和10 mmol/L dNTPs 2 μL;将7 μL 混合物加入到12 μL总RNA混合物中,37℃孵育5 min;最后加入1 μL逆转录酶,使其终体积达20 μL。42℃孵育60 min,70℃孵育10 min。PCR反应体系为20 μL,以GAPDH作为内参照,设置PCR反应条件为95℃ 2 min,94 ℃ 30 s,58 ℃ 30 s,72 ℃ 40 s,共35 个循环。1.5%琼脂糖分离 PCR产物,凝胶成像仪(Dolphin DOC,Wealtec)分析图像。
4.7 免疫印迹验证 取出冻存子宫体部平滑肌组织,以 1∶6 加入组织裂解液(1 mmol/L Tris-HCl、0.5 mmol/L EDTA、2%SDS、5 mmol/L DTT、10 mmol/L PMSF),冰内匀浆 10 min,静置1 h,30 s超声粉碎,4℃、12 000×g离心40 min,收集上清液,Bradford法测蛋白浓度。50 μg蛋白再经浓缩胶、分离胶SDSPAGE电泳分离,电转移至硝酸纤维素膜,5%脱脂奶粉封闭非特异性蛋白结合位点,然后膜与相应HSP27抗体4℃孵育12 h,洗涤;Ⅱ抗室温孵育1 h,洗涤,加入化学发光底物,暗室X光片曝光、显影、定影显示条带。
4.8 免疫组织化学验证 新鲜标本迅速用4%多聚甲醛固定24 h,组织脱水、包埋,切片机切4 μm的石蜡切片,50℃烤片30 min,经脱蜡处理和胰酶修复后加Ⅰ抗,分别滴加1∶100稀释的HSP27抗体,37℃ 1 h,PBS洗3次,每次3 min。加生物素标记Ⅱ抗,37℃ 1 h,PBS洗3次,每次3 min。滴加SABC:20~37℃,20 min,PBS洗4次,每次5 min。DAB显色:1 mL蒸馏水中加DAB显色液A、B、C各一滴,充分混匀、显色、复染、照相。由2个观察者对切片进行盲式阅片,显微镜下随机选取5个视野,Motic显微图像摄像系统摄像,北航MIAS生物医学图像分析管理系统测定产物的吸光度,以此反映阳性产物的相对表达量。
5 统计学处理
用SPSS 16.0统计学软件进行分析。数据采用均数±标准差(mean±SD)表示。2-DE图谱采用ImageMaster 2D Platinum软件进行分析,两两匹配比较,以蛋白质表达水平(相对体积)增加≥2倍为差异有统计学意义。
结 果
1 比较蛋白质组学2-DE图谱鉴定结果
通过2-DE获得NIL和IL组的子宫体部平滑肌2-DE图谱,经蛋白质斑点脱色、还原、烷基化、胰酶酶解、萃取、脱盐后,MALDI-TOF-MS分析,以基质峰和酶自动降解片段峰进行校正,去除角蛋白和胰酶自切峰,精确标定强度为基质峰强度2倍以上的峰,HSP27和αB-crystallin均获得PMF谱,将PMF谱数据输入Mascot查询系统,检索SWISS-PROT和NCBI蛋白质数据库。图1为临产后子宫体部平滑肌2-DE图谱(右侧)和局部放大的差异蛋白HSP27(Spot B)和αB-crystallin(Spot E),2个蛋白在临产后子宫体部平滑肌表达上调(P<0.05)(左侧)。同时发现:临产前后子宫平滑肌2-DE图谱均存在4个分子量相等、等电点不同的HSP27蛋白斑点(Spot A、Spot B、Spot C和Spot D),但2组经ImageMaster 2D Platinum软件分析比较,仅Spot B具有显著差异(P<0.05),其它3个蛋白斑点无显著差异,见表2。图2是Spot B经MALDI-TOF-MS分析得到的PMF谱。
2 HSP27 mRNA的RT-PCR检测结果
以HSP27引物对未临产和临产的子宫体部平滑肌组织进行RT-PCR扩增,HSP27 mRNA在临产后子宫体部平滑肌组织表达明显增高,NIL组和IL组的相对表达量依次为0.4801±0.0453和0.8320±0.0981,差异显著(P <0.01),见图3。

Figure 1.Representative 2-DE map of corpus uteri myometrium in in-labor situation(right).HSP27(Spot B)and αB-crystallin(Spot E)proteins were up-regulated in copus uteri myometrium in in-labor situation(P <0.05).Close-up images of Spot B and Spot E from not-in-labor(NIL,n=6)and in-labor(IL,n=6)2-DE profiles were on the left.Spot A,Spot C and Spot D were also HSP27 identified by MALDI-TOF-MS,but they had no significant difference between IL group and NIL group(P >0.05).图1 临产后子宫体部平滑肌2-DE图谱

表2 MALDI-TOF-MS鉴定的临产前后子宫体部平滑肌蛋白质点Table 2.Protein spots in in-labor(IL)and not-in-labor(NIL)group identified by MALDI-TOF-MS

Figure 2.A representative peptide mass fingerprint of spot B.图2 B点蛋白质的PMF图

Figure 3.RT-PCR for HSP27 mRNA expression in corpus uteri myometrium in in-labor(IL)and not-in-labor(NIL)groups.M:DNA marker.Mean ± SD.n=6.**P <0.01 vs NIL.图3 临产前后子宫体部平滑肌组织HSP27 mRNA表达的比较
3 HSP27蛋白的免疫印迹分析
免疫印迹结果显示,HSP27蛋白在临产后子宫体部平滑肌组织表达明显上调,与蛋白质组学的结果一致。另外,NIL组和IL组表达的HSP27的印迹均为2条,提示HSP27在妊娠晚期和临产后子宫体部平滑肌内的表达可能存在特殊的构象变化,见图4。

Figure 4.Western blotting analysis of HSP27 protein in corpus uteri myometrium in in-labor(IL)and not-in-labor(NIL)groups.β-actin was a loading control.图4 HSP27蛋白免疫印迹分析
4 HSP27蛋白的免疫组化表达
HSP27蛋白阳性表达定位于胞浆。NIL组HSP27阳性表达面积较少,部分胞浆着色;IL组HSP27阳性表达面积较多,胞浆广泛着棕黄色。NIL组和IL组HSP27阳性产物表达的平均吸光度分别为0.2290 ±0.0419 和 0.6360 ±0.1083,差异显著(P<0.01),与蛋白质组学的结果一致,见图5。

Figure 5.Expression of HSP27 in corpus uteri myometrium in in-labor(IL)and not-in-labor(NIL)groups(immunohistochemical staining,×400).Mean ±SD.n=6.**P <0.01 vs NIL.图5 临产前后子宫体部平滑肌HSP27蛋白的表达和吸光度值比较
讨 论
1 临产后子宫体部平滑肌高表达HSP27和αB-crystallin
应用比较蛋白质组学技术,我们成功地鉴定出临产后子宫体部平滑肌高表达HSP27和αB-crystallin两个small HSPs,通过半定量RT-PCR、免疫印迹和免疫组化实验均证实了HSP27蛋白的表达变化与蛋白质组学研究结果高度一致。这与我们之前报道[3]的自然临产后HSP27的基因HSPB1表达上调了4.09倍,催产素诱导的临产子宫体部平滑肌HSPB1上调了4.76倍的结果相一致,提示无论是从基因水平还是蛋白质水平,small HSPs确实参与了临产后子宫体部平滑肌收缩事件。Salintnone等[4]报道small HSPs可以通过分子伴侣变性蛋白、维持细胞间氧化还原状态以及修饰丝状肌动蛋白的聚合作用来调节平滑肌细胞的收缩、迁徙和存活。体外实验[5]也证实热休克蛋白可增加线粒体及收缩蛋白的功能,雌激素在转录及翻译过程中可调节热休克蛋白表达与合成,证实热休克蛋白是一类受雌激素调节、具有增强平滑肌收缩功能的蛋白质。
2 HSP27和αB-crystallin参与子宫平滑肌收缩的可能机制
Bitar等[6]发现HSP27抗体可抑制蛙皮素诱导的小肠平滑肌细胞的收缩作用,最早提出HSP27参与平滑肌收缩的观点。HSP27可以通过改变平滑肌细胞F-actin的稳定性和微丝结构来调节细胞骨架的动力学[7]。人血小板中HSP27能促进F-actin的形成[8]。Bitar等[9]发现兔结肠平滑肌 HSP27 的磷酸化对于平滑肌细胞内部HSP27的结构重组非常必要,且与蛋白激酶C信号转导通路直接相关。兴奋剂诱导HSP27发生磷酸化,而磷酸化的HSP27又通过调节原肌球蛋白(tropomyosin)的细肌丝来调节actin-myosin的交互作用。Somara等[10]报道HSP27可与 actin、tropomyosin、钙调蛋白(calmodulin,CaM)产生免疫共沉淀。磷酸化HSP27调节磷酸化的CaM-tropomyosin的交互作用,结肠平滑肌细胞微丝通过磷酸化结合或解离收缩蛋白产生收缩或舒张效应。HSP27受热休克以及细胞因子、生长因子、激素肽等应激刺激发生磷酸化,而磷酸化的HSP27可以改变actin的细胞骨架以及影响依赖actin的细胞事件。细胞骨架的结构改变是平滑肌收缩的物质基础。
Gerthoffer等[11]指出HSP27的磷酸化作用是actin-myosin发生交互作用诱发平滑肌收缩的先决条件。磷酸化可以诱导small HSPs的三级分子结构发生改变,形成同源/异源寡聚体,从而与actin的细胞骨架发生亲和作用,如磷酸化可以诱导αB-crystallin和HSP27多聚体解离成单聚体及二聚体亚单位。人平滑肌HSP27磷酸化的3个主要位点:Ser-15、Ser-78和Ser-82,与甘氨酸结合可以阻止HSP27寡聚体的形成,抑制 HSP27介导的 F-actin蓄积[12],支持HSP27磷酸化具有稳定actin微丝的作用。MacIntyre等[13]报道临产前HSP27与αB-crystallin主要定位于人子宫下段平滑肌细胞的胞核周围和胞浆,而临产后HSP27则迁移至人子宫平滑肌细胞F-actin束中,且与HSP27-Ser15的磷酸化增加及HSP27-Ser82脱磷酸化同步发生。White等[14]报道妊娠晚期大鼠子宫平滑肌的张力促进HSP27蛋白表达的增加,分娩时磷酸化HSP27(Ser15)由孕鼠子宫平滑肌细胞胞膜位置迁至胞浆,提示HSP27表达增加和移位是其对晚期妊娠至分娩启动的转化过程中子宫张力增加的机械应激反应。Gaestel等[15]认为HSP27的翻译后修饰作用除了磷酸化之外,还可能发生脱酰胺作用或酰化作用、氧化作用以及糖基化作用。本实验中NIL组和IL组子宫体部平滑肌2-DE图谱均发现4个分子量相等、等电点不同的HSP27蛋白点,2组比较仅有1个HSP27蛋白斑点在临产后表达增加。蛋白印迹实验也证实2组子宫体部平滑肌HSP27的印迹表达均为2条,提示HSP27可能通过特定的翻译后修饰作用发生了分子构象变化,协助维持妊娠晚期平滑肌的细胞结构。一旦应激,表达迅速增加,通过与平滑肌细胞收缩装置(actin和/或tropomyosin)发生交联,参与临产后子宫平滑肌细胞的收缩活动。
临产后人子宫体部平滑肌αB-crystallin表达上调。目前αB-crystallin在平滑肌组织中的功能尚不十分清楚,有报道[16]细胞受到压力刺激时,B-crystallin可作为分子伴侣来阻止变性蛋白发生凝集和增加细胞对压力的耐受。亦有证据[17]提示αB-crystallin对肌细胞的中间丝具重要的调节作用。αB-crystallin基因突变可致中间丝断裂凝集、骨骼肌发育障碍和心肌病的发生[18]。Sugiyama 等[19]发现培养的小鼠成肌细胞诱导分化的肌小管中存在αB-crystallin与HSP27的特殊交互作用,二者共同定位于发育良好的肌小管actin束上,但成肌细胞HSP27表达缺失,提示small HSPs参与肌小管中最初的肌原纤维组装。MacIntyre等[13]报道临产前人子宫下段平滑肌中HSP27与αB-crystallin发生交联,临产后该交联作用下降。人子宫平滑肌αB-crystallin和HSP27之间的交联机制尚不明确,可能通过分子伴侣协同作用和/或参与细胞骨架塑形来诱发平滑肌细胞收缩。
3 子宫体部和子宫下段αB-crystallin参与平滑肌收缩的功能可能不同
本研究中临产后子宫体部平滑肌αB-crystallin表达显著上调,而MacIntyre等[13]报道临产后子宫下段平滑肌αB-crystallin表达显著下调,提示αB-crystallin在临产后子宫体部和子宫下段收缩中承担的功能可能不同。由于子宫体部和子宫下段的组织结构、妊娠晚期受到的内分泌环境和机械应激刺激不同,二者在分娩启动中扮演的角色也不同。接受应激信号后,子宫体部表现以功能增加为主,而子宫下段则表现以功能抑制为主。此种反方向的作用机制,为分娩发动中子宫体部平滑肌收缩与子宫下段平滑肌舒张的协调奠定了分子基础[20]。对于临产后αB-crystallin蛋白表达上调而子宫下段其表达下调机制的深入研究有助于我们更进一步理解分娩启动后的子宫平滑肌的收缩机制,改善不良妊娠结局。
Small HSPs作为免疫佐剂和肿瘤治疗的靶点,可以改变细胞或组织的应激能力,增加药物的疗效等都展现了广阔的前景。利用本研究发现的子宫体部平滑肌具有的small HSPs分子靶点及临产时上调的规律,将small HSPs与宫缩剂结合,对于临床治疗梗阻性难产、协调宫缩、增加疗效、减少药物用量等方面提供了新的思路。
(致谢:感谢中南大学湘雅医院卫生部蛋白质组学重点实验室肖志强教授对于本课题的意见!)
[1] Havelock JC,Keller P,Muleba N,et al.Human myome-trial gene expression before and during parturition[J].Biol Reprod,2005,72(3):707-719.
[2] Bradford MM.A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding[J].Anal Chem,1976,72:248-254.
[3] 张卫社,吴新华,伍招娣,等.自然临产及缩宫素诱导临产子宫体部平滑肌差异基因表达谱分析[J].现代妇产科进展,2007,16(3):206-209.
[4] Salinthone S,Tyagi M,Gerthoffer WT.Small heat shock proteins in smooth muscle[J].Pharmacol Ther,2008,119(1):44-54.
[5] Papaconstantinou AD,Goering PL,Umbreit TH,et al.Regulation of uterine hsp90α,hsp72 and HSF-1 transcription in B6C3F1 mice by β-estradiol and bisphenol A:involvement of the estrogen receptor and protein kinase C[J].Toxicol Lett,2003,144(2):257-270.
[6] Bitar KN,Kaminski MS,Hailat N,et al.HSP27 is a mediator of sustained smooth muscle contraction in response to bombesin[J].Biochem Biophys Res Commun,1991,181(3):1192-1200.
[7] Quinlan R.Cytoskeletal competence requires protein chaperones[J].Prog Mol Subcell Biol,2002,28:219-233.
[8] Butt E,Immler D,Meyer HE,et al.Heat shock protein 27 is a substrate of cGMP-dependent protein kinase in intact human platelets.Phosphorylation-induced actin polymerization caused by Hsp27 mutants[J].J Biol Chem,2001,276(10):7108-7113.
[9] Bitar KN.HSP27 phosphorylation and interaction with actin-myosin in smooth muscle contraction[J].Am J Physiol Gastrointest Liver Physiol,2002,282(5):G894-G903.
[10] Somara S,Bitar KN.Phosphorylated HSP27 modulates the association of phosphorylated caldesmon with tropomyosin in colonic smooth muscle[J].Am J Physiol Gastrointest Liver Physiol,2006,291(4):G630-G639.
[11] Gerthoffer WT,Gunst SJ.Invited review:focal adhesion and small heat shock proteins in the regulation of actin remodeling and contractility in smooth muscle[J].J Appl Physiol,2001,91(2):963-972.
[12]Lavoie JN,Gingras-Breton G,Tanguay RM,et al.Induction of Chinese hamster HSP27 gene expression in mouse cells confers resistance to heat shock.HSP27 stabilization of the microfilament organization[J].J Biol Chem,1993,268(5):3420-3429.
[13]MacIntyre DA,Tyson EK,Read M,et al.Contraction in human myometrium is associated with changes in small heat proteins[J].Endocrinology,2008,149(1):245-252.
[14] White BG,MacPhee DJ.Distension of the uterus induced HspB1 expression in rat uterine smooth muscle[J].Am J Physiol Regul Integr Comp Physiol,2011,301(5):R1418-R1426.
[15] Gaestel M. sHsp-phosphorylation:enzymes, signaling pathways and functional implications[J].Prog Mol Subcell Biol,2002,28:151-169.
[16] Chávez Zobel AT,Loranger A,Marceau N,et al.Distinct chaperone mechanisms can delay the formation of aggresomes by the myopathy-causing R120G αB-crystallin mutant[J].Hum Mol Genet,2003,12(13):1609-1620.
[17] Perng MD,Cairns L,van den IJssel P,et al.Intermediate filament interactions can be altered by HSP27 and αB-crystallin[J].J Cell Sci,1999,112(Pt 13):2099-2112.
[18] Vicart P,Caron A,Guicheney P,et al.A missense mutation in the αB-crystallin chaperone gene causes a desminrelated myopathy[J].Nat Genet,1998,20(1):92-95.
[19]Sugiyama Y,Suzuki A,Kishikawa M,et al.Muscle develops a specific form of small heat shock protein complex composed of MKBP/HSPB2 and HSPB3 during myogenic differentiation[J].J Biol Chem,2000,275(2):1095-1104.
[20]张卫社,郭海燕,谢庆生,等.分娩活跃期子宫体部与子宫下段平滑肌差异基因表达谱的筛选[J].国际病理科学与临床杂志,2007,27(5):388-392.
