发酵过程流加L-谷氨酸提高ε-聚赖氨酸的产量*
2013-08-12董难陈旭升任喜东曾昕孙启星张建华毛忠贵
董难,陈旭升,任喜东,曾昕,孙启星,张建华,毛忠贵
(江南大学生物工程学院,工业生物技术教育部重点实验室,江苏 无锡,214122)
ε-聚赖氨酸(ε-poly-L-lysine,ε-PL)具有抑菌谱广、耐热性强、水溶性好和安全性高等特点,被认为是取代现行化学食品防腐剂的理想选择,已在日本、韩国和美国等国家得到广泛应用。同时,它还被广泛用于生物可降解材料、乳化剂、高吸收性水凝胶、药物载体、抗癌增进剂和生物芯片外被等领域的应用研究[1]。因此,ε-PL 是一种附加值高、应用范围广和市场潜力大的新兴生物技术产品。
当前,微生物发酵法是ε-PL 生产的唯一方式。尽管国际上最高ε-PL 发酵水平已经达到48.3 g/L[2],但国内发酵水平普遍处于25 g/L 左右[3-5]。限制国内ε-PL 发酵水平的关键在于产生菌的ε-PL 合成速率低下。大量研究结果表明,筛选ε-PL 合成前体L-赖氨酸结构类似物(S-2-氨基乙基-L-半胱氨酸)抗性突变菌株是提高产生菌ε-PL 合成能力最有效方法之一[6-9]。然而,仅仅解除天门冬氨酸激酶的反馈抑制以增强L-赖氨酸合成能力,而不增加其上游三羧酸循环碳代谢流和相匹配的氮骨架供应能力,依然不能充分发挥ε-PL 高产突变株潜力。目前,还未见基于强化三羧酸循环和氮骨架供应而提出的菌株改造或发酵工艺优化策略的报道。Takehara 等指出L-谷氨酸通过转氨作用为L-赖氨酸合成过程提供氮骨架,同时生成的α-酮戊二酸会进入三羧酸循环[10]。因此,本研究利用1 株ε-PL 高产菌Streptomyces sp.M-Z18 考察外源流加L-谷氨酸对其菌体生长和ε-PL合成的影响。
1 材料与方法
1.1 菌株
Streptomyces sp. M-Z18 由工业生物技术教育部重点实验室人员从土壤中筛选,经紫外线和亚硝基胍诱变后,筛选S-2-氨基乙基-L-半胱氨酸抗性而获得的1 株ε-PL 高产突变株。
1.2 培养方法
种子培养和摇瓶发酵按照文献[11]进行。
分批发酵和补料-分批发酵按照文献[12]进行,并作如下调整:发酵起始pH6.8 下降至3.8 时,用10% ~14%的氨水或2 mol/L H2SO4维持恒定直至发酵结束。添加L-谷氨酸以高浓悬浮液形式加入到发酵罐中。
1.3 分析方法
菌体干重(DCW)、甘油和ε-PL 含量测定均按文献[12]进行。
L-谷氨酸测定:利用SBA-40D 生物传感分析仪(山东省科学院生物研究所)测定。进样量:25 μL。
2. 结果与分析
2.1 摇瓶发酵过程中L-谷氨酸添加浓度对ε-PL 合成和菌体生长的影响
当摇瓶发酵至18 h(ε-PL 开始合成),开始添加L-谷氨酸,使得摇瓶中L-谷氨酸浓度分别控制在0、5、10、15 和20 g/L 水平。96 h 发酵结束后,ε-PL 产量和菌体量如图1 所示。

图1 摇瓶发酵中L-谷氨酸浓度对Streptomyces sp.M-Z18 菌体生长和ε-PL 合成的影响Fig.1 Effect of L-glutamate acid on cell growth and ε-PL production by Streptomyces sp. M-Z18 in shake-flask fermentation
从图1 可以看出,ε-PL 产量和菌体量随L-谷氨酸添加浓度的增加而提高。在添加20 g/L L-谷氨酸时,ε-PL 产量和菌体量分别达到3.82 g/L 和10.84 g/L,较未添加L-谷氨酸发酵提高了82.3% 和58.9%。这说明在发酵过程中添加L-谷氨酸不仅能够促进Streptomyces sp. M-Z18 菌体的生长,还能促进ε-PL 的合成。然而,由于L-谷氨酸在ε-PL 发酵条件(30℃,pH3.8)下溶解度只有10.4 g/L[13],所以在添加20 g/L L-谷氨酸时出现了部分L-谷氨酸未溶解现象。故将下述5 L 发酵罐中L-谷氨酸添加浓度设定在15 g/L。
2.2 发酵罐分批发酵中L-谷氨酸添加时机对ε-PL合成的影响
为了考察L-谷氨酸添加时机对ε-PL 合成过程的影响,分别选择在ε-PL 合成起始阶段(14 h)和高峰阶段(36 h)添加15 g/L L-谷氨酸,实验结果如图2B和图2C 所示。从发酵时间上看,不管是ε-PL 合成起始阶段还是高峰阶段添加L-谷氨酸,均比未添加L-谷氨酸(图2A)的发酵时间缩短5 h,从而使得甘油平均消耗速率提高了10.5%。从ε-PL 产量上看,在ε-PL 合成起始阶段和高峰阶段添加L-谷氨酸的ε-PL产量分别为7.86 g/L 和7.53 g/L,较未添加L-谷氨酸发酵(6.59 g/L)提高19.3% 和14.3%。ε-PL 合成起始阶段添加L-谷氨酸时,ε-PL 的产率为4.01 g/(L·d);高峰阶段添加L-谷氨酸ε-PL 的产率为3.84 g/(L·d),较未添加L-谷氨酸的产率2.97 g/(L·d)分别提高35%和29.3%。另外,前期添加L-谷氨酸和高峰期添加L-谷氨酸的DCW 分别为21.63 g/L 和21.05 g/L,与未添加L-谷氨酸的15.75 g/L 相比,也分别提高37.0%和33.6%。上述实验结果表明,添加L-谷氨酸可以在一定程度上促进碳源的消耗、ε-PL的合成和菌体的生长。但不同时间添加谷氨酸对菌体的生长和ε-PL 的合成影响较小。
从图2B 和图2C 可以看出,不管是ε-PL 合成前期还是高峰期添加L-谷氨酸,15 g/L L-谷氨酸大约在11 h 就会被消耗完。为了保证在分批发酵过程中始终有L-谷氨酸被利用,待ε-PL 合成前期培养基中添加的L-谷氨酸消耗完后,重新添加15 g/L L-谷氨酸,重复此过程直至发酵结束,实验结果如图2D 所示。分批发酵过程中流加L-谷氨酸,使得分批发酵时间延长至56.5 h,比未添加L-谷氨酸发酵时间延长4.5 h;甘油消耗速率[25.5 g/(L·d)]比未添加L-谷氨酸时甘油消耗速率[27.7g/(L·d)]也有所下降。但这和添加谷氨酸可以促进碳源消耗是不矛盾的。谷氨酸也可以作为碳源被细胞利用,在流加谷氨酸的试验中谷氨酸共添加45 g/L,谷氨酸的消耗速率可计算得到为19.1 g/(L·d)。因此,虽然流加谷氨酸导致甘油的消耗速率降低了,但是总的碳源消耗速率却提高了。值得一提的是,ε-PL 产量和产率分别为11.01 g/L 和4.68 g/(L·d),与未添加L-谷氨酸相比分别提高了67.1%和57.5%;DCW 为25.67 g/L,比未添加L-谷氨酸相比提高了63.0%。上述实验结果表明,分批发酵过程中连续多次流加L-谷氨酸相比于一次添加更有利于菌体的生长和ε-PL 的合成。
2.3 补料-分批发酵中流加谷氨酸对ε-PL 合成的影响
补料-分批发酵是大量生产ε-PL 的有效方法。为了获得更高的ε-PL 产量和产率,将上述建立的连续流加L-谷氨酸方式与补料- 分批发酵方法相结合,实验结果如图3A 所示。经过连续174 h 补料-分批发酵和8 次L-谷氨酸流加,最终实现ε-PL 产量和产率分别达到31.65 g/L 和4.36 g/(L·d),与未添加L-谷氨酸的补料-分批发酵ε-PL 产量(21.22 g/L)和产率[3.03 g/(L·d)]相比(图3B),分别提高49.2%和43.9%。由此可见,在补料-分批发酵过程中流加L-谷氨酸是实现高产量、高产量和高生产强度ε-PL 发酵的有效方式。
另外,流加L-谷氨酸的补料-分批发酵DCW 为37.7 g/L,与未添加L-谷氨酸的补料- 分批发酵的38.04 g/L 相当。虽然流加L-谷氨酸的补料-分批发酵的DCW 并未大于未添加L-谷氨酸补料-分批发酵的DCW,但从图3 中可以看出,流加谷氨酸补料-分批发酵前期的菌体生长速率要大于未添加谷氨酸补料-分批发酵前期的菌体生长速率。因此,流加谷氨酸的补料分批发酵更早达到菌体生长稳定期。到达菌体生长稳定期后,谷氨酸对菌体生长的促进效果没有在对数期那样明显,但更早地到达稳定期有助于在发酵高峰期累积大量菌体,从而提高ε-PL 产量。
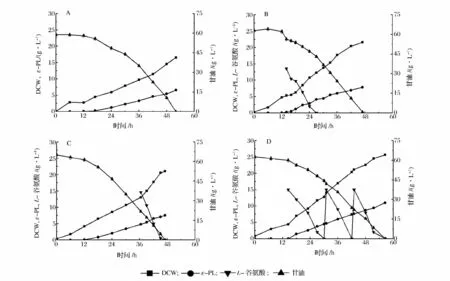
图2 分批发酵过程中L-谷氨酸添加时机对菌体生长和ε-PL 合成影响Fig.2 Effect of L-glutamate acid feeding time on cell growth and ε-PL production during batch fermentation by Streptomyces sp. M-Z18 in 5 L fermenter at pH 3.8
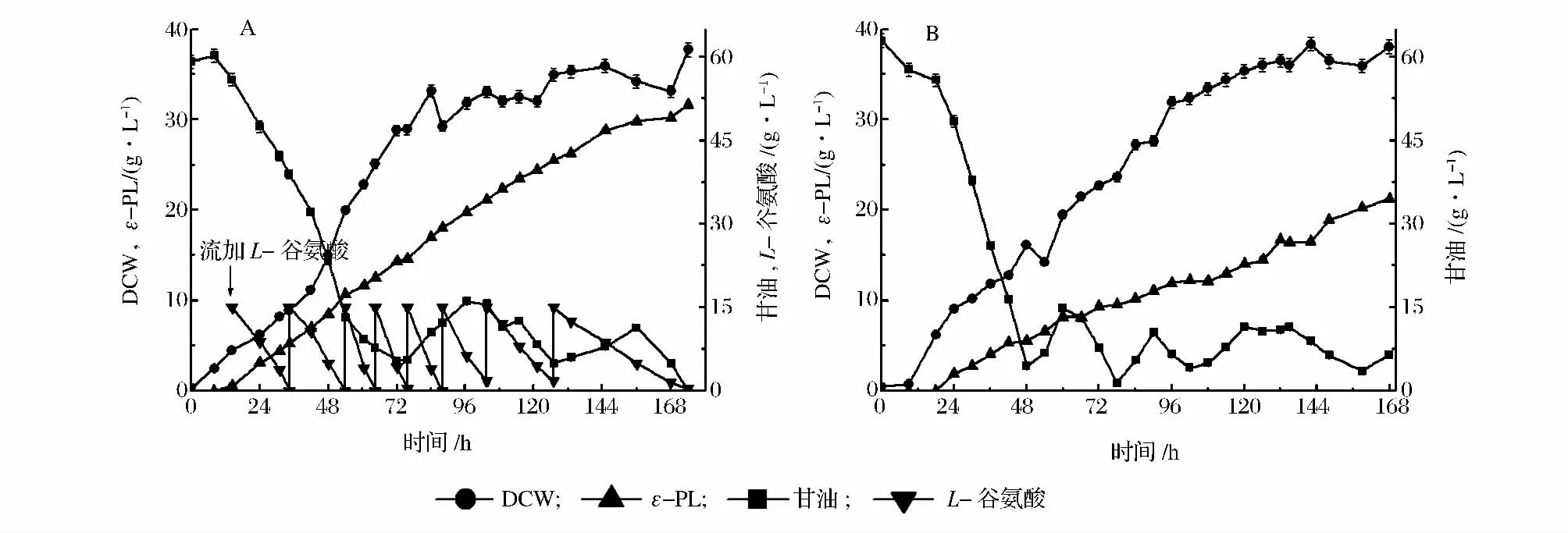
图3 补料-分批发酵中流加(A)和未流加(B)L-谷氨酸的ε-PL 发酵过程参数比较Fig.3 Comparison of ε-PL fed-batch fermentation process with and without L-glutamate acid feeding by Streptomyces sp. M-Z18 in 5 L fermenter at pH3.8
3 讨论
本研究以1 株S-2-氨基乙基-L-半胱氨酸抗性ε-PL 突变株Streptomyces sp. M-Z18 为对象,借助L-谷氨酸流加策略,实现ε-PL 补料-分批发酵产量和产率分别达到31.65 g/L 和4.36 g/(L·d),比原有发酵工艺分别提高49.2%和43.9%。可见,发酵过程中连续流加L-谷氨酸为进一步提高S-2-氨基乙基-L-半胱氨酸抗性突变株ε-PL 合成能力提供了参考。
L-谷氨酸提高ε-PL 合成能力的可能机制在于:L-谷氨酸通过转氨作用为ε-PL 合成前体L-赖氨酸合成过程提供氮骨架,同时生成的α-酮戊二酸进入三羧酸循环又进一步提供碳骨架,从而增加了L-赖氨酸合成能力,进而加快了ε-PL 合成,最终导致ε-PL产量提升。显然,上述L-谷氨酸的双重作用(碳源+氮源)还有待于利用稳定性同位素标记、关键酶活性和代谢通量分析等实验加以证实。
[1] Kazuya Y,Yoshimitsu H. Biotechnological production of poly-epsilon-L-lysine for food and medical applications[J]. Amino-Acid Homopolymers Occurring in Nature,2010,15:61 -75.
[2] Kahar P,Iwata T,Hiraki J,et al. Enhancement of ε-polylysine production by Streptomyces albulus strain 410 using pH control[J]. J Biosci Bioeng,2001,91(2):190 -194.
[3] Liu Sheng-rong,Wu Qing-ping,Zhang Ju-mei,et al. Production of ε-poly-L-lysine by Streptomyces sp. using resinbased,in situ product removal[J]. Biotechnol Lett,2011,33(8):1 581 -1 585.
[4] 贾士儒,莫治文,谭之磊,等. 一种提高ε-聚-L-赖氨酸产量的新方法[P]. 中国专利,CN101671703A.2010 - 03 -17.
[5] 张扬,张全景,冯小海,等. Kitasatospora sp. MY 5 -36 菌株补料重复发酵生产ε-聚赖氨酸[J]. 过程工程学报,2010,10(2):344 -347.
[6] Hiraki J,Hatakeyama M,Morita H,et al. Improved ε-poly-L-lysine production of an S-(2-aminoethyl)-L-cysteine resistant mutant of Streptomyces albulus[J]. Seibutu Kougaku Kaishi,1998,76:487 -493.
[7] 张超,王正刚,段作营,等. 大剂量紫外诱变选育ε-聚赖氨酸高产菌[J]. 生物加工过程,2007,5(3):64 -68.
[8] 陈玮玮,朱宏阳,徐虹.ε-聚赖氨酸高产菌株选育及分批发酵的研究[J]. 工业微生物,2007,37(2):28 -30.
[9] 贾士儒,董惠钧,姜俊云,等.ε-聚赖氨酸高产菌株的选育[J]. 食品与发酵工业,2004,30(11):14 -17.
[10] Takehara M,Hibino A,Saimura M,et al. High-yield production of short chain length poly(e-L-lysine)consisting of 5 -20 residues by Streptomyces aureofaciens,and its antimicrobial activity[J]. Biotechnol Lett,2010,32(9):1 299 -1 303.
[11] Chen Xu-sheng,Tang Lei,Li Shu,et al. Optimization of medium for enhancement of ε-poly-L-lysine production by Streptomyces sp. M-Z18 with glycerol as carbon source[J]. Bioresour Technol,2011,102(2):1 727 - 1 732.
[12] Chen Xu-sheng,Li Shu,Liao Li-juan,et al. Production of ε-poly-L-lysine using a novel two-stage pH control strategy by Streptomyces sp. M-Z18 from glycerol[J].Bioprocess Biosyst Eng,2011,34(5):561 -567.
[13] 陈卓贤,沈春明,陈国良. 味精生产工艺学[M]. 北京:中国轻工业出版社,1990:227.
