组蛋白去乙酰化酶5在非小细胞肺癌迁移中的作用研究
2011-05-30洪伟俊宁允叶胥武剑邵艳李强
洪伟俊 宁允叶 胥武剑 邵艳 李强
第二军医大学长海医院呼吸内科,上海 200433
肺癌是死亡率极高的恶性肿瘤,其中80%为非小细胞肺癌(non-small cell lung cancer,NSCLC),只有20%~30%的患者有手术机会,但是即使是在手术切除的早期NSCLC患者中,也有三分之二的患者存在复发和转移的危险,5年生存率仍然较低(8%~14%)。丛蕾等[1]研究发现,晚期NSCLC患者单纯化疗后中位生存期仅为12.2个月,1年生存率只有52.9%,复发和转移仍是导致死亡的主要因素。目前针对肺癌的主要治疗手段——放疗、化疗等对提高肺癌患者的生存率已达到一个平台期,深入探讨肺癌发生、发展的分子生物学机制,以寻找新的安全有效的治疗途径、改善预后成为肿瘤研究的热点。
组蛋白乙酰化状态是调控基因转录的关键,而组蛋白乙酰化水平受组蛋白去乙酰化酶(histone deacetylases,HDACs)和组蛋白乙酰化酶(histone acetyltransferases,HATs)调节,HDACs和HATs失衡是肿瘤发生、发展的重要分子机制。组蛋白去乙酰化酶5(histone deacetylase 5,HDAC5)是Verdel等[2]于1999年在小鼠基因组中发现的。人HDAC5基因定位于染色体17q21,广泛表达于肺、脑、心肌、骨骼肌、胎盘等多种组织。HDAC5在多种肿瘤中表达降低,但其在肺癌细胞迁移中的作用尚未见报道。本研究旨在检测肺癌组织及其相对应的癌旁组织中HDAC5基因表达情况;体外利用HDAC5基因过表达质粒,研究HDAC5对肺腺癌细胞NCI-H1299迁移的影响。
1 资料和方法
1.1 患者一般资料
所有标本均来自我院2009年4月—2009年11月间手术的NSCLC患者,其中男性15例,女性13例;腺癌20例,鳞癌8例;年龄41~70岁,平均年龄为57.8岁。在手术切除病灶后用锋利刀片迅速切取原发肿瘤和距离切缘2 cm处癌旁组织各1块,组织标本冻存于液氮中。
1.2 主要试剂
NCI-H1299细胞(购自中国科学院上海细胞所);胎牛血清(Hyclone);高糖DMEM培养基(Hyclone)、OPTI-MEMⅠ培养基(Gibco);兔抗人-HDAC5(SAB);辣根过氧化酶(HRP)标记的兔抗人GAPDH抗体(康成生物);HRP标记的山羊抗兔IgG(Abcam);RNA提取试剂盒(TRIzol)(Invitrogen),RT-PCR试剂盒(Takara),real time-PCR试剂盒(Takara);HDAC5野生型基因质粒(pDEST26-HDAC5wt)由澳大利亚迪肯大学Sean L.McGee教授惠赠。
1.3 方法
1.3.1 细胞培养
NCI-H1299细胞贴壁生长于含10%胎牛血清、100 U/mL青霉素、100 μg/mL链霉素的DMEM高糖培养基中,37 ℃、CO2体积分数为5%、相对湿度为90%的温箱(Heraus)内培养,当细胞融合达80%以上时0.25%胰酶消化传代。
1.3.2HDAC5基因转染及筛选
将NCI-H1299细胞接种于24孔板中,每孔为5×105个细胞,细胞融合至80%以上,用Fugene HD试剂盒进行转染。实验分组:转染HDAC5wt基因的实验组(HDAC5wt)、未转染任何质粒的正常对照组(control)、转染空质粒的阴性对照组(vector)。用OPTI-MEMⅠ培养基稀释质粒DNA至浓度为:2 μg DNA/100 μL OPTIMEMⅠ培养基,分别以Fugene HD转染试剂:DNA比例为2∶2、4∶2、6∶2配制转染复合物,室温下温育15 min,将转染复合物加入到培养细胞中,OPTI-MEM Ⅰ培养6~8 h后换成含10%胎牛血清的DMEM培养基继续培养,48 h后收集细胞提取蛋白。再以选定的最佳转染比例转染NCI-H1299细胞,36 h时加入含1 000 μg/mL G418和10%胎牛血清的DMEM培养基进行选择性培养,3周后筛选出阳性克隆扩增培养,并用500 μg/mL的G418维持筛选。
1.3.3 冻存组织处理
于液氮中取出标本,剪取少许组织并加入液氮后快速研磨,收集于2 mL EP管,分别加入TRIzol及RIPA蛋白裂解液后匀浆,提取mRNA、蛋白质,分别进行real time RT-PCR、Western blot实验。
1.3.4 Real time RT-PCR检测HDAC5 mRNA表达
用TRIzol法抽提总mRNA,测定其浓度和纯度,用RT-PCR逆转录试剂盒逆转录为cDNA后进行PCR扩增。HDAC5上游引物:5’-CCATCAGCCAGAAGATGTATGC-3’,下游引物: 5’-ATTCCTCGGCGTGGTGTCCT-3’,PCR产物201 bp;β-actin上游引物:5’-GGACTTCGAGCAAGAGATGG-3’,下游引物: 5’-AGCACTGTGTTGGCGTACAG-3’,PCR产物 234 bp。PCR条件:94 ℃预变性2 min,94 ℃变性30 s,60 ℃退火延伸40 s,30个循环;72 ℃终延伸5 min。
1.3.5 Western blot检测HDAC5蛋白
RIPA蛋白裂解液裂解细胞及组织,4 ℃、134 000×g离心20 min,取上清液即为蛋白提取物。用BCA法测定蛋白浓度,30 μg变性蛋白聚丙烯酰胺(PAGE,10%)凝胶电泳,转膜,5% BSA封闭,1∶1 000一抗4 ℃过夜,1∶2 000山羊抗兔HRP标记二抗室温反应1 h ,ECL显色,成像。
1.3.6 MTT比色法测定细胞增殖
稳定转染的H1299细胞(HDAC5wt)、空白质粒转染细胞(vector)、未转染细胞(control)融合达80%以上时,胰酶消化后形成单细胞悬液,以每孔5 000个细胞接种于 96孔板中,每组设10个复孔,分别于第1~5天时检测细胞增殖情况。细胞贴壁后进行初始490 nm波长光下的吸光度(D490)测定,之后分别于第1~5天分别测定各组细胞的D490值:每孔加入100 μL MTT液培养4 h后,去除培养基,加入DMSO 150 μL,室温下充分震荡10 min,用酶标仪测定D490。重复3次。
1.3.7 Transwell实验
分别将已融合至80%以上的3组细胞进行胰酶消化,形成单细胞悬液,872×g离心5 min后以无血清培养基重悬细胞,取200 μL无血清单细胞悬液(2×105个细胞)接种至置于24孔板内的8 μm PET膜的transwell 小室中,外室中加入含10%胎牛血清的DMEM高糖培养基,温箱中温育8 h取出,中性甲醛固定30 min,用棉签小心擦去上层未迁移细胞,用瑞氏染色、拍照,镜下计数6个视野中已跨膜迁移的细胞数目。
1.3.8 划痕实验
以每孔2×106个细胞接种于6孔板中,当细胞融合达90%~100%时,弃去培养基,以无菌PBS清洗细胞1次后,用200 μL枪头沿孔直径笔直划痕,用无菌PBS洗细胞3次,去除划下的细胞,加入无血清培养基,常规培养,分别在0、12、24 h观察、拍照。
1.4 统计学处理
所得数据用表示,应用SPSS 11.5软件包进行统计学分析,两样本均数间比较采用配对t检验,组间差异采用单因素方差分析(One Way ANOVA),差异显著者采用LSD-t检验进行两组间比较,P<0.05为差异有统计学意义。
2 结 果
2.1 HDAC5在肺癌组织标本中的表达
Real-time RT-PCR结果显示,28例组织标本中有20例肺癌组织中HDAC5 mRNA表达低于癌旁组织,其余8例HDAC5 mRNA表达稍微升高或无明显差异(图1)。Western blot检测NSCLC及癌旁组织HDAC5蛋白表达情况,结果显示,与癌旁组织相比,肺癌组织中HDAC5的蛋白表达明显降低(图2),其表达趋势与mRNA表达一致,提示HDAC5在NSCLC中的低表达可能与NSCLC的发生、发展相关。
2.2 体外细胞转染中转染试剂与质粒的转染比例筛选
分别以Fugene HD转染试剂与质粒比例为 2∶2、4∶2、6∶2配制转染复合物,转染NCI-H1299细胞,如图3所示,3种比例的转染试剂都成功将HDAC5wt基因转入目的细胞,其中4∶2组转染效率明显高于2∶2组,即前者HDAC5蛋白表达量高于后者;而6∶2组质粒转染后,HDAC5蛋白表达量与4∶2组相近。因此,采用Fugene HD转染试剂∶质粒=4∶2的转染比例进行后续相关研究。

2.3 基因过表达鉴定
分别将HDAC5wt质粒及空质粒转染入NCI-H1299细胞后,用G418进行筛选,形成稳定表达株。与未转染组及空质粒组相比,HDAC5wt组细胞中HDAC5的转录水平显著提高(图4),蛋白表达量明显增加(图5)。

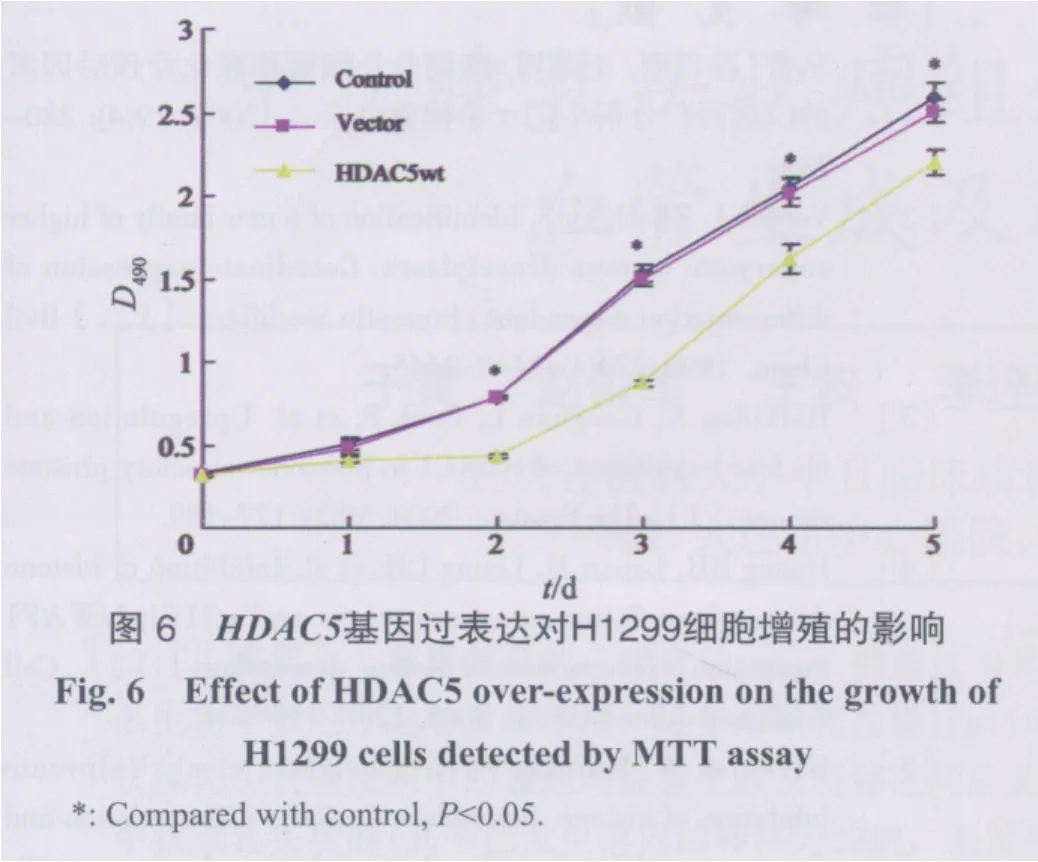
2.4 HDAC5基因对H1299细胞增殖的影响
采用MTT法检测control、vector、HDAC5wt质粒转染3组细胞增殖情况,细胞贴壁后3组细胞初始D490值无统计学差异,表明3组细胞接种数量一致。在24 h时,3组间D490无统计学差异;在第2~5 天对数生长期内实验组细胞D490值较正常对照组及阴性对照组显著降低,而正常对照组与阴性对照组之间D490在同一检测时间点无显著性差异(图6)。
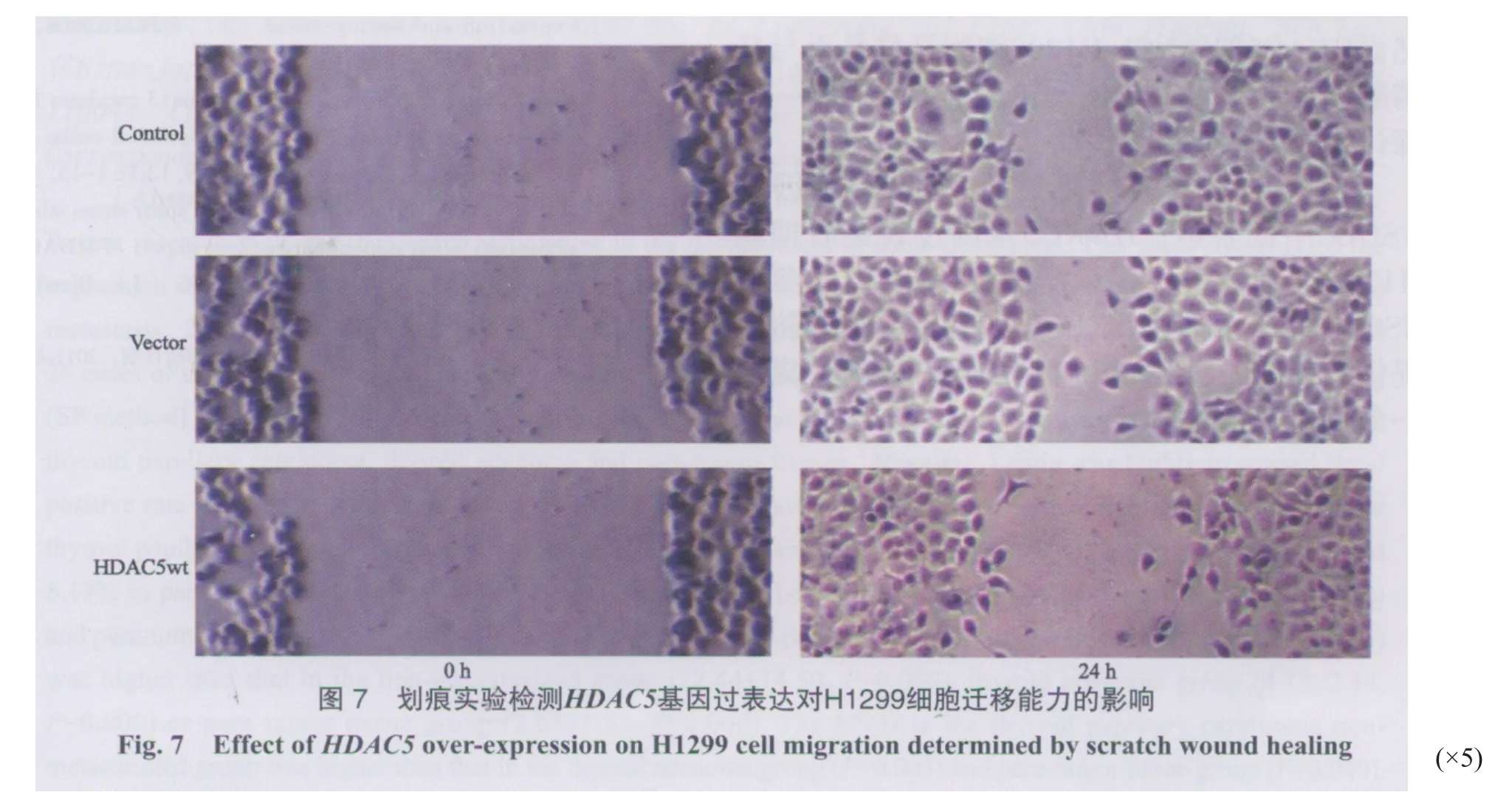
2.5 HDAC5过表达对H1299细胞迁移的影响
我们采用Transwell检测HDAC5基因对H1299细胞跨膜迁移能力的影响。结果显示,实验组(158.3±19.4)跨膜迁移细胞数显著低于正常对照组(247.0±12.1)及阴性对照组(236.0±14.9,P均<0.05),而正常对照组与阴性对照组间穿膜细胞数无显著差异,提示HDAC5过表达抑制了NCI-H1299细胞跨膜迁移能力。
同时,我们采用划痕实验直观地观察HDAC5基因对H1299细胞迁移能力的影响。结果如图7所示,与初始划痕宽度相比,在24 h内3组细胞的划痕都有不同程度的愈合。HDAC5组细胞在24 h时划痕愈合程度明显低于其他两组,表明该组细胞的迁移能力低于其他两组,从另一个角度证实了HDAC5基因过表达抑制NCI-H1299细胞迁移。
3 讨 论
HDACs和HATs在肿瘤细胞基因转录调控中起重要作用。HDACs使组蛋白去乙酰化后恢复带正电荷的特性,并与带负电的DNA紧密结合,染色质呈致密卷曲的阻抑结构,从而抑制基因转录;HATs通过在组蛋白的N-端赖氨酸残基上引入乙酰基,促进基因转录。两者在体内处于一个动态平衡,共同参与基因的表达调控。一旦这种平衡破坏,细胞便异常增殖、分化,最终导致肿瘤的发生发展。HDACs在不同肿瘤中存在差异性表达,其中大多数亚型在多种肿瘤中呈高表达状态:HDAC1在肺癌、前列腺癌、胃癌、结肠癌和乳腺癌中高表达[3],HDAC2在胃癌、结直肠癌和宫颈癌中高表达[4-5];目前仅发现HDAC5在多种肿瘤中低表达。Osada等[6]报道HDAC5基因表达与NSCLC预后显著相关,HDAC5表达降低的患者其预后较差。本研究检测了HDAC5在28例NSCLC组织及相应癌旁组织中的表达,结果证实HDAC5在癌组织较对应的癌旁组织表达明显降低,提示HDAC5基因的低表达可能在NSCLC的发生、发展中起重要作用。Urbich等[7]的研究表明HDAC5抑制血管形成,因此推测HDAC5可能与肺癌的转移相关。
将HDAC5质粒转染至人肺腺癌细胞NCI-H1299,分别采用Transwell方法研究细胞跨膜迁移能力、采用划痕方法研究细胞侧向迁移能力,结果表明,HDAC5基因过表达抑制H1299细胞迁移。由于在24 h时过表达HDAC5基因并未影响H1299细胞的增殖能力,因此排除了细胞迁移研究中因过表达HDAC5基因后细胞增殖差异对跨膜迁移细胞量及划痕愈合能力的影响,从而证实HDAC5基因抑制H1299细胞迁移。
HDAC5蛋白上有多个基序:核定位信号基序(NLS)、出核信号基序(NES)、C-末端结合蛋白基序(CtBP)、肌增强子2结合基序(MEF2)、14-3-3伴侣蛋白结合基序、酶活性区域(DAC)等,HDAC5在不同的生理病理过程中发挥不同作用,参与这一过程的基序有所差异。HDAC5被磷酸化后可以在细胞核和细胞质之间穿梭运动。Urbich、Roy等[7-8]研究表明,HDAC5及其磷酸化后的核定位在肿瘤细胞的增殖、血管内皮细胞的迁移过程中起重要作用。而多种因素参与或影响HDAC5的磷酸化及其核质转运过程,如血管内皮生长因子(VEGF)、Ga13蛋白、血管紧张素Ⅱ(AngⅡ)、流体剪切力等[9-11]。然而HDAC5抑制NCI-H1299细胞迁移是否与其磷酸化后的核定位或核质转运相关尚需进一步探讨。
综上,本研究结果显示HDAC5基因在NSCLC中低表达;HDAC5基因过表达后抑制H1299细胞迁移,提示HDAC5基因可能参与NSCLC的转移,为NSCLC患者晚期转移的防治提供一种新的思路,为抑制肿瘤细胞转移提供一个新的靶点。
[1]丛蕾, 崔言刚, 王潍博.晚期非小细胞肺癌化疗预后因素的COX回归分析[J].中国癌症杂志, 2009, 19(4): 280-283.
[2]Verdel A, Khochbin S.Identification of a new family of higher eukaryotic histone deacetylases.Coordinate expression of differentiation dependent chromatin modifiers [J].J Biol Chem, 1999, 274(4): 2440-2445.
[3]Halkidou K, Gaughan L, Cook S, et al.Upregulation and nuclear recruitment of HDAC1 in hormone refractory prostate cancer [J].The Prostate, 2004, 59(2): 177-189.
[4]Huang BH, Laban M, Leung CH, et al.Inhibition of histone deacetylase 2 increases apoptosis and p21Cip1/WAF1 expression,independent of histone deacetylase 1 [J].Cell death and differentiation, 2005, 12(4): 395-404.
[5]Hrzenjak A, Moinfar F, Kremser ML, et al.Valproate inhibition of histone deacetylase 2 affects differentiation and decreases proliferation of endometrial stromal sarcoma cells[J].Molecular cancer therapeutics, 2006, 5(9): 2003-2010.
[6]Osada H, Tatematsu Y, Saito H, et al.Reduced expression of class II histone deacetylase genes is associated with poor prognosis in lung cancer patients [J].Int J Cancer, 2004,112(1): 26-32.
[7]Urbich C, Rössig L, Kaluza D, et al.HDAC5 is a repressor of angiogenesis and determines the angiogenic gene expression pattern of endothelial cells [J].Blood, 2009, 113(22):5669-5679.
[8]Roy S, Shor AC, Bagui TK, et al.Histone Deacetylase 5 Represses the Transcription of Cyclin D3 [J].J Cell Biochem, 2008, 104(6): 2143-2154.
[9]Ha CH, Wang W, Jhun BS, et al.Protein kinase D-dependent phosphorylation and nuclear export of histone deacetylase 5 mediates vascular endothelial growth factor induced gene expression and angiogenesis [J].J Biol Chem, 2008,283(21): 14590-14599.
[10]Liu G, Han J, Profirovic J, et al.Galpha13 regulates MEF2-dependent gene transcription in endothelial cells: role in angiogenesis [J].Angiogenesis, 2009, 12(1): 1-15.
[11]Wang W, Ha CH, Jhun BS, et al.Fluid shear stress stimulates phosphorylation dependent nuclear export of HDAC5 and mediates expression of KLF2 and eNOS [J].Blood, 2010,115(14): 2971-2979.
