喀斯特地区白三叶形态和遗传多样性研究
2010-07-13李莉王元素洪绂曾
李莉,王元素,洪绂曾
1.贵州师范大学生命科学学院,贵州 贵阳 550001;2.中国农业大学动物科技学院草地所,北京 100094;3.贵州省草原监理站,贵州 贵阳 550001
白三叶(Trifolium repens)是温带最重要的豆科牧草,营养价值高,家畜喜欢采食[1],其根际共生根瘤菌固定大气中的氮,因此,是减少N肥施用的最佳选择,已经在有机农场广泛应用[2]。白三叶是我国喀斯特地区混播草地中最稳定的牧草种群之一[3,4],而且侵袭性最强[5],一些研究并在竞争与共存方面探索了稳定机理。
从分子生态学的角度探讨白三叶形态进化、遗传多样性与种群稳定持久性的关系方面,国内外的研究不多,国内只有张贤等人(2006)研究了白三叶遗传空间多态性[6]。由于白三叶基因多样性受微环境的影响很大,不同地区其遗传变异性不尽相同。因此,开展白三叶空间和时间上的遗传多样性研究,对理解喀斯特地区白三叶种群的稳定持久性有重要的意义。
1 研究方法
在贵州省威宁县对3个不同年限的白三叶地块进行调查,收集种子进行温室培养,分别进行形态学观测;取新生叶片进行DNA提取与扩增,分析遗传多样性。
1.1 样地描述
A:100年白三叶。威宁县石门坎乡,英国传教士柏格理(Samuel Pollard) 1900年到1915年在贵州威宁一带传教,在石门坎苗族聚居地建立教会学校—石门坎中学,于 1906年从英国带来白三叶、多年生黑麦草(Lolium perenne)等牧草,于 1906—1908年建植了一块白三叶作为绿化观赏,并建设了一个草坪足球场和一个足球练习场[7]。2006年6月调查时白三叶和草地早熟禾(Poa pratensis)仍有相当比例,侵入的杂草主要有平车前(Plantago depressaWilld.)。种子主要取自足球练习场。
B:40年白三叶。1965年,贵昆铁路通车,在威宁金钟镇有一小站—树舍。威宁黑山羊久负盛名,特别在石门坎等苗族聚居区,喜欢养山羊。从1965—1970年,威宁县食品公司组织羊只用人赶往树舍火车站装车(从石门坎山羊养殖地到火车站距离大约120 km)。1965开始,由山羊传播的白三叶就长满了沿途路边和火车站周围。2006年9月采集种子。
C:20年白三叶。1985年在灼圃示范牧场播种的混播试验地。2006年9月采集种子。
1.2 形态学测定
种床准备:50格种苗培养盘,每格规格 10 cm×10 cm;床土为m(石英砂)︰m(细土)︰m(腐殖质)=3︰4︰3混合而成,高压灭活。
种子处理:播种前为破除硬实,用细砂与种子混合,沙磨。
播种:每个样种50格,每格10粒种子,浇透水后播种,覆土1 cm。
管理:浇水按需进行,晴天每天一次,阴天2~3 d一次。N、P、K全肥每2月一次,满足生长需求。成苗后间苗,每格只留下1株苗。
监测指标:中叶长、宽以及长宽比,叶质量,茎质量,叶高,生长点个数,匍匐茎长度等。
1.3 遗传多样性分析
白三叶分子标记采用张贤等人(2006)[6]的优化 RAPD分析条件。10碱基随机引物(5′GTGACGTAGG 3′) 购自上海生工公司。100bp DNA ladder、Taq DNA聚合酶、dNTP购自天为时代公司。PCR仪为PTC200型,电泳仪为DYY-6B型。
方法概述如下:白三叶DNA提取和RAPD反应程序及其优化条件参照张贤等(2006)[6]。遗传多样性用Alpha Imager 2200软件分析。
2 结果与分析
2.1 白三叶形态学特性分析
不同时间和空间的白三叶形态学特征有很大的差异性(表1)。开始分枝即80日龄的单株叶片数以C样(100年草地)的最高,显著高于A样(20年草地)和B样(40年草地)(P<0.05),但种群内的变异系数也最高,分别比A样和B样高出20.39和25.85个百分点,单株叶片数最多的是最少的16倍。A样和B样的叶片数、变异系数和数量范围都非常接近。

表1 白三叶形态学数量性状分析Table 1 Visible traits of white clover
到5月龄时,单株叶片数与80日龄时有所不同。平均单株叶片数以A样的最低(P<0.05),只有B样和C样的70%左右。但种群内变异系数最大的仍然是C样,分别比A样和B样高18.83和23.27个百分点,最多株是最少株的32倍。B样和C样的变异系数接近。
5月龄时第 3叶片中叶的长度以 A样的最长(P<0.05),分别是B样和C样的110%和118%;三个样地的种群内变异系数很接近,在 20.58%~23%之间,最低与最大值范围也非常接近。
5月龄第三叶片的宽度三个样地之间差异显著(P<0.05),最宽的是A样,其次是B样,宽度最小的是C样。三个样地种群内变异系数范围在20%左右,A、B、C样种群内叶宽最大值分别是最小值的2.91、2.55和2.40倍,而种群之间差异不大。
中叶长宽比主要用来描述叶片的形状。长宽比值以C样的最大(P<0.05),其比值大于1,说明C样的叶长大于宽,为椭圆形;A和B样的比值都小于1,说明二者的叶片多为心形。三个样地的种群内变异系数都不大,表明叶形是稳定的形态学性状。
叶片高度以A样最高(P<0.05),分别比B样和C样高出14.46%和21.87%。三个样的种群内变异系数比较大,都接近30%。
每株的生长点个数以A样的最少,分别比B样和C样低30.37%和31.91%。但三个样种群内的变异系数都很大,其中最高的是C样(64.42%),分别比A样和B样高出18.36和15.68个百分点。
单株匍匐茎长度(分枝长)用来显示白三叶的侵占力和地上生物量。匍匐茎最短的是 A样(P<0.05),仅相当于 B样和 C样的 36.39%和34.70%。三个种群的变异系数却非常高,分别为128.69%、108.31%和138.87%。比如C样中,最短的一株还没有分枝,而最长的已经达到47.50 cm,说明种群内个体之间差异非常大。
基于5月龄各形态学变量之间Pearson Correlation进行的系统树状聚类图见图1,可以看出,叶数、生长点和分枝长度有很紧密的相关性,白三叶分枝越长,生长点个数越多,叶数就越多。而中叶的长宽比(即叶形)与叶层高、中叶长、中叶宽相关,其中叶层高与中叶长相关性高。

图1 白三叶形态学变量相关性聚类树状图Fig.1 Hierarchical cluster of Pearson correlation of morphyologocal variables

图2 三个年龄白三叶种子温室培养Fig.2 Cultivating in glasshouse
图2直观地表现了三个时间段白三叶形态学特征的差异。温室培养80日龄时,100年和50年的已经开始分枝,叶数多,而20年的个体小,基本上没有分枝,叶数少。到180日龄,100年白三叶分枝多、匍匐茎长,叶片茂密、斜生,20年的仍然分枝很少,叶片少而直立;而40年的介于二者之间。
在温室培养180日后,对所有植株进行质量相关特征的分析。结果表明(表2),三个不同年龄草地处理的平均单株叶质量、根质量、地上生物量、地上DM与地下DM的比值等指标数值接近,差异不显著。但是,与茎有关的指标如茎质量等差异显著。即草地年限越大的白三叶,茎的生物量越高。20年草地(A)的茎质量和茎根质量比值显著低于40年(B)和 100年草地的(C)(P<0.05),只分别为B和C的57%~47%。而A的叶茎比值显著高于B、C草地的(P<0.05),分别为B和C的2倍和4倍。
叶质量、茎质量、根质量、地上生物量等指标的种群内变异系数则随着草地年限的增加而增大。如20年草地的个体变异系数为0.451,而100年的则为 0.950,增大了 1倍多。叶茎质量比、茎根质量比、地上与地下生物量等比值则变异系数相似,不随草地年限的增加而变化。

表2 白三叶质量特征分析Table 2 Weight-related traits of white clover1)
2.2 白三叶遗传多样性分析
从图3和图4可以看出,不同年龄的白三叶遗传多样性差异显著。20年白三叶有14个等位基因,而100年白三叶只出现8个等位基因(40年的由于保存失误,使样品损坏)。20年白三叶中,等位基因M频率最高,为17.24%,其次为F和K,均为10.34%,最低的是A、B、H、L,都只出现3.45%。100年白三叶,等位基因K频率最高,为32.26%,其次为E,16.13%,而等位基因B、C、F、L、M、N都没有出现,频率为0。
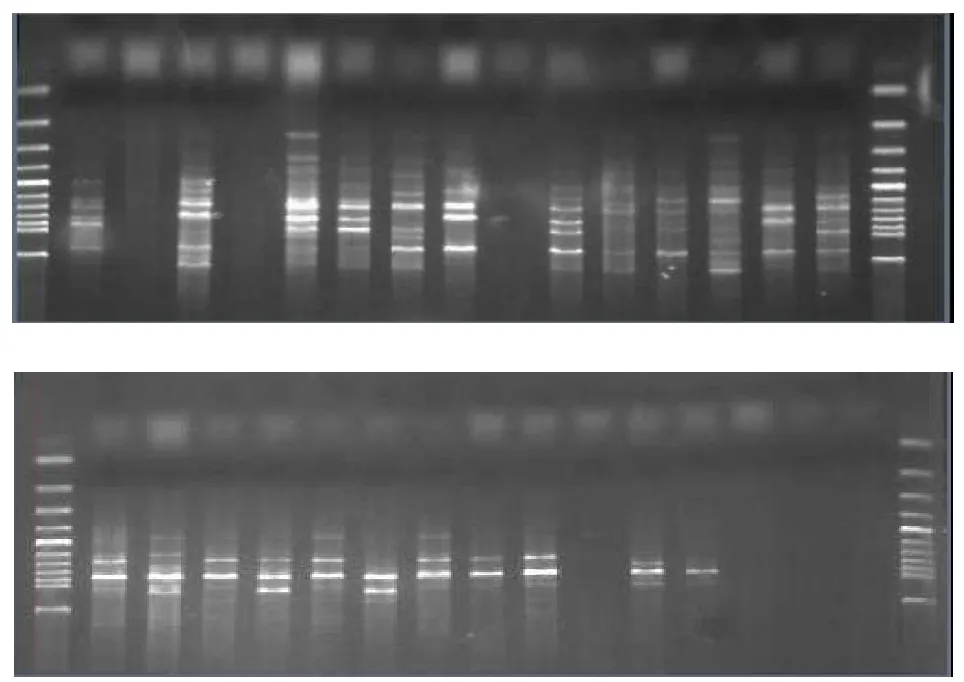
图3 不同年龄段白三叶RAPD标记Fig.3 RAPD of two ages white clovers
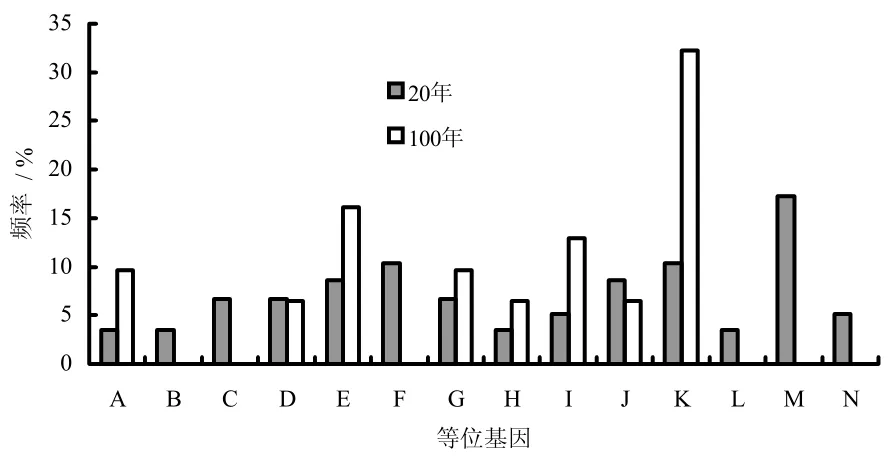
图4 不同年限白三叶基因频率分析Fig.4 ALLEL frequance of two different age white clovers
3 讨论与结论
随着年限的增加,白三叶形态学特征发生变化以适应环境,单株叶数、生长点数、中叶长、中叶长宽比以及种群内个体之间变异性随着年限的增加而增加,而叶层高度、中叶宽则下降。三个不同年龄草地处理的平均单株叶质量、根质量、地上生物量、地上生物量与地下生物量的比值等指标数值接近,差异不显著(P>0.05)。但是,与茎有关的指标如茎质量等差异显著。这是白三叶为适应踩踏和或家畜采食而产生的形态学变化。植物个体水平上对利用或放牧有两种响应对策:短期发生生理反应以应对光和组织被采食后碳水化合物的供给不足;长期则发生形态学的变化进化“回避”机制以减少被采食的机会而持久存在[8]。“回避”是指减少被采食的可能性和强度,其机理包括结构特性、物理特性和化学物质的变化,“忍耐”机理则包括被牧食后残留分生组织来源和有效性以及促进再生的生理过程[9]。在长期放牧下“回避”型植物呈垫伏状,“回避”机制可能比“忍耐”机制更有效[10]。本研究中,白三叶随年限增长,叶片数减少,而茎生物量增加,这是“回避”机制的结果。Rhoades(1985)[11]把回避机制随放牧强度增加而增强的现象叫诱导防御(inducible defences)。白三叶虽然对环境因素敏感,对干旱、霜冻和荫蔽耐受力差,但耐重牧和踩踏[12]。白三叶的匍匐茎贡献很大的生物量,它紧贴地面生长,不容易被采食和损害[4]。
100年白三叶的等位基因数和遗传多样性指数没有 20年的高,这是因为,首先,白三叶是严格的异花授粉植物,在利用条件下以营养生殖为主,以种子自熟下繁为辅;一般认为在基因流方面有大面积的连续分布[2]。一个白三叶植株可产生很多的匍匐茎和分枝,从而形成一个由众多克隆构件组成的占据一定面积的体系。这样的克隆斑块可从几平方厘米到几平方米的面积[13]。经过100年的变迁,白三叶种群很可能由少数的大克隆体占统治地位,增加了取样几率。第二,100年白三叶所在地位置偏远,交通极不发达,外来白三叶基因的侵入机会较小;而 20年白三叶位于灼圃示范牧场,引进的其他白三叶品种增加了基因漂流和融合的机会。
[1]BAKER M J, WILLIAMS W M.White clover[M].Blackwell Scientific Publication, Oxford, UK, 1987.
[2]VAN TREUREN R, BAS N, GOOSSENS P J, et al.Genetic diversity in perennial ryegrass and white clover among old Dutch grasslands as compared to cultivars and nature reserves[J].Molecular Ecology, 2005,14: 39-52.
[3]蒋文兰.贵州威宁混播草地初级生产力及群落稳定性调控途径的研究[D].甘肃农业大学.1991: 13-14, 19, 79-80.JIANG Wenlan.Controlling and Regulating primary Productivity and Association Stability of Mixed Pasture in Weining County of Guizhou Province[D].Gansu Agricultural University.1991: 13-14, 19, 79-80.
[4]王元素, 蒋文兰, 洪绂曾, 等.白三叶与不同禾草混播群落17 年稳定性比较研究[J].草业学报, 2006, 15(3): 55-62.WANG Yuansu, JIANG Wenlan, HONG Fuzeng, et al.A 17 year comparative study on the stability ofTrifolium repens/grass communities[J].Acta Prataculturae Sinica, 2006, 15(3): 55-62.
[5]王元素, 洪绂曾, 蒋文兰, 等.喀斯特地区红三叶混播草地群落对长期适度放牧的响应[J].生态环境学报, 2007, 1: 117-124.WANG Yuansu, HONG Fuzeng, JIANG Wenlan, et al.Responses ofTrifolium pratensemixed communities to long-term moderate grazing in Karst region[J].Ecology and Environment, 2007, 16(1): 117-124.
[6]张贤, 张英俊, 吴维群.白三叶RAPD分析条件优化[J].草地学报,2006, 14: 219-222.ZHANG Xian,ZHANG Yingjun,WU Weiqun.Optimization on RAPD analytical system of White Clover[J].ACTA AGRESTIA SINICA, 2006,14:219-222.
[7]向郢.福音下的石门坎[N].南方周末, 2006-10-19(第32版).XIANG Yin.Shimenkan under the Gospel[N].Southern Weekly,2006-10-19(D32).
[8]LEMAIRE G,CHAPMAN D.Tissue flows in grazed communities.HODGSON J,ILLIUS AW.The Ecology and Management of Grazing Systems[C].1996, CAB International.
[9]BRISKE D D.Strategies of plant survival in grazed systems: A functional interpretation.In: HODGSON J, ILLIUS AW.The Ecology and Management of Grazing Systems[C].1996, CAB International.
[10]DETLING J K, PAINTER E L.Defoliation responses of western wheatgrass populations with diverse histories of prairie dog grazing[J].Oecologia, 1983, 57: 65-71.
[11]RHOADES D F.Offensive-defensive interactions between herbivores and plants: their relevance in herbivore population dynamics and ecological theory[J].American Naturalist, 1985, 125: 205-238.
[12]GUSTINE D L, HUFF D R.Genetic variation within and among white clover populations from managed permanent pastures of the northeastern USA[J].Crop Science, 1999, 39: 524-530.
[13]GUSTINE D L, SANDERSON M A.Molecular analysis of white clover population structure in grazed swards during two growing seasons[J].Crop Science, 2001, 41: 1143-1149.
