环境因素对干旱-半干旱区城市银杏叶片碳同位素组成的影响
2010-07-13胡清静张成君郭景褚兆晶安娟
胡清静,张成君,郭景,褚兆晶,安娟
兰州大学资源环境学院,甘肃 兰州 730000
影响植物同位素组成(δ13C)的因素较多。例如,许多生物因素(植物间的竞争、植物的不同发育期、植物的不同部位、化学组成、叶龄、叶片的结构等)和非生物因素(水分、温度、湿度、光、CO2浓度、海拔高度、土壤盐分、土壤养分、大气污染物等)。研究表明,水分条件对植物同位素组成的影响较大,并对此做了大量的研究工作。同时,大气中CO2浓度的变化也是改变植物同位素组成的主要因素之一。Farquhar等[1]研究表明,植物的δ13C值可由下式来表示:
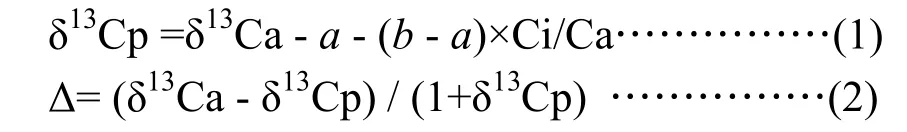
式(1)和式(2)中,δ13Cp和δ13Ca分别为植物组织及大气CO2的碳同位素比率,a和b分别为CO2扩散和羧化过程中的同位素分馏,而Ci和Ca分别为细胞间及大气的CO2浓度。
在植物生长过程中,植物叶片的气孔参数随环境参数影响的变化较大,叶片气孔的变化可能影响到a、Ci等的变化,进而影响到植物同位素组成的变化,而目前对这方面的研究还不是很成熟[2-4]。城市环境中影响植物生长的环境因素相对自然环境复杂,因此,本文主要针对干旱-半干旱区城市中植物叶片碳同位素组成随生长季节的变化,了解叶片气孔的发育过程,并了解降水、光照、大气污染物浓度等环境指标的变化对植物叶片碳同位素组成的影响,从而进一步认识影响植物叶片碳同位素组成的主要环境因素和特征。
1 样品的采集和分析
植物银杏(Ginkgo biloba)叶片样品采自兰州大学校园内。兰州地处内陆,属中温带大陆性气候,冬无严寒、夏无酷暑,气候干燥,气温日差较大,光照充足,太阳辐射强。年平均气温在0~14 ℃之间,年均降水量300 mm左右,降水各季分配不匀,主要集中在6~9月,无霜期180 d以上。兰州市区植物大约四月初开始发芽长叶。从2009年4月15日至2009年10月15日,每15天采集一次,分东西南北四个方向采集不同银杏叶片。各个方向离地面高大约2米左右,采集5~6片。主要采集成熟叶片,并截取每片叶片的中心部位进行前处理。陈立群[5]分析表明在同一片叶子上,中间部位是气孔密度和气孔指数最稳定的区域。采集后的银杏叶片在实验室内用蒸馏水洗净后,部分叶片在45 ℃恒温下干燥48 h以上,烘干用做碳同位素组成分析。另外2~3片银杏叶用双氧水和冰醋酸铵溶液(体积比1∶1)浸泡后,在70 ℃下煮3~4 h,直至叶片褪成白色为止。在加热过程中上下表皮间有气泡逸出,可以将上下表皮分开。然后将叶片中心部位的上下表皮平铺在同一载玻片上,并用番红染色。再在载玻片上滴加甘油,尽量消除气泡。多余的甘油用滤纸吸取,干后用指甲油封上盖玻片进行气孔参数测定。
采集的银杏叶片主要进行同位素组成和气孔参数测定。同位素分析将烘干后的银杏叶片研磨成60~100目的粉末,将大约0.1~1 μg粉末样装入石英舟中,在氧气流下,850 ℃加热约15 min,产生的CO2气体通过液氮、酒精-液氮冷阱收集、纯化后引入到 MAT-252稳定同位素质谱仪中分析碳同位素组成。结果以PDB-δ表示,样品分析精度≤0.2‰。样品在中国科学院兰州地质研究所测定。
将封上盖玻片的上下表皮层载玻片在兰州大学实验中心的高倍银光显微镜下观察并拍照。其中测定气孔密度样品在10×10倍镜下拍照(每个样品拍5~6张)并统计,测定气孔长、宽的样品在10×40倍镜下拍照(每个样品拍 8~10张)并统计。照片在image 5.0软件中观测气孔密度和气孔的大小。
2 结果与分析
银杏叶片碳同位素组成分析主要包括从4月15日开始大约每隔 15天采集一次东、南、西、北四个方位叶片的碳同位素组成,以及这四个方向叶片混合后的碳同位素组成。不同方向叶片的碳同位素组成分析结果见图 1。混合叶片碳同位素组成分析结果见图 2。从四月初生长开始,到大约十月中旬天气变冷,植物停止生长为止,不同方向的银杏叶片碳同位素组成总体上逐渐偏轻变负,但是不同方向叶片的变化没有太明显的规律性。从五月份以来,上旬不同方向叶片同位素组成相对偏重,下旬偏轻。在九月上旬不同方向叶片同位素组成最轻。不同方向的混合叶片碳同位素组成总体趋势与不同方向叶片碳同位素组成总趋势一致。
针对兰州市区银杏叶片在整个生长季中气孔的演化特征,生长季中降水、温度、光照和城市中比较特征的大气污染物浓度的变化(转换为污染指数的变化)对叶片碳同位素组成的影响进行分析(图2),图2中标出了不同参数与叶片碳同位素组成之间的相关系数值(r)。将气孔比表面积(单个叶片中气孔的总面积与叶片面积之比)、气孔面积(单个叶片中气孔的总面积)、气孔密度(单个叶片中气孔的总个数)作为气孔的主要参数,以了解气孔参数特征对叶片同位素组成的影响。
2.1 光照对银杏叶片同位素组成的影响
研究表明,植物组织、甚至组成植物叶片类脂化合物的碳同位素组成受光照的影响比较明显[6]。一般来说,阳面叶的同位素组成偏重,阴面叶的同位素组成偏轻[7]。Lockheart等[7]和 Farquhar等[8]研究认为光强度的下降会导致植物叶片有较高的胞间和大气CO2浓度比值(Ci/Ca)。根据式(1),当植物碳同位素组成在气孔关闭时,Ci值减小;相反,气孔张开时,Ci值增加。光照对气孔的开闭有直接的影响,光照加强时,气孔关闭,可能的原因是防止水分的过度蒸腾[9]。植物冠层中阳面叶比阴面叶具有较高的δ13C值, 也是由于阳面叶蒸发加强导致水分消耗增多, 因此气孔关闭程度较阴面叶高,以利于植物的生长[10]。Kürschner 等[11]和 Poole 等[12]的研究也表明阳面叶和阴面叶的气孔特征有极大的差异性,表明光照对植物叶片气孔影响较为直接,而叶片通过气孔的开闭适应生长的需要,进而影响了叶片碳同位素组成特征。
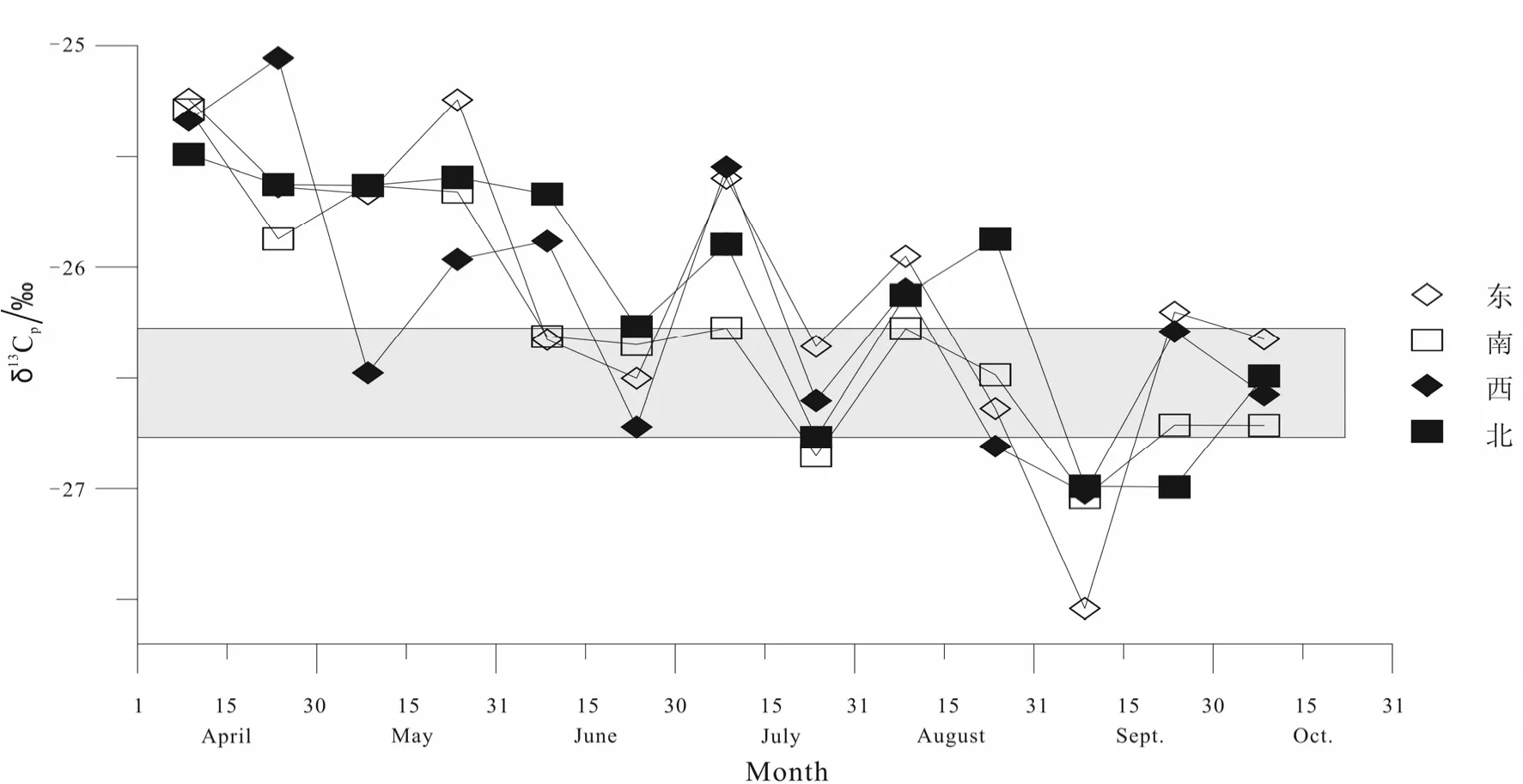
图1 不同方位银杏叶片碳同位素组成Fig.1 The carbon isotopic compositions of Ginkgo leaves at difference directions

图2 银杏叶片碳同位素组成与气孔参数、环境因素之间的相关性分析Fig.2 The comparing among the carbon isotope of Ginkgo leaves and parameters of leaves and environmental parameters
然而,本文对兰州市校园中银杏叶片生长期中的同位素分析结果发现(图1),同位素组成与光照(方向性)表现不是很明显。一般来说,南方和西方的光照时间及强度相对较大,但总体上没有十分明显的规律性。Lockheart 等人对相同样品的分析结果显示出,没有1995年采集的样品差异性强[6-7],阳面叶和阴面叶之间的同位素组成变化不如以前的明显。这其中的规律性我们目前还不十分清楚,但城市中植物生长环境与较空旷的自然环境存在一定的差异性。例如,周围高大建筑影响太阳对植物叶片的照射、城市中的大气环境条件等的影响,还需要进一步观察分析。
Farquhar等[1,8]和 Vogel[13]的研究表明,影响植物叶片碳同位素组成的因素主要有3个,包括大气CO2的δ13C、大气CO2的浓度(Ca)、以及CO2在叶片胞间浓度(Ci)与大气中CO2的浓度的比值(Ci/Ca),也可以表示为压力比值(pi/pa)。在一定地区,δ13C和Ca基本保持不变,因此,影响叶片碳同位素组成主要是Ci/Ca,或者pi/pa。而这个比值随气孔参数变化,即是说光照的变化对叶片气孔参数影响较大。但是,我们的分析结果中(图3)气孔密度、气孔比表面积、气孔平均面积等参数也没有明显的规律性。说明影响气孔的因素不仅是光照,可能还有其它的一些因素。

图3 银杏生长期中叶片气孔参数的变化特征Fig.3 The characteristic of the stomatal parameters in the growing period of Ginkgo
2.2 降水和温度对植物叶片碳同位素组成的影响
图2分析表明,银杏叶片碳同位素组成与降水量(r=0.586)、蒸发量(r=0.492)、温度(r=0.207)和有效湿度(r=0.761)之间均有一定的相关性。特别是与有效湿度之间的相关性最好(r=0.761,P<0.01),说明有效湿度是控制植物碳同位素组成的一个主要环境因素。
目前的研究发现,植物碳同位素组成与降水量的关系十分密切,尤其是在干旱-半干旱地区[14,15,16]。例如,ZHANG等对中国西北地区和青藏高原现代植物的调查表明,C3植物的碳同位素组成与降雨有明显的关系,干旱环境C3植物的碳同位素值较重[15]。同时,王国安[16]分析发现了北方C3草本植物碳同位素组成与湿度有关,而某些种类的C3植物碳同位素组成也与温度相关[14]。Krner等[17]发现植物的δ13C值随温度升高而降低,而Stuiver和Braziunas[18]的结果则显示温度与δ13C呈正相关。Lee等[19]和Feng等[20]通过对中国宝鸡至外蒙古北界断面的植物、土壤有机碳同位素组成分析,得出植物同位素组成与有效湿度之间有较好的相关性。尽管从现代植物样品的分析结果,植物碳同位素组成与降水量之间的相关性较为明显,而与温度变化不十分明显,但可以肯定的是在干旱-半干旱区降水和温度对植物碳同位素组成有一定的影响。而在降水量较大的地区,这种关系可能不十分明显。Schulze等[21]测定了不同生活型乔木的Δ值,在降雨量高于475 mm的地区,群落的平均Δ值基本相同;而在降雨量低于475 mm的地区,则从19‰下降至17‰ (220 mm)。说明在降雨量高的地区植物碳同位素组成变化不十分显著。
对干旱区植物碳同位素组成的分析表明,随着降水量(特别是有效湿度)的增加,碳同位素组成偏轻;相反,碳同位素组成偏重。这与植物对水分的利用状况一致。水分较多的情况下,叶片气孔开度较大,Ci值大,导致植物碳同位素组成偏轻。对于不同种属植物,尽管有相同的规律,但是因植物生理等因素的差异,碳同位素组成相差较大。同种植物之间碳同位素比率由于环境的差异(如水分、空气湿度、污染物等),其δ13C值可达3‰~5‰[22,23]。
虽然,叶片碳稳定同位素组成(δ13C)反映了环境变化对Ci/Ca的影响。但是当水分对植物生长并不是一个主要限制因子时,δ13C值受到蒸气压亏损(vapor pressure deficit, VPD)、光照、叶片含量等多个因子的影响[24]。因此,从我们对兰州市银杏叶片同位素组成的分析结果(图2)也可以看出,同位素组成与多个环境因素之间有不同程度的相关性。
2.3 大气污染物对植物叶片碳同位素组成的影响
从图2中,我们发现银杏叶片碳同位素组成与植物生长期兰州市大气污染指数(大气中氮氧化物、硫氧化物、大气颗粒物)之间的相关性较高(r=0.550),反映了大气污染物对植物碳同位素组成的影响。
O’leary[22]分析得出由于污染物、光照、蒸气压亏损等可造成植物碳同位素组成之间的差异。对兰州市叶片δ13C值与大气二氧化硫浓度的分析,得出δ13C值与大气二氧化硫浓度呈显著性正相关(r=0.828,P<0.01)[25]。Martin等[26]研究在不同SO2、O3、NOx浓度下生长的植物叶片δ13C值的变化情况,发现生长在污染物高浓度下叶片δ13C值要高于偏正清洁环境下的叶片δ13C值。说明随大气污染物浓度上升,植物碳同位素组成偏重。大气中的二氧化硫、臭氧、氮氧化物会降低叶片的气孔导度和光合速率。当大气中污染物浓度升高时,植物的稳定碳同位素分馏降低,Pi值减小进而使得Pi/Pa值减小,植物的δ13C值偏正。因此,在周围大气污染物浓度高的情况下,植物为了防止有害气体对自身的伤害而关闭部分气孔致使细胞内二氧化碳浓度降低,Pi减小,δ13C值偏正。
2.4 环境因素对植物叶片碳同位素组成的影响
由于植物碳同位素组成受到较多环境因素的影响,尤其是在城市中的植物不仅受到自然光照、降水量、蒸发量、温度等环境参数的控制,而且,由于城市中人类活动产生了大量的大气污染物,以及城市特殊的环境导致不同方向光照强度与自然环境中的差异,众多的原因使植物碳同位素组成的影响因素变得相对复杂。当对某一城市环境中植物碳同位素组成影响因素的了解,对城市生态系统的认识却有相当重要的价值。因此,我们通过对兰州市区校园中银杏叶片生长期中碳同位素组成与生长季中降水、温度、光照、大气污染物浓度的变化(转换为污染指数的变化)、气孔比表面积、气孔面积、气孔密度等环境因素的综合相关性分析,了解各环境因素对植物碳同位素组成的影响。
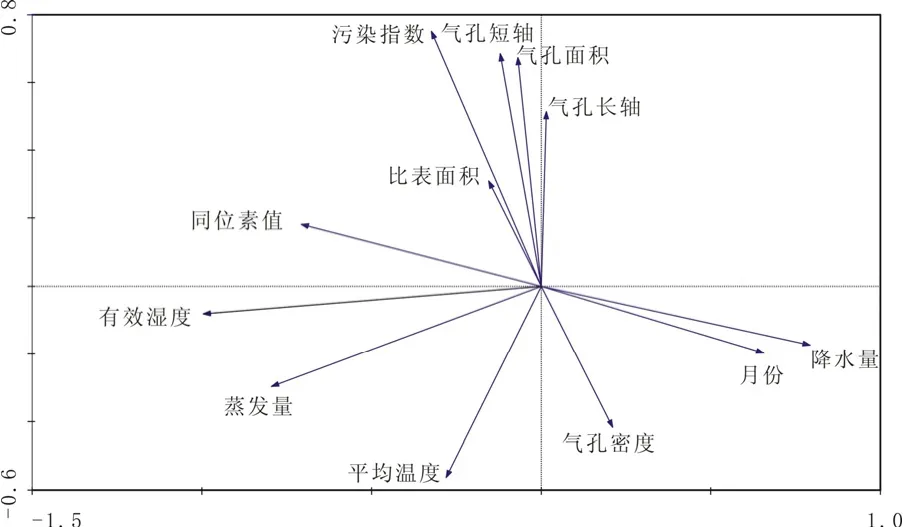
图4 兰州市银杏叶片生长期碳同位素组成与环境因素之间的相关性分析Fig.4 The relationships among environmental parameters and carbon isotope of Gingko leaves in the period of growing at Lanzhou
对于多指标的相关性分析,CANOCO分析软件是目前生物学中运用较广的一个数理统计软件。分析结果见图4。从图中可以明显反映出不同环境参数之间的相互关系,其中污染指数与叶片的气孔参数的相关性很好,说明了大气污染对植物叶片气孔的影响较为明显,这也反映了叶片碳同位素组成受大气污染物的影响。叶片同位素组成与降水量和有效湿度有较好的相关关系,而与温度的相关性相对较差。叶片气孔参数对叶片碳同位素组成有一定的影响,但是反映不出如Farquhar等[1]提出的植物叶片同位素组成受 Ci/Ca,或者 Pi/Pa的制约,而是受到不同的环境参数共同影响,并且这些环境参数之间也是相互影响,造成植物碳同位素组成随环境的差异有很大的不确定性。事实上,Leuning[27]通过长期的实验观察和分析发现,非常多的植物种属在不同的环境中Ci/Ca比值基本处于恒定状态。因此,植物同位素组成的变化则明显受其它因素的影响。
3 结论
影响植物碳同位素组成的环境因素较多,在环境参数越复杂的地区控制因素则越多。城市环境相对于自然环境来说,不仅气象因素对植物同位素组成有较大的影响,而且,城市环境中人为因素如大气污染物、城市建筑对植物受光的改变等均可以影响植物的碳同位素组成。兰州市校园中银杏各向叶片生长期的同位素组成受到降水、温度、光照、大气污染物浓度的变化(转换为污染指数的变化)、气孔比表面积、气孔面积、气孔密度等环境因素的综合影响,通过CANOCO统计分析表明,城市中大气污染物浓度的变化对植物同位素组成的影响较大。
致谢:本文受国家自然科学基金(40773064)资助。在实验样品采集和制备过程中,李建伟、张明镇和高贤良同学给予我极大的帮助,在此致以诚挚的谢意!
[1]FARQUHAR G D, OLEARY M H, BERRY J A.On the relationship between carbon isotope discrimination and intercellular carbon dioxide concentration in leaves[J].Australian Journal of Plant Physiology,1982, 9: 121-137.
[2]BEERLING D J.Carbon isotope discrimination and stomatal responses of maturePinus sylvestrisL.trees exposed in situ for three years to elevated CO2and temperature[J].Acta Oecologica, 1997, 18:697-712.
[3]BEERLING D J, CHALONER W G, MATTEY D P.Shifts in the δ13C composition ofSalix herbaceaeL.leaves in response to spatial and temporal gradients of atmospheric CO2concentration[J].Proceedings of the Royal Society of London Series B, 1993, 253: 53-60.
[4]VAN D E, WATER P K, LEAVITT S W, et al.Trends in stomatal density and the13C/12C ratios of Pinus-exilis needles during last glacial-interglacial cycle[J].Science, 1994, 264: 239-243.
[5]陈立群.银杏类植物的叶表皮特征及其气孔频度对大气 CO2浓度的指示[D].北京: 中国科学院机构知识仓储系统//中国科学院植物研究所博士学位论文, 2000: 37-38.CHEN Liqun.The epidermal characters of ginkgophytes and their stomatal frequencies as an indicator of atmospheric CO2[D].Knowledge Repository of CAS , PhD thesis Of Institute Of Botany , The Chinese Academy of Sciences , 2000: 37-38.
[6]LOCKHEART M J, VAN BERGEN P F, EVERSHED R P.Variations in the stable carbon isotope compositions of individual lipids from the leaves of modern angiosperms: implications for the study of higher land plant-derived sedimentary organic matter[J].Organic Geochemistry, 1997, 26: 137-153.
[7]LOCKHEART M J, POOLE I, VAN BERGEN P F, EVERSHED R P.Leaf carbon isotope compositions and stomatal characters: important considerations for palaeoclimate reconstructions[J].Org.Geochem,1998, 29(4): 1003-1008.
[8]FARQUHAR G D, EHLERINGER J R, HUBICK K T.Carbon isotope discrimination and photosynthesis[J].Annual Review of Plant Physiology and Plant Molecular Biology, 1989, 40: 503-537.
[9]DELUCIA E H, SCHLESINGER W H, BILLINGS S.Water relations and the maintenance of Sierran conifers on hydrothermally altered rock[J].Ecology, 1988, 69: 303-311.
[10]WARING R H, SILVESTER W B.Variation in foliar δ13C values with in tree crowns of Pinus radiata.Tree Physiology, 1994, 14: 1203-1213.
[11]KÜRSCHNER W M, VANDER B J, VISSCHER H, DILCHER D L.Oak leaves as biosensors of late Neogene and early Pleistocene paleoatmospheric CO2concentrations[J].Marine Micropaleontology,1996, 27: 299-312.
[12]POOLE I, WEYERS J D B, LAWSON T, RAVEN J A.Variations in stomatal density and index: implications for palaeoclimatic reconstructions[J].Plant Cell and Environment, 1996, 19: 705-712.
[13]VOGEL J C, EHLERINGER J R, HALL A E, et al.Variability of carbon isotope fractionation during photosynthesis.In Stable Isotopes and Plant Carbon-Water Relations[M].Academic Press, San Diego,CA, pp, 1993: 29-46.
[14]王国安, 韩家懋, 刘东生.中国北方黄土区 C-3草本植物碳同位素组成研究[J].中国科学: D辑, 2003, 33(6): 550-556.WANG Guoan, HAN Jiaxin, LIU Dongsheng.The study of C-3 herb isotopic composition in the northern part of loess area in China[J].Science in China: Series D, 2003, 33(6): 550-556.
[15]ZHANG Chengjun, CHEN Fahu, JIN Ming.Study on modern plant C-13 in the Western China and its significance[J].China Geochemical Acta, 2003, 22(2): 97-106.
[16]王国安, 韩家懋, 周力平.中国北方 C3植物碳同位素组成与年均温度关系[J].中国地质, 2002, 29 (1): 55-57.WANG Guoan, HAN Jiamao, ZHOU Liping.The annual average temperature in northern China[J].Geology in China, 2003, 33(6):550-556.
[17]KRNER C H, FARQUHAR G D, ROKSAUDIC Z.A global survey of carbon isotope discrimination in plants from high altitude[J].Oecologia, 1988, 74: 623-632.
[18]STUIVER M, BRAZIUNAS T F.Tree cellulose13C/12C isotope ratios and climatic change[J].Nature, 1987, 328: 58-60.
[19]LEE X Q, FENG Z D, GUO L L, et al.Carbon isotope of bulk organic matter: A proxy for precipitation in the arid and semiarid central East Asia[J].Global Biogeochemical Cycles, 2005, 19(4): GB4010.1-GB4010.8.
[20]FENG Z D, WANG L X, JI Y H, et al.Climatic dependency of soil organic carbon isotopic composition along the S-N Transect from 34°N to 52°N in central-east Asia[J].Palaeogeography, Palaeoclimatology, Palaeoecology, 2008, 257: 335-343.
[21]SCHULZE E D, WILLIAMS R J , FARQUHAR G D, et al.Carbon and nitrogen isotope discrimination and nitrogen nutrition of trees along a rainfall gradient in northern Australia[J].Australian Journal of Plant Physiology , 1998, 25 : 413-425.
[22]O’LEARY M H.Carbon isotopes in photo synthesis[J].Biosci, 1988,38 (5): 328-336.
[23]SUN Bainian, DILCHER D L, BEERLING D J, et al.Variation in Ginkgo biloba L.leaf characters across a climatic gradient in China[J].PANS, 2003, 100(2): 7141-7146.
[24]HAMERLYNCK E P, HUXMAN T E, MCAULIFFE J R, et al.Carbon isotope discrimination and foliar nutrient status of Larreatri dentata (creosote bush) in contrasting Mojave Desert soils[J].Oecologia,2004, 138: 210-215.
[25]黄雅娟, 张成君, 殷树鹏, 等.兰州市大气中 SO2对植物碳同位素组成的影响[J].安徽农业科学, 2008, 36(34): 15170-15171.HUANG Yajuan, ZHANG Chengjun, YIN Shupeng, et al.Effects of Atmospheric SO2on Carbon Isotope Composition of Plants in Lanzhou City.Journal of anhui agricultural sciences, 2008, 36(34):15170-15171.
[26]MARTIN B, BYTNEROWICZ A, THORSTENSON Y R.Effects of air pollutants on the composition of stable carbon isotopes, δ13C, of leaves and wood, and on leaf injury[J].Plant Physiol, 1988, 88:218-223.
[27]LEUNING R.A critical appraisal of a combined stomata-photosynthesis model for C3 plants[J].Plant Cell and Environment, 1995, 18: 339-355.
