紫茎泽兰(Eupatorium adenophorum)入侵地的生物多样性
2010-07-13张修玉许振成宋巍巍黎忠赵晓光胡习邦
张修玉 ,许振成,宋巍巍,黎忠,赵晓光,胡习邦
1.环境保护部华南环境科学研究所,广东 广州 510655;2.中国科学院广州地球化学研究所,广东 广州 510640;3.中水珠江规划勘测设计有限公司水环境与生态修复研究所,广东 广州 510610
紫茎泽兰(Eupatorium adenophorum)是菊科(Compositae)泽兰属(Eupatorium)的多年生丛生型半灌木状草本植物,原产于中美洲的墨西哥与哥斯达黎加等国[1],19世纪曾作为观赏植物引种到欧洲,后来引种到澳洲和亚洲,是一种抗逆性强、生态适应广、传播速度快且群体自然繁殖与演替能力极强的植物[2]。上世纪40年代从中缅边境传入西双版纳后,以平均每年20 km的速度向东和向北快速蔓延扩散[3],目前其入侵面积超过3000万hm2[4],是我国外来入侵物种中危害最为严重的植物之一,已被列入我国公布的第1批外来入侵物种名单之首。物种入侵导致的生物多样性丧失被看作是全球变化的重要内容之一[5],研究紫茎泽兰入侵对生物多样性与生态系统功能的影响具有重要的意义。
由于紫茎泽兰能够通过分泌化感物质[6-8],抑制其它植物的生长发育,因此在热带、亚热带自然森林植被遭受严重破坏和土地利用方式不合理的地区,很容易被紫茎泽兰所侵占,从而给当地物种组成和多样性的演替造成一定的影响。目前对紫茎泽兰入侵地生物多样性的研究大多集中在植物部分,对入侵地土壤动物和土壤微生物的多样性研究较少。本文拟通过对西双版纳紫茎泽兰不同入侵程度样地的植物、土壤动物与土壤微生物的多样性进行立体、全面、系统的对比分析,探讨西双版纳紫茎泽兰入侵对生物多样性的影响,旨在为揭示紫茎泽兰入侵后的物种演替趋势提供科学依据,并为紫茎泽兰迹地植被的恢复重建与后续管理提供理论参考。
1 研究地区概况
研究地点位于西双版纳勐海县格朗和哈尼族乡雷达山。研究区地带性植被属季节性雨林,但由于人类活动的影响,研究样地原生植被已不复存在。目前植被群落高度约1.3 m,总盖度约95%,主要物种包括紫茎泽兰(Eupatorium adenophorum)、野菊(Dendranthema indicum)、马唐(Digitaria sanguinalis)、鬼针草 (Bidens bipinnata)、狗尾草(Setaria viridis)、山莓(Rubus corchorifolius)、菝葜(Smilax china)、芒箕(Dicranopteris linearis)、五节芒(Miscanthus floridulu)、乌毛蕨(Blechnum orientale)、胜红蓟(Ageratum conyzoides)等。研究区属于热带、亚热带西南季风气候,年降雨量为1418.2 mm,土壤类型为赤红壤。
2 研究方法
2.1 紫茎泽兰入侵程度类型划分
由于研究区域原来分布有不同的植被且被破坏程度不同,导致紫茎泽兰的入侵程度也不尽相同,出现不同的紫茎泽兰群落演替阶段。有的区域已形成紫茎泽兰单优种群,有的区域是紫茎泽兰和当地植物共同竞争生长,也有当地植物占优势而紫茎泽兰仅为零星分布的区域和几乎无植物生长的裸露地区域。在研究区内根据紫茎泽兰和当地植物群落竞争演替的不同阶段,以及紫茎泽兰盖度和大致入侵时间的不同,参考牛红榜等人[9]的方法划分为重度入侵区、中度入侵区和轻度入侵区3个入侵类型。在本研究中,紫茎泽兰为绝对优势种群,占草本植物层总盖度95%以上的为重度入侵区;紫茎泽兰为优势种群,有部分本地植物,紫茎泽兰盖度占草本植物层总盖度 55%~75%的为中度入侵区;紫茎泽兰与当地植物处于竞争生长,种群优势不明显,紫茎泽兰盖度占草本植物层总盖度低于35%的为轻度入侵区。
2.2 调查、取样与试验方法
2.2.1 植物调查方法
在 3种不同的入侵类型样区各设置一个 50 m×50 m的大样方,为了减少群落交错区的边缘效应,在每个大样方内,从距离四周边线10 m处起,在30 m×30 m范围内按照对角线方向各设置5个5 m×5 m的小样方,共计设15个的小样方,对小样方内的物种、株数、高度、盖度等指标进行登记。
2.2.2 土壤动物调查与取样方法
在15个5 m×5 m的小样方内,随机设置两个样点,一个为1 m×1 m,在其样方内按0~5,5~10,10~15,15~20 cm的土壤连续分四层取样,手拣法获取大型动物;另一个样方设为0.3 m×0.3 m,以100 cm3土壤容重器的土体为一个土样,分 4层(0~5、5~10、10~15、15~20 cm)均匀取样,每层取4个土样带回室内,3个土样用Tullgren法(干漏斗法),另一土样的 1/4用 Baermunn法(湿漏斗法)分离提取中小型土壤动物。所有获取的土壤动物标本用75%酒精保存,然后根据尹文英《中国亚热带土壤动物》和《中国土壤动物检索图鉴》进行分类鉴定[10-11]。
2.2.3 土壤微生物调查与取样方法
在每个5 m×5 m的小样方内,随机设置3个样点,从3个样点各取0~20 cm的土层土壤样品约500 g,充分混合后,用灭菌封口袋装回实验室,每袋土壤样品1500 g,共15袋样品,放在4 ℃的冰箱中保存备用,土壤细菌、真菌、放线菌数量分别采用牛肉膏蛋白胨培养基、马丁氏培养基和改良高氏1号培养基培养,自生固氮菌用Ashby无氮培养基选择性培养,平板计数法测定;反硝化细菌和氨氧化细菌采用最大或然计数法(MPN)测定[12]。每种入侵类型土壤5个混合土样,每一个土样在涂平板时设置5个稀释梯度,每个浓度设置3个重复。
2.3 数据分析方法
2.3.1 植物部分数据分析方法
主要物种重要值(I)计算公式为:I=Rc+Rd+Rf,式中Rc为相对盖度(%),Rd为相对密度(%),Rf为相对频度(%)。其中,Rc为(第i物种的盖度/所有物种的盖度和)×100,Rd为(第i物种的密度/所有物种的密度和)×100,Rf为(第i物种的频度/所有物种的频度和)×100。
物种丰富度指数、多样性指数、均匀度指数和优势度指数作为植物多样性的测度指数[13],公式见表1。

表1 生物多样性评价指标Table 1 Evaluating indicator of biodiversity
2.3.2 土壤动物部分数据分析方法
动物频度统计方法与植物相同,多度按照如下方法进行统计[14]:某类个体数量占总捕获量10.00%以上者为优势类群,占 1.00%~10.00%者为常见类群,不足1.00%者为稀有类群。
物种多样性指数、均匀度指数和优势度指数计算公式见表1。密度类群指数[15]按如下公式计算:
DG=(g/G)∑(DiCi/DimaxC)
其中g为群落中的类群数,G为总类群数;Di为第i类群的密度,Dimax为各类群中第i类群的最大密度;C为研究的群落数,Ci为第i个类群在C个群落中出现的次数。
2.3.3 土壤微生物部分数据分析方法
在本研究中,我们把微生物类群作为物种数,数量作为个体数,物种多样性、优势度与均匀度指数计算公式见表1。
2.3.4 统计分析方法
本研究所有数据统计分析前,对数据进行了正态分布检验,对不符合正态分布的数据进行了对数转换,然后利用SPSS16.0系统软件进行方差分析。
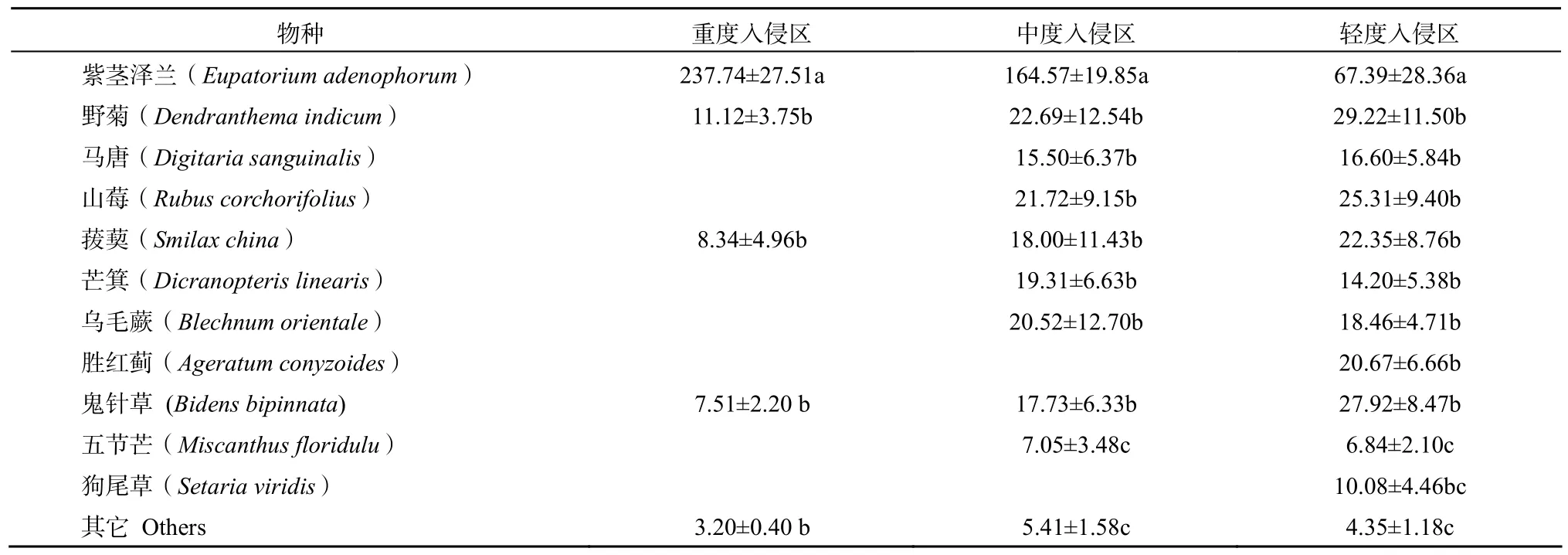
表2 不同入侵类型的植物物种及其重要值Table 2 Plant species and important value of different invading types %
3 结果与分析
3.1 植物多样性
调查结果表明(见表2),紫茎泽兰不同程度入侵区的植物种类及其重要值差别较大。在重度入侵区,除紫茎泽兰外,仅零星分布野菊、菝葜和鬼针草等少数几种植物,紫茎泽兰的重要值高达237.74%,而其它物种的重要值均不足 12%,且与紫茎泽兰的重要值相比,差异显著。在中度入侵区与轻度入侵区,植物种类数均有所增加,表现为轻度入侵区>中度入侵区>重度入侵区;同时,紫茎泽兰的重要值明显降低,且野菊、菝葜和鬼针草等其它物种的重要值却随入侵程度的降低而升高,但与紫茎泽兰的重要值相比,差异依旧显著。紫茎泽兰不同入侵程度的植物多样性方面(见表3),丰富度指数、多样性指数、优势度指数与均匀度指数均表现为轻度入侵区>中度入侵区>重度入侵区。与重度入侵区相比,中度入侵区的多样性指数差异显著,丰富度指数、优势度指数与均匀度指数差异不显著;轻度入侵区的优势度指数与均匀度指数差异显著,丰富度指数与多样性指数差异极显著。

表3 不同入侵类型的植物多样性Table 3 Plant biodiversity of different invading types
3.2 土壤动物多样性
对紫茎泽兰不同入侵程度类型的土壤动物种类鉴定发现(见表4),三种入侵区的土壤动物分别隶属 23目,其中弹尾目、蜱螨目和膜翅目土壤动物为优势种群,三者总量占捕获总数的61.77%;双尾目、鞘翅目、综合目、双翅目、园马陆目与地蜈蚣目土壤动物为常见类群,占捕获总量的31.89%;革翅目、缨翅目、直翅目、鳞翅目、等翅目、等足目、蜘蛛目、半翅目、蚯蚓目、原尾目、蜚蠊目、端足目、石蜈蚣目与拟蝎目为稀有类群,仅占捕获总量的6.34%。

表4 不同入侵类型的土壤动物数量与种类Table 4 Soil animal quantity and category of different invading types
紫茎泽兰不同入侵程度的土壤动物多样性方面(见表5),丰富度指数、多样性指数、优势度指数、均匀度指数与密度类群指数均表现为轻度入侵区>中度入侵区>重度入侵区。但与重度入侵区相比,中度入侵区的丰富度指数、多样性指数、优势度指数、均匀度指数与密度类群指数差异均不显著;轻度入侵区的丰富度指数、多样性指数、优势度指数与均匀度指数差异不显著,只有密度类群指数差异显著。

表5 不同入侵类型的土壤动物多样性Table 5 Soil animal biodiversity of different invading types
3.3 土壤微生物多样性
三种不同入侵程度的土壤微生物可培养数量类群方面,细菌表现为中度入侵区>重度入侵区>轻度入侵区,且三者之间无显著差异;放线菌、真菌、自生固氮菌与氨氧化细菌均表现为重度入侵区>中度入侵区>轻度入侵区,与重度入侵区相比,中度入侵区与轻度入侵区的放线菌和真菌均表现为差异显著,但中度入侵区与轻度入侵区的氨氧化细菌无差异显著,而自生固氮菌只有轻度入侵区差异显著;反硝化细菌表现为轻度入侵区>中度入侵区>重度入侵区,且三者之间无显著差异。
紫茎泽兰不同入侵程度的土壤微生物多样性方面(见表7),多样性指数表现为重度入侵区>轻度入侵区>中度入侵区,且与重度入侵区相比,轻度入侵区与中度入侵区均表现为差异显著;优势度指数表现为轻度入侵区=中度入侵区>重度入侵区,且三者之间无差异显著;均匀度指数表现为重度入侵区>轻度入侵区=中度入侵区,且三者之间无差异显著。
4 讨论
4.1 植物多样性
紫茎泽兰对入侵地植物多样性有明显的降低作用,本研究中4种多样性指数均表现为轻度入侵区>中度入侵区>重度入侵区,即随入侵程度的增强,植物多样性降低,这与丁晖等人[16]的研究结果一致。究其原因,可能与紫茎泽兰的化感作用有关[6-7,17]。研究表明[18],化感作用是植物通过分泌化学物质并向环境中释放,从而对其他植物的生长产生抑止作用,最终影响植物群落的分布与演化。谢明惠等人[19]也研究证明,紫茎泽兰根系释放的化感物质是导致其土壤环境发生变化的重要因素,可为其成功入侵提供有利条件。

表6 土壤微生物可培养类群Table 6 quantity of cultivable soil microbe groups g-1

表7 不同入侵类型的土壤微生物多样性Table 7 Soil microbial biodiversity of different invading types
另外,紫茎泽兰入侵地区土壤种子库变化与植物根系特征也会影响植物多样性。党伟光等[20]研究发现,紫茎泽兰在其入侵地土壤种子库中能迅速占有绝对优势,这导致其他物种从种子阶段就开始亏缺,从而最终影响了地上部植物多样性;于亮等[21]则研究证明,在重度入侵区,紫茎泽兰根系系统发达程度显著高于中度和轻度入侵区域。
4.2 土壤动物多样性
紫茎泽兰对入侵地土壤动物多样性也有一定的降低作用,但这种影响已远低于对植物多样性的影响,与重度入侵区相比,只有轻度入侵区的密度类群指数差异显著,轻度入侵区的其它多样性指数与中度入侵区的各多样性指数均为差异不显著。刘志磊等人[22]也研究发现,紫茎泽兰入侵后土壤动物类群总数与群落个体总数均有所减少,减少程度表现为针叶林>草地>阔叶林。王文琪等人[23]对撂荒地、林区和农林交错带3种不同生境紫茎泽兰入侵地土壤节肢动物群落的组成与结构研究表明,农林交错地和林地物种数与个数减少程度大于撂荒地且差异显著。其它物种入侵方面,Belnap和Phillips[24]研究表明,旱雀麦入侵美国犹他州导致了土壤无脊椎动物数量的降低;Ratsirarson等[25]则发现南非开普半岛国家公园中入侵植物组成的桉树园和松树园中膜翅目(Hymenoptera)、盲蛛目(Opiliones)和端足目(Amphipoda)3类无脊椎动物物种数低于土著植物林。
然而,并不是所有的物种入侵都会降低土壤动物的多样性,Kourtev等[26]与Aplet[27]研究发现,火树(Myrica faya)等物种入侵反而能使土壤中蚯蚓数量明显增多。Gremmen等[28]比较了新西兰次南极区群岛外来植物群落与土著植物群落的土壤动物,发现前者的大型无脊椎动物和螨类物种多于后者。Porazinska等[29]调查了入侵北美的须芒草属植物(Andropogo bladhii)和土著植物土壤中线虫的多样性,发现虽然多样性没有显著差异,但入侵植物土壤中线虫的密度有所增加。Posey等[30]则研究证明,入侵美国东海岸的芦苇与土著植物互花米草土壤中的大型动物之间无明显差异。可见,紫茎泽兰入侵对土壤动物多样性影响的机制方面,有可能是先通过化感作用抑止地上部植物的生长,导致地上部分的初级生产力和植物组织化学特性发生改变,再通过分解作用进入碎屑食物网的资源产生质量的不同,进而影响土壤动物的组成和多样性。但紫茎泽兰的化感作用是否会直接影响地下部土壤动物的多样性分布还有待长期野外的追踪调查。
4.3 土壤微生物多样性
紫茎泽兰对入侵地土壤微生物多样性的影响表现为多样性指数与均匀度指数为重度入侵区最高,而其优势度指数却低于中度入侵区与轻度入侵区。诸多研究证明[31-32],紫茎泽兰入侵能通过调节土壤有机质库,提高土壤呼吸强度与土壤肥力,增加了有效钾、土壤脲酶、磷酸酶和蔗糖酶等物质的含量,导致入侵地土壤微生物群落的互惠互利,从而进一步改变土壤微生物群落结构,增加了土壤微生物的多样性,特别是增加了与土壤养分循环相关的微生物功能类群数量,进而提高了土壤可利用的养分水平,创造了对自身生长有利的土壤环境,并最终形成对自身生长有利的微生态环境以利于入侵扩张。
Duda等人[33]研究表明,美国西部的入侵植物盐生草(Halogeton glomeratus)的土壤细菌功能多样性显著高于土著植物。Saggar等人[34]比较了入侵新西兰的外来植物绿毛山柳菊(Hieracium pilosella)与土著植物群落土壤中微生物的生物量,结果发现绿毛山柳菊显著增加了土壤微生物的生物量。入侵植物之所以能改变其入侵地土壤微生物的多样性,究其原因,有可能是外来植物入侵后会通过根系分泌不同的化学物质改变根系对土壤的物质输入,影响土壤微生物的结构与功能[35]。但另有研究证明,入侵植物的扩散和蔓延依赖于特有的土壤真核微生物群落结构模式[36]。因此,有关紫茎泽兰入侵对土壤微生物多样性的影响机制,还需要通过室内模拟试验进一步深入研究。
5 结论
(1)不同程度入侵区的植物种类及其重要值差别较大,在重度入侵区,紫茎泽兰的重要值高达237.74%,但随着入侵程度的降低,植物种类有所增加且紫茎泽兰的重要值随之降低;丰富度指数、多样性指数、优势度指数与均匀度指数均表现为轻度入侵区>中度入侵区>重度入侵区。
(2)三种入侵区的土壤动物隶属 23目,其中弹尾目、蜱螨目和膜翅目为优势种群,三者总量占捕获总数的61.77%;丰富度指数、多样性指数、优势度指数、均匀度指数与密度类群指数均表现为轻度入侵区>中度入侵区>重度入侵区。
(3)土壤微生物可培养数量类群方面,细菌表现为中度入侵区>重度入侵区>轻度入侵区,放线菌、真菌、自生固氮菌与氨氧化细菌均表现为重度入侵区>中度入侵区>轻度入侵区;多样性指数表现为重度入侵区>轻度入侵区>中度入侵区,优势度指数表现为轻度入侵区=中度入侵区>重度入侵区,均匀度指数表现为重度入侵区>轻度入侵区=中度入侵区。
[1]徐海根, 王健民, 强胜, 等.生物多样性公约热点研究: 外来物种入侵、生物安全、遗传资源[M].北京:科学出版社, 2004, 24.XU Haigen, WANG Jianmin, QIANG Sheng, et al.Popular study of Convention on Biological Diversity: biotic intrusion of exotic species,bio-safety and genetic resources[M].Beijing: Science Press, 2004, 24.
[2]李爱芳, 高贤明, 党伟光, 等.不同生境条件下紫茎泽兰幼苗生长动态[J].生物多样性, 2007, 15(5): 479-485.LI Aifang, GAO Xianming, DANG Weiguang, et al.Growth trends of Crofton weed (Eupatorium adenophorum) seedlings in different habitats[J].Biodiversity Science, 2007, 15(5):479-485.
[3]WANG R, WANG Y Z.Invasion dynamics and potential spread of the invasive alien plant speciesAgemtina adenophom(Asteraceae) in China[J].Diversity and Distributions, 2006, 12(4): 397-408.
[4]GUI F R, Wan F H, Guo J Y.Population genetics ofAgemtina adenophomusing inter-simple sequence repeat(ISSR) molecular markers in China[J].Plant Biosystems, 2008, 142(2): 255-263.
[5]黄艺, 郑维爽.《生物多样性公约》国际履约过程变化分析[J].生物多样性, 2009, 17(1): 97-105.HUANG Yi, ZHENG Weishuang.Dynamics in international implementation of the Convention on Biological diversity[J].Biodiversity Science, 2009, 17 (1): 97-105.
[6]王俊峰, 冯玉龙.人工群落中苗期紫茎泽兰的化感作用和对光环境的适应[J].生态学报, 2006, 26(6): 1809-1817.WANG Junfeng, FENG Yulong.Allelopathy and light acclimation characteristic forAgeratina adenophoraseedling grown in man-made communities[J].Acta Ecologica Sinica, 2006, 26(6): 1809-1817.
[7]李渊博, 徐晗, 石雷, 等.紫茎泽兰对五种苦苣苔科植物化感作用的初步研究[J].生物多样性, 2007, 15(5): 486-491.LI Yuanbo, XU Han, SHI Lei, et al.Allelopathic effects ofEupatorium adenophorumon five species of the family Gesneriaceae[J].Biodiversity Science, 2007, 15(5): 486-491.
[8]韩利红, 冯玉龙.发育时期对紫茎泽兰化感作用的影响[J].生态学报, 2007, 27(3): 1185-1191.HAN Lihong, FENG Yulong.The effects of growth and development stage on allelopathy ofEupatorium adenophorum[J].Acta Ecologica Sinica, 2007, 27(3):1185-1191.
[9]牛红榜, 刘万学, 万方浩.紫茎泽兰入侵对土壤微生物群落和理化性质的影响[J].生态学报, 2007, 27(7): 3051-3060.NIU Hongbang, LIU Wanxue, WAN Fanghao.Invasive effects ofAgeratina adenophoraSprengel(Asteraceae) on soil microbial community and physical and chemical properties[J].Acta Ecologica Sinica,2007, 27(7): 3051-3060.
[10]尹文英.中国亚热带土壤动物[M].北京: 科学出版社, 1992.YIN Wenying.Subtropical soil animal in China[M].Beijing: Science Press, 1992.
[11]尹文英.中国土壤动物检索图鉴[M].北京:科学出版社, 1998.YIN Wenying.Illustrated handbook of searching soil animal in China[M].Beijing: Science Press, 1998.
[12]赵斌, 何绍江.微生物学实验[M].北京:科学出版社, 2003: 69-75.ZHAO Bin, HE Shaojiang.Microbiology experiment[M].Beijing:Science Press, 2003: 69-75.
[13]马克平.生物多样性研究的原理与方法[M].北京: 科学出版社,1994: 1-237.MA Keping.Principles and Methods of researching biodiversity[M].Beijing: Science Press, 1994: 1-237.
[14]刘红, 袁兴中.曲阜孔林土壤动物多样性研究[J].应用生态学报,1999, 10(5): 609-611.LIU Hong, YUAN Xingzhong.Diversity of soil animals in Confucian graveyard of Qufu[J].Chinese Journal of applied ecology, 1999, 10(5):609-611.
[15]廖崇惠, 李健雄.再论DG指数的性质与应用[J].生物多样性, 2009,17 (2): 127-134.LIAO Chonghui, LI Jianxiong.Re-evaluating the character and application of density-group index (DG)[J].Biodiversity Science, 2009, 17(2): 127-134.
[16]丁晖, 徐海根, 刘志磊.外来入侵植物紫茎泽兰对植物多样性的影响[J].生态与农村环境学报, 2007, 23(2):29-32.DING Hui, XU Haigen, LIU Zhilei.Impacts of invasion ofEupatorium adenophorumon vegetation diversity[J].Journal of Ecology and Rural Environment, 2007, 23(2):29-32.
[17]郑丽, 冯玉龙.紫茎泽兰叶片化感作用对 10种草本植物种子萌发和幼苗生长的影响[J].生态学报, 2005, 25(10): 2782-2787.ZHENG Li, FENG Yulong.Allelopathic effects ofEupatorium adenophorumon seed germination and seedling growth in ten herbaceous species[J].Acta Ecologica Sinica, 2005, 25(10): 2782-2787.
[18]张开梅, 石雷, 李振宇.蕨类植物的化感作用及其对生物多样性的影响[J].生物多样性, 2004, 12(4): 466-471.ZHANG Kaimei, SHI Lei, LI Zhenyu.Fern allelopathy and its impact on biodiversity[J].Biodiversity Science, 2004, 12(4): 466-471.
[19]谢明惠, 任琴, 张青文, 等.紫茎泽兰根区土壤酚酸类物质组成及其对土传病菌的影响[J].应用生态学报, 2010, 21(2): 306-311.XIE Minghui, REN Qin, ZHANG Qingwen, et al.Composition of phenolic allelochemicals inEupatorium adenophorumroot zone soils and its effects on soil-borne pathogen[J].Chinese Journal of Applied Ecology, 2010, 21(2):306-311.
[20]党伟光, 高贤明, 王瑾芳.紫茎泽兰入侵地区土壤种子库特征[J].生物多样性, 2008, 16 (2): 126-132.DANG Weiguang, GAO Xianming, WAGN Jinfang.Soil seed bank traits in an area invaded byEupatorium adenophorum[J].Biodiversity Science, 2008, 16(2): 126-132.
[21]于亮, 桂富荣, 李隽, 等.不同入侵程度紫茎泽兰群落的根系分布特征[J].西南农业学报, 2009, 2(5): 1444-1448.YU Liang, GUI Furong, LI Juan, et al.Distribution characteristics ofAgeratina adenophora(Compositae) roots in different invaded areas[J].Southwest China Journal of Agricultural Sciences, 2009, 2(5):1444-1448.
[22]刘志磊, 徐海根, 丁晖.外来入侵植物紫茎泽兰对昆明地区土壤动物群落的影响[J].生态与农村环境学报, 2006, 22(2): 31-35.LIU Zhilei, XU Haigen, DING Hui.Impacts of invasive alien plantEupatorium adenophorumon soil animal communities in Kunming[J].Journal of Ecology and Rural Environment, 2006, 22(2): 31-35.
[23]王文琪, 王进军, 赵志模, 等.不同生境紫茎泽兰对生态群落中节肢动物多样性的影响[J].西南大学学报: 自然科学版, 2009, 31(12):14-20.WANG Wenqi, WANG Jinjun, ZHAO Zhimo, et al.Effects of Different Habitats ofEupatorium adenophorumSpreng on the Biodiversity Arthropods[J].Journal of Southwest University: Natural Science Edition, 2009, 31(12): 14-20.
[24]BELNAP J, PHILLIPS S L.Soil biota in an ungrazed grassland: response to annual grass (Bromus tectorum) invasion[J].Ecological Applications, 2001, 11: 1261-1275.
[25]RATSIRARSON H, ROBETSON H G, PICKER M D, et al.Indigenous forests versus exotic eucalypt and pine plantations: a comparison of leaf-litter invertebrate communities[J].African Entomology, 2002, 10:93-99.
[26]KOURTEV P S, HUANG W Z, EHRENFELD J G.Differences in earthworm densities and nitrogen dynamics in soils under exotic and native plant species[J].Biological Invasions, 1999, 1: 237-245.
[27]APLET G H.Alteration of earthworm community biomass by the alien Myrica faya in Hawaii[J].Oecologia, 1990, 82: 414-416.
[28]GREMMEN N, CHOWN S L, MARSHALL D J.Impact of the introduced grassAgrostis stoloniferaon vegetation and soil fauna communities at Marion Island, sub-Antarctic[J].Biological Conservation,1998, 85: 223-231.
[29]PORAZINSKA D L, BARDGETT R D, BLAAUW M B, et al.Relationships at the aboveground-belowground interface: plants, soil biota and soil processes[J].Ecological Monographs, 2003, 73: 377-395.
[30]POSEY M H, ALPHIN T D, MEYER D L, et al.Benthic communities of common reed Phragmites australis and marsh cordgrass Spartina alterniflora marshes in Chesapeake Bay[J].Marine Ecology Progress Series, 2003, 261: 51-61.
[31]蒋智林, 刘万学, 万方浩, 等.紫茎泽兰入侵对土壤肥力特征的影响及其动态研究[J].农业环境科学学报, 2008, 27(1): 0267-0272.JIANG Zhiling, LIU Wanxue, WAN Fanghao, et al.Effects ofAgeratina adenophoraInvasion on Soil Nutrient Properties and Their Seasonal Dynamics[J].Journal of Agro-Environment Science, 2008, 27(1):0267-0272.
[32]刘潮, 冯玉龙, 田耀华.紫茎泽兰入侵对土壤酶活性和理化因子的影响[J].植物研究, 2007, 27(6): 729-735.LIU Chao, FENG Yulong, TIAN Yaohua.Effects ofEupatorium adenophorumSprengel invasionon Soil Enzyme Activities and Physical and Chemical Factors[J].Bulletin of Botanical Research, 2007, 27(6):729-735.
[33]DUDA J J, FREEMAN D C, EMLEN J M, et al.Differences in native soil ecology associated with invasion of the exotic annual chenopod,Halogeton glomeratus[J].Biology and Fertility of Soils, 2003, 38:72-77.
[34]SAGGAR S, MCINTOSH P D, HEDLEY C B, et al.Changes in soil microbial biomass, metabolic quotient, and organic matter turnover under Hieracium (H.pilosellaL.)[J].Biology and Fertility of Soils,1999, 30: 232-238.
[35]KOURTEY P S, HUANG W Z, EHRENFELD J G.Differences in earthworm densities and nitrogen dynamics in soils under exotic and native plant species[J].Biological Invasions, 1999, 1: 237-245.
[36]祖元刚, 高崇洋, 王文杰, 等.喜树替代紫茎泽兰过程中根际微生物群落特征[J].中国科学: C辑, 2006, 36 (5): 459-467.ZU Yuangang, GAO Chongyang, WANG Wenjie, et al.Biotic community characteristics of rhizospheric microorganism in the process of Camplotheca Acuminata replacingEupatorium adenophorum[J].Science in China: Series C, 2006, 36 (5): 459-467.
