声音在耳内的信号转导及其分子生物学机制(2)
2010-01-25汤浩石丽娟于利曹宇
汤浩 石丽娟 于利 曹宇
1.3 耳蜗的机械-电转换机制
1.3.1柯蒂器的机械-电转换作用 柯蒂器是耳蜗内的声音感受器,包括毛细胞和各种支持细胞,它们乘载于基底膜之上,并沿着基底膜的长轴进行延伸。柯蒂器中的大多数细胞功能不清,但已明确内、外毛细胞功能主要是感受声音刺激的。基底膜本身的机械特性及其与毛细胞、听神经的有序组合排列,使基底膜的任何位置都对应某一特定频率,即与此频率对应位置上的毛细胞对此频率是最敏感的,这些频率从蜗顶到蜗底按对数级排列。
1.3.1.1纤毛束的机械偏转诱发毛细胞兴奋
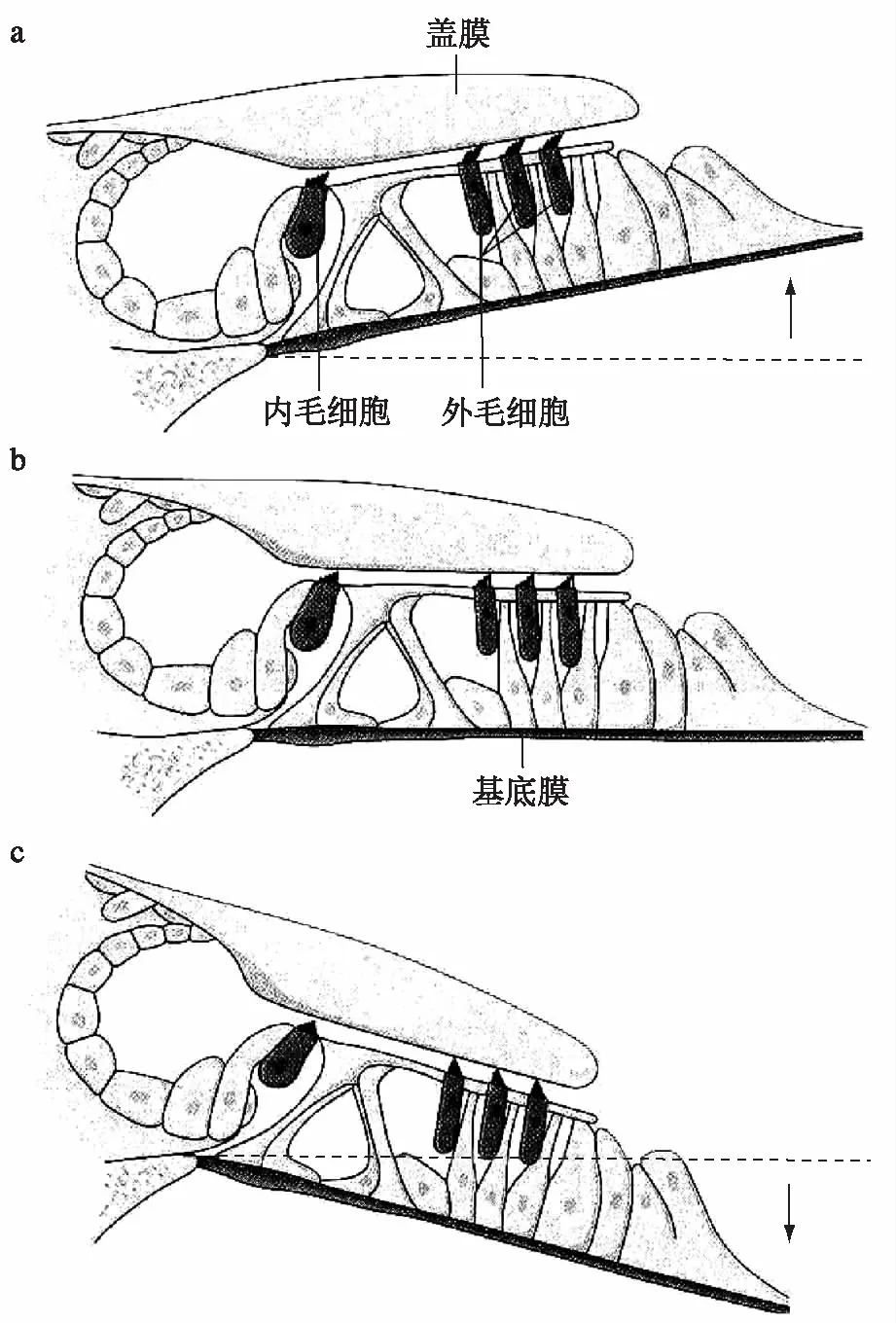
图2基底膜和盖膜振动时毛细胞顶部听纤毛受力情况示意图
a 表示基底膜向上偏移时听纤毛受到一个向外(动毛侧)的剪切力偏斜于引起毛细胞兴奋的方向
b 为基底膜处于振动的中点时纤毛束重新回到休止的位置
c 表示基底膜向下偏移时听纤毛则弯向内侧(与a的方向相反),将引起毛细胞超极化(引自Miller&Towe,1979)
如图2所示,外毛细胞顶端的听毛有些埋植于盖膜的胶状物中,有的则与盖膜的下面相接触,由于基底膜与盖膜的附着点不在同一个轴上,故当行波引起基底膜振动时,盖膜与基底膜便各自沿不同的轴上下移动,于是在两膜之间发生交错式移行运动,使毛细胞的听纤毛受到一个剪切力的作用而发生弯曲或偏转;内毛细胞的纤毛较短,不与盖膜接触,呈游离状态,主要由内淋巴的运动使其弯曲或偏转。当基底膜向上偏移时,听纤毛束弯向动毛侧,即偏斜于引起细胞兴奋的方向(图2a),可使毛细胞产生去极化电位,引起递质释放[3~5],其递质一般认为是谷氨酸类物质,其受体属于NMDA型,该递质使突触后膜(听神经纤维末梢)产生局部电位(类似兴奋性突触后电位),该电位达到阈电位时,则产生动作电位即传入性神经冲动。反之,当基底膜下移时听纤毛弯向内侧(图2c),则产生超极化电位,神经冲动发放减少。图2b则表示基底膜处在振动的中点,纤毛束重新回到休止的位置。

图3 耳蜗毛细胞声音调谐曲线
上图是简化的耳蜗及基底膜模式图,下图为频率-刺激强度曲线。从A、B、C、D四个声调曲线形状上看,都有一个向下的尖峰,表明相对应的频率反应的阈值(最小刺激强度)。图上面的A、B、C、D分别表示基底膜上相对应的毛细胞都对一个特定频率最为敏感,这个频率就是细胞的特征性频率(或最适频率)。 当刺激频率升高或降低时,阈值明显升高(敏感性突然降低) (引自Pickles 1988)
1.3.1.2毛细胞兴奋与刺激频率的相关性 为进一步证明毛细胞兴奋与刺激频率的关系,Pickles[6]用改变刺激频率和刺激强度建立一个曲线,用A、B、C、D四个频率分别刺激耳蜗,在每个频率上都逐渐增加刺激的声强,直到产生一个符合标准的反应(在这里定为1 mV)。因此这个曲线在一定的频率范围内反映了毛细胞对于刺激的反应阈值(曲线向下尖端A、B、C、D)。图3中可见,当刺激频率升高或降低时,阈值明显升高(敏感性突然降低)。基底膜上不同部位的毛细胞反应阈值即代表它的最适频率,该频率两侧(或高或低)的频率刺激,刺激声强在逐渐升高,表明其敏感性在降低。实验证明,耳蜗不仅对声音频率的选择性非常敏锐,而且具有声能的机械放大作用。
近年来利用细胞电压钳和膜片钳技术对毛细胞的感受器电位进行了深入的研究,发现在毛细胞的顶部有机械门控离子通道[7~9],也称机械电换能通道,该通道对机械力的作用非常敏感。当静纤毛处于相对静止状态时,有少部分通道开放并伴有少量的内向离子流,如果用玻璃微杆使静纤毛向动纤毛一侧弯曲时,通道进一步开放,大量阳离子内流引起去极化而产生感受器电位。当静纤毛向背离动纤毛的一侧弯曲时通道关闭,内向离子流停止并出现外向离子流,造成膜的超极化。
尽管有些详细机理尚不清楚,但有一点是肯定的,即毛细胞顶部纤毛的机械偏转是诱发毛细胞兴奋的适宜刺激,而纤毛偏转的方向则是诱发毛细胞是否兴奋的必要条件。此外,声音强弱能影响基底膜振动的幅度,声音强则振幅大,声音弱则振幅小。而音调的不同,则主要靠各个频率引起的行波最大振幅处的毛细胞的不同来辨别,这就是耳蜗的感音换能作用。不难看出,听觉经验的丰富程度紧紧依赖于内耳的感受器,即听毛细胞的功能。在听觉生成过程中,毛细胞发挥着近乎于不可思议的作用:它们不但可以测量原子的尺寸,还可以将刺激范围从静态的输入转换成以10 kHz为单位的频率。如果不考虑它所回应的刺激类型,毛细胞就是一个生物张力标尺。可见,听毛细胞将机械能转换成电信号的过程是极其复杂而神秘的。
1.3.2毛细胞纤毛束的分子生物学特征 耳蜗毛细胞顶部表面的静纤毛以阶梯形排成3列,蜗底处静纤毛短,靠近蜗顶静纤毛逐渐变长,认为这种梯度变化很可能是产生音频排列和调谐功能的形态学基础。尽管耳蜗的功能活动机制还有许多不明之处,但可以肯定,内、外毛细胞的结构与功能的相互偶连,是柯蒂器的机械-电转换的关键,而毛细胞顶部纤毛束的机械偏转是毛细胞兴奋的前提条件。
毛细胞起源于外胚层表面,并且保留了上皮的特征。单个毛细胞是柱状或者烧瓶状的,没有树突和轴突(图4)[3]。毛细胞顶点的周围同非感觉性的支持细胞相连,一种特殊的盐溶液(内淋巴液)浸润着细胞的顶部,在细胞的基底侧面内淋巴液同普通的细胞外液(外淋巴液)完全分离。在紧密连接的下方,由中间连接或带状桥粒为毛细胞提供一种有力的机械吸附作用。
作为声音(机械)刺激的感受器的毛细胞的纤毛束,从平坦的细胞顶部表面突出(图4a、b)。依它所出现的动物种类的不同,一束纤毛束长度可以从小于1 μm到超过100 μm不等。这种细胞器是一丛20~300个的柱状突起,这些立方纤毛呈六边形排列。由于这些立体纤毛在整个细胞表面上的长度不一,纤毛束看上去就像皮下注射器针头的尖端部,呈一个倾斜的结构(图4b)。每个立体纤毛都是一个刚性的圆柱,它的细胞骨架包含有一束纤维蛋白横向连接起来的肌动蛋白丝,横向连接使得立体纤毛比一束毫无连接的肌动蛋白丝更加坚固。立体纤毛的核心由一层管状的浆膜壳覆盖着。尽管一根独立的立体纤毛的直径在沿着其长度的方向似乎是不变的,但是仍然可以用千分尺测出它在逐渐变细。当立体纤毛变细的程度从0.5 μm到大约其直径的四分之一时,其肌动蛋白从1 000个减少到只有几十个。这些减少了的微丝构成了支架使立体纤毛固定于细胞表皮之上。在位于顶部膜表面的下方,由相互连接的肌动蛋白构成一个稍厚的网状结构,而立体纤毛的骨架是其突出的底部最细的地方,所以,作用在立体纤毛尖端的机械力会使它绕着它的基部的插入处旋转。

图4脊椎动物单个毛细胞的形态学特征及其纤毛束的扫描电镜图
a. 蛙的内耳感觉上皮示意图,可见毛细胞具有明显的上皮细胞特征。圆柱形的毛细胞在其顶端周围通过一种连接复合体与相邻的支持细胞相连接。机械敏感性细胞器从细胞的顶部表面一直延伸到纤毛束.在细胞膜的基底外侧的表面形成传入和传出突触联系
b. 毛细胞的顶部表面的扫描电镜图,显示纤毛束突出到内淋巴约8 μm。 每个纤毛束由大约60根静纤毛组成,每个圆柱形的静纤毛都有一个逐渐变细的根部,按不同长度呈阶梯状排列。在纤毛束较高的一边树立着仅仅一根动纤毛,动纤毛具有类似轴突样的结构,并且在其顶尖部呈球状膨隆。毛细胞被支持细胞所包绕,支持细胞的顶部表面分布着一些较短的微绒毛。
