桑黄类大型真菌中多酚类物质研究进展
2024-04-08阮馨颖宋婷婷张作法金群力蔡为明
阮馨颖,宋婷婷,张作法,金群力,陈 春,蔡为明,*
(1.中国计量大学 生命科学学院,浙江 杭州 310019; 2.浙江省农业科学院 园艺研究所,浙江 杭州 310021)
桑黄在中国已有2 000多年的药用历史,最早以“桑耳”为名出现在秦汉时期的《神农本草经》中,直至隋唐时期甄权的《药性论》首次出现了“桑黄”一词,其中记载“桑耳,使。一名桑臣,又名桑黄。味甘、辛,无毒。能治女子崩中带下,月闭血凝,产后血凝,男子痃癖,兼疗伏血,下赤血。”[1-2]随着现代生物学的深入研究,桑黄类真菌成为一类形态高度相似并且具有药用价值的大型木腐型多孔真菌的泛称,因其含有多酚类、多糖类、萜类、甾体类等多种丰富的生物活性成分[3],成为开发利用的热点。
桑黄类真菌的多酚物质是一类具有多元酚结构的重要次级代谢产物,按结构可分为吡喃酮类、呋喃酮类、黄酮类、酚酸类、儿茶素类化合物。酚羟基是多酚物质中的重要官能团,是其发挥多种活性作用的关键结构[4]。研究表明,桑黄多酚类物质具有抗肿瘤[5]、抗氧化[6]、保肝[7]、抑菌[8]、抗病毒[9]、神经保护[10]、抗炎、降血糖[11]等药理活性,在医药领域有着广泛的应用潜力。此外,不同品种、不同产地、野生与人工栽培的桑黄类真菌在多酚物质的含量与活性上存在明显差异[12-14],其合成量随生长时期表现出一定的规律性与相关性,因此桑黄类真菌多酚物质的代谢调控也具有重要意义。本文结合已有的研究报道,重点就桑黄类真菌的分类,以及桑黄类真菌中多酚物质及其药理活性、提取方法与代谢调控等方面作简要综述,并对桑黄类真菌多酚物质的后续研究方向进行展望,以期推动桑黄产业的持续发展。
1 桑黄类真菌的种属分类现状
古代文献中记载了桑黄的药用价值,学者们借助现代分类学技术,以及对古代本草著作的考证,以期寻找真正的“桑黄”[2,15]。学者通过分析其形态特征和核糖体内转录间隔区核酸序列(ITS),发现真正的“桑黄”是未曾发表过的新种,并将其命名为Inonotussanghuang[16-17]。桑黄主要分布于中国、日本、韩国,野外仅生长在桑属(Morus)的树干。包海鹰等[2]依据历代本草文献中所记载“桑黄”的生境、形态特征和药用功效,结合现代药理活性等研究,认为粗毛纤孔菌I.hispidus是古代中药记载的“桑黄”。本文讨论的桑黄类真菌是属于真菌界(Fungi)担子菌门(Basidiomycota)锈革孔菌目(Hymenochaetales)锈革孔菌科(Hymenochaetacea)[18]的一类具有相似形态特征的药用真菌,是“桑黄”真菌的一个广泛的概念。2016年以前,桑黄类真菌主要被归为3个属,分别是木层孔菌属(Phellinus)、纤孔菌属(Inonotus)、嗜蓝孢孔菌属(Fomitiporia)[19]。至2016年,Zhou等[20]和侯伟男等[21]广泛收集分析锈革菌科(Hymenochaetaceae)的样本,对此前定义的“桑黄”及相近种重新分类,并建立了新属——桑黄孔菌属(Sanghuangporus)。之后桑黄孔菌属不断得到补充,目前该属包含15个种,见表1。所有中药材和保健食品的基原种类必须辨别清楚,但桑黄类真菌长期以来存在种属分类混乱的问题[22],此前一直以农副产品的身份进入市场,市场宣传混乱[23],使得桑黄的药用资源无法充分利用,对桑黄相关产业造成了严重影响。在不断探究梳理桑黄类真菌的种属分类问题和亲缘关系的基础上,研究人员进行桑黄类真菌相关研究时,首先要明确桑黄类真菌的类别,再对其亲缘关系进行甄别,并正确使用拉丁名称,可避免出现同名异种、同种异名的问题,以便后续资源的整合利用。近几年,湖北、安徽、吉林、甘肃、浙江等省份相继出台了桑黄中药材质量或中药饮片炮制规范的地方标准,助力桑黄产业发展[24]。
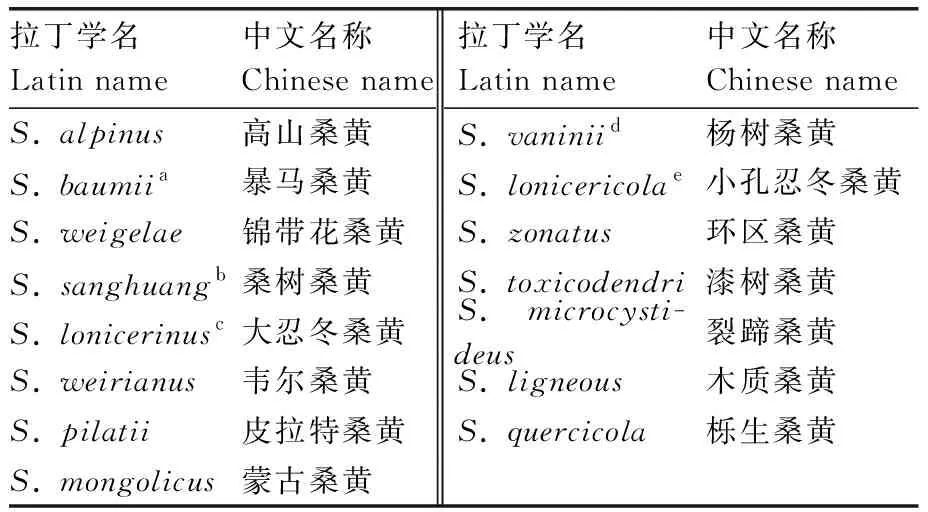
表1 桑黄孔菌属类桑黄成员Table 1 Members of Sanghuangporus
2 桑黄类真菌多酚物质及其药理作用
2.1 桑黄多酚物质
桑黄类真菌的多酚物质在大型药用真菌中表现突出[25],也是桑黄类真菌次生代谢产物的重要组成部分,种类繁多,结构不尽相同,具有代表性的重要桑黄类真菌酚类物质见表2。陈万超等[3]、昝立峰等[26]对近几年在桑黄类真菌中分离得到的多酚物质进行了归纳总结,根据结构特征可将其分为吡喃酮类、呋喃酮类、黄酮类、酚酸类、儿茶素类化合物。桑黄类真菌的多酚物质以吡喃酮类化合物为主,主要有苯乙烯基吡喃酮和苯并吡喃酮及其衍生物,是一种黄色多酚类色素[27],有牛奶树碱(hispidin)、桑黄酚A(hispolon)、hypholomine B、骨碎补内酯(davallialactone)、纤孔素菌(inoscavin A)、桑黄素D(phelligridin D)、baumin等。研究者使用超高效液相色谱串联三重四级杆飞行时间质谱法(UPLC-QTOF-MS)在野生与栽培杨树桑黄(S.vaninii)中鉴定出40个化合物,吡喃酮酚类物质占20个[13]。同样利用该技术鉴定出S.sanghuang与S.vaninii有19种相同的多酚物质,其中17个为吡喃酮酚类化合物[14]。宋吉玲等[28]采用液相色谱质谱联用法(LC-MS)分析鉴定了P.igniarius醇提物中15种化学成分,大多数为苯乙烯基吡喃酮衍生物。phellinsin A、phellinusfurans A、phellinusfurans B、烟碱酮(inotilone)是在桑黄类真菌中分离得到的呋喃酮类化合物,具有免疫调节、抗真菌等多种作用[29-30]。桑黄类真菌中的黄酮类物质研究较多,其体外抗氧化能力与总黄酮含量之间具有良好的量效关系[31],分离得到较多的是二氢黄酮类衍生物,如二氢莰非素、圣草素、山萘酚、柚皮素等。酚酸是一类含有酚环的有机酸,在桑黄类真菌中发现了没食子酸、原儿茶酸、咖啡酸、原儿茶醛等几种单苯环酸类化合物。儿茶素类化合物是茶多酚的主要成分(占70%~80%)[32],但在桑黄中存在较少,鉴定到的有儿茶素、表没食子儿茶素没食子酸酯、没食子儿茶素没食子酸酯、儿茶素没食子酸酯等[13]。
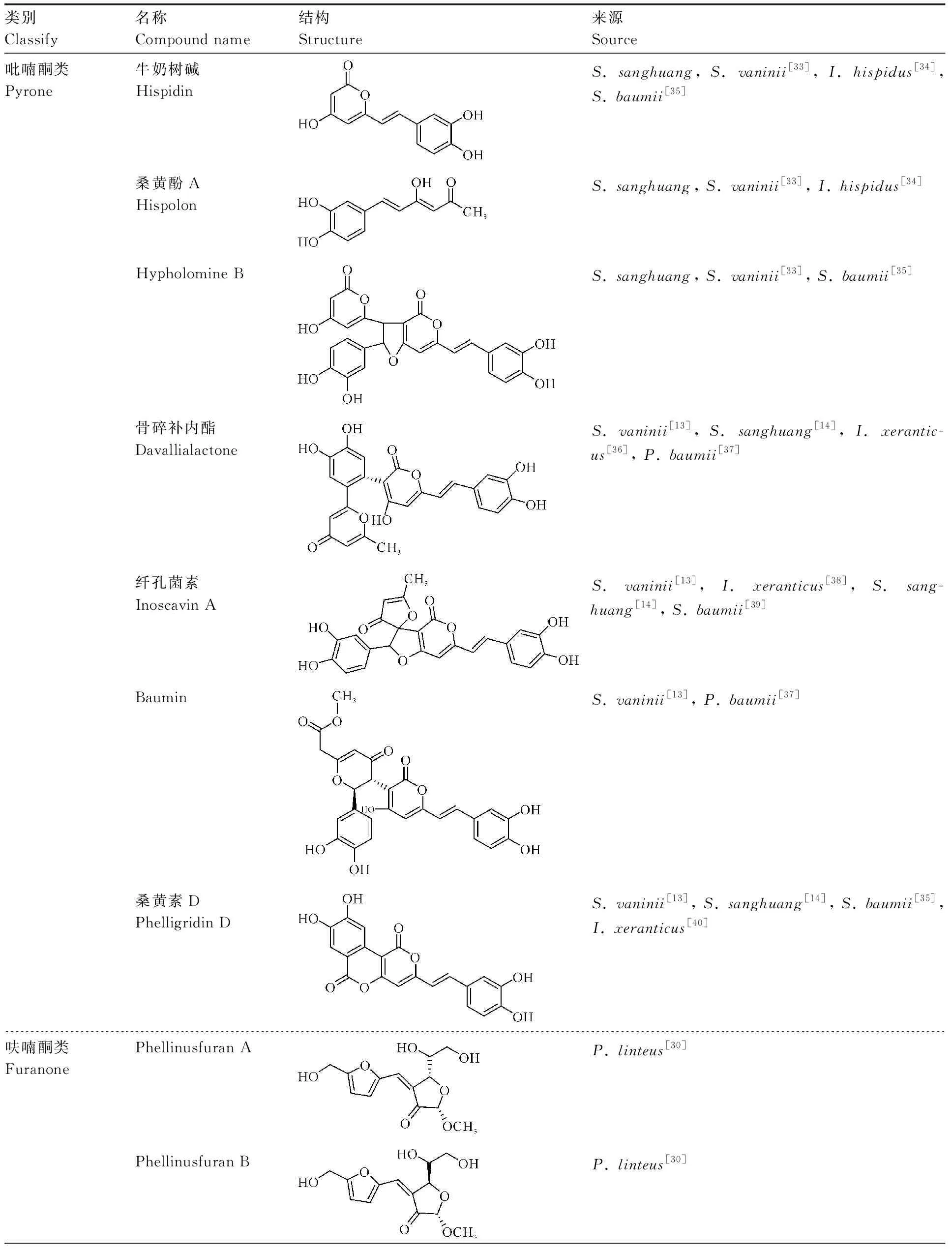
表2 桑黄类真菌中代表性多酚类物质Table 2 Representative polyphenols in sanghuang fungi
2.2 抗肿瘤
1968年,日本学者发现桑黄水提物对小鼠肉瘤细胞S-180增殖抑制率高达98.6%[46]。自此,桑黄的抗肿瘤作用研究不断深入。hispolon是桑黄中的一种活性酚类化合物,其可通过靶向PI3K/Akt、MAPK和NF-κB等多种细胞信号通路,诱导细胞凋亡,暂停细胞周期,抑制细胞转移,从而发挥抗癌作用,且hispolon中的苯乙烯吡咯碳骨架具有药物开发前景[5]。hispolon可以抑制Hep3B细胞转移,并以时间和剂量依赖的方式抑制Hep3B细胞的生长,并诱导其细胞S期阻滞和凋亡[47-48]。许多抗癌药物通过诱导细胞凋亡和细胞周期阻滞来实现抗癌功能,推测hispolon具备开发为新型化疗药物的潜力。杨树东等[44]从粗毛纤孔菌的醇提物中分离到一种多酚类物质,即3,3′-亚甲基双{6-[2-(3,4-二羟苯基)乙烯基]-4-羟基-2H-吡喃-2-酮},噻唑蓝(MTT)抗肿瘤筛选结果表明,该化合物对人肝癌细胞HepG2增殖起抑制作用,半抑制浓度IC50值为2.3 μg·mL-1,并初步断定该化合物可诱导HepG2细胞凋亡。昝立峰等[49]测定发现,鲍马桑黄的乙酸乙酯萃取物对人非小细胞肺癌细胞(NCI-H1299)、胃癌细胞(SGC-7901)、肝癌细胞(HepG2)、乳腺癌细胞(MDAMB-231)、宫颈癌细胞(Hela)均表现出不同程度的细胞毒性,推测其主要活性成分为吡喃酮类多酚。
2.3 抗氧化
自由基对机体产生的氧化损伤与多种疾病的形成相关,机体需要自我产生或从外界摄取的方式获得超氧化物歧化酶、过氧化氢酶等内源性抗氧化剂或中草药、维生素、微量元素等外源性抗氧化剂,通过这些抗氧化剂形成屏障,应对自由基造成的细胞损害[50]。吕国英等[14]测定了野生桑树桑黄和杨树桑黄子实体乙醇提取物的DPPH清除率、超氧阴离子清除率和β-胡萝卜素漂白实验,结果表明,桑树桑黄和杨树桑黄均具有很强的抗氧化活性,并利用关联分析明确了乙醇提取物中的主要成分属于多酚类物质。昝立峰等[51]对粗毛纤孔菌子实体中的乙醇提取物进行了抗氧化活性测定,当提取物浓度为800 mg·L-1时,抗氧化活性达到89.52%,与市场上常用的合成抗氧化剂BHA的能力相当(抗氧化活性达到92.10%),乙醇提取物中多酚物质的含量高达69.76 μg·mg-1,有望开发成为一种新的天然抗氧化剂。Jung等[52]从褐黄纤孔菌(Inonotusxeranticus)和裂蹄木层孔菌(P.linteus)的液体发酵液中分离到4种多酚物质,分别为hipidin及其二聚体、3,14-二乙酰吡啶、hypholomine B和1,1-二苯乙烯基吡啶,它们均表现出较强的抗氧化能力。
2.4 抗肝纤维化
研究表明,减弱或消除肝纤维化的致病因素可在一定程度上逆转肝纤维化。因此,抗肝纤维化药物的研究对干预治疗肝纤维化和降低重症肝病的发病率有重要的预防意义[53]。从裂蹄木层孔菌(P.linteus)中提取的多酚物质phellinulin A,对二甲亚硝胺和TGF-1激活的大鼠肝星状细胞有显著影响,可阻止其转化为肌成纤维细胞样细胞,证明phellinulin A对体内肝纤维化有抑制作用[7]。脂肪肝与肝纤维化常相继发或合并存在,是引起肝纤维化的重要病因之一[54]。P.linteus的乙酸乙酯提取物(PL-EA)可通过调节高脂肪和高果糖饮食喂养小鼠的脂质和葡萄糖代谢稳态来缓解非酒精性脂肪肝(NAFLD);从PL-EA中纯化得到的主要活性成分hipidin和hypholomine B,可以显著降低油酸和棕榈酸诱导的脂质积累水平,具有缓解NAFLD的作用[55]。
2.5 抗炎
Kim等[56]研究表明,从暴马桑黄(P.baumii)中分离出来的酚类物质phelligridin D可以减少脂多糖诱导的人牙周韧带细胞中炎性分子的产生。Wu等[57]研究表明,来自P.linteus的hispolon通过抑制脂多糖或脂磷壁酸诱导的细胞中的iNOS/NO产生和炎性细胞凋亡,有效抑制了巨噬细胞中的炎症反应。冯佳亮[58]从粗毛纤孔菌中分离得到4种多酚类物质,其主要成分phelligridin D可显著降低脂多糖诱导的巨噬细胞RWA264.7产生的NO、IL-1β和IL-6的含量,其抗炎分子机制是通过抑制NF-κB信号通路上相关蛋白p65的磷酸化,进而抑制NF-κB信号传导。
2.6 抗病毒
Awadh Ali等[9]从粗毛纤孔菌子实体中分离出2种酚类化合物hispolon和hispidin,两者表现出明显的抗甲型流感病毒A(H1N1和 H3N2)和乙型流感病毒的活性。Hwang等[35]从P.baumii子实体乙醇提取物中分离出的多酚物质hispidin、hypholomine B、inoscavin A, davallialactone、phelligridin D表现出良好的抗病毒活性,均可抑制H1N1、H5N1和H3N2神经氨酸酶活性,并减少病毒诱导的细胞病变效应的量。
2.7 其他
桑黄类真菌中多酚物质除具有上述抗肿瘤、抗氧化、抗肝纤维化、抗炎、抗病毒活性以外,还有预防卒中、降血糖、神经保护、抑菌等作用。
Suabjakyong等[59]发现,含有多酚的P.igniarius提取物对小鼠神经母细胞瘤细胞系中的丙烯醛毒性有保护作用,通过P.igniarius多酚提取物(20 μg·kg-1)对脑卒中小鼠进行腹膜内处理,可有效减少梗死,表明P.igniarius多酚提取物可用于预防缺血性卒中。用适当浓度的桑黄多酚hispdin(2 μmol·L-1)预处理帕金森病(PD)细胞模型[1-甲基-4-苯基吡啶(MPP)诱导MES23.5细胞]获得的线粒体功能障碍减弱,结果表明,hispdin对MPP诱导的线粒体功能障碍和细胞凋亡具有神经保护作用,说明hispdin可被视为预防PD的辅助药物[10]。Yang等[11]对2型糖尿病小鼠给予不同剂量的P.baumii酚类物质(50、100、150 mg·kg-1),持续灌胃60 d,模型小鼠血糖明显降低,以150 mg·kg-1剂量给药时效果与降糖药二甲双胍相当。桑黄子实体与菌丝体中的不同溶剂提取物对大肠埃希菌、金黄色葡萄球菌等多种致病菌都表现出一定的抑菌效果,其抑菌活性可能与含有的黄酮类化合物有关[8,60]。
3 桑黄类真菌多酚物质的提取方法
将多酚类化合物从桑黄类真菌中分离提取出来是研究其分子结构与生物活性的关键一步,其提取方法主要有溶剂萃取法、物理场强化提取法、超临界萃取、大孔吸附等。
3.1 溶剂浸提法
溶剂浸提法主要分为水溶剂提取和有机溶剂提取2种,通常遵循相似相溶原则,根据目标化合物选择合适的提取溶剂是该方法的关键[61]。常用的溶剂主要有水、甲醇、乙醇、正丁醇、乙酸乙酯、丙酮等,各溶剂的不同特性导致所提取的多酚在含量和生物活性上存在较大差异。陈志娜等[62]利用4种不同溶剂(70%乙醇、甲醇、乙酸乙酯、氯仿)从粗毛纤孔菌中提取总黄酮和总多酚,并测定其提取物的含量、抗氧化活性和抑菌活性,结果表明,甲醇提取的总黄酮和总多酚含量最高,70%乙醇提取物的抗氧化能力最强,甲醇提取物和70%乙醇提取物抑菌活性最优。张洋洋等[63]采用氯化胆碱与尿素组成的新型深共熔溶剂提取杨树桑黄多酚,该方法最优条件的提取率(23.17±0.88 mg·g-1)显著优于传统溶剂提取法(12.45 mg·g-1)。溶剂浸提法因工艺简单、稳定性好、成本较低的优点而被广泛应用,但存在提取效率低、溶剂残留、试剂消耗大、污染环境等缺点。
3.2 物理场强化提取法
在传统的溶剂浸提中加入某种物理场,如微波、超声波等,相比于溶剂浸提法可提高萃取效率,减少活性物质的降解[64]。冯子旺等[65]采用超声提取法,在乙醇体积分数60%,超声时间30 min,温度50 ℃,料液比1∶25的条件下,桑黄总多酚提取率可达26.4 mg·g-1。陈晓平等[66]研究表明,微波时间、微波功率的恰当选择可显著提高桑黄黄酮的提取率。
3.3 超临界萃取法
CO2超临界萃取法是一种绿色、高效的分离技术,具有无毒、无残留溶剂、避免热敏性物质和易氧化物质变性的优点,对于提取天然化合物具有明显优势[67]。张倩等[68]使用CO2超临界萃取法萃取的桑黄黄酮类化合物含量达4.67%,得率显著高于其他提取方法。
3.4 大孔树脂吸附法
刘奥建立了I.baumii发酵与桑黄素LA异位树脂分离耦合的新工艺,将发酵罐与树脂层析柱连接,当发酵进行到一定程度时,启动分离模式,发酵液经树脂层析柱吸附后,返回发酵罐中继续发酵,大孔树脂洗脱得到目标化合物桑黄素LA[69]。桑黄素LA属于苯乙烯吡喃酮类多酚化合物,在发酵过程中随桑黄素LA浓度的增大而产生负反馈作用,抑制了桑黄素LA的增长,该工艺采用发酵与分离结合的方法,利于及时从发酵液中分离抑制性产物或有毒物质,既有效提升了I.baumii发酵中桑黄素LA的产量与分离效果,也是抑制性产物生产分离的新思路,为发酵与分离工艺一体化奠定了基础。
4 桑黄类真菌多酚物质的代谢调控
初级代谢过程在所有生物体中基本一致,植物和微生物还利用初级代谢产物进行次级代谢活动[70-71],进一步合成或分解得到各种不同的次级代谢物,且具有多样性。不同于初级代谢,次级代谢通常受到营养物质、生长速度、反馈控制、诱导、酶和转录因子等因素的影响[71],所以研究者可通过控制次生代谢产物合成,从而调整目标代谢物的合成量。
多酚类物质作为一类极其重要的天然次级代谢活性物质,广泛分布于植物和大型真菌中,不仅赋予其抵御外界各种生物与非生物胁迫的能力,也对人类医药的发展具有重要意义,图1为桑黄类真菌中的苯丙烷代谢途径。因此,加强桑黄多酚物质代谢的研究,对于调控该类真菌中关键药用成分的合成与提高桑黄品质具有重要意义。
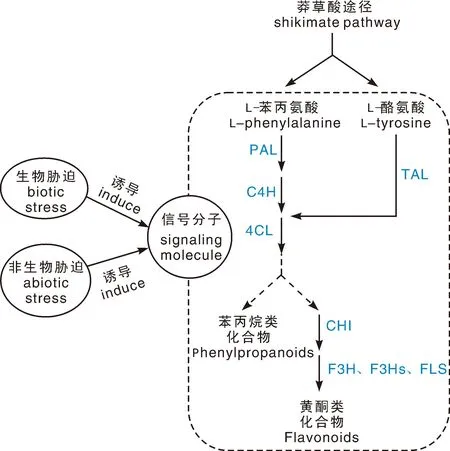
PAL,苯丙氨酸解氨酶;C4H,肉桂酸-4-羟化酶;4CL,4-香豆酰-CoA连接酶;TAL,酪氨酸解氨酶;CHI,查耳酮异构酶;F3H,黄烷酮3-羟化酶;FLS,黄酮醇合成酶。PAL, Phenylalaninammo-nialyase; C4H, Cinnamate-4-hydroxylase; 4CL, 4-Coumarate:CoA ligase; TAL, Tyrosine ammonilyase; CHI, Chalcone isomerase; F3H, Flavanone-3-hydroxylase; FLS, Flavonol synthase.图1 桑黄类真菌中的苯丙烷代谢途径Fig.1 Metabolic pathways of phenylpropane in sanghuang fungi
桑黄多酚类物质合成量的变化随生长时期表现出一定的规律性与相关性,不同菌种间的多酚含量差异明显,且表现出的抗氧化活性不同[72-75]。此外,培养条件、外源添加物、诱变等外界因素也与酚类物质的合成密切相关,例如,适宜的碳氮比与发酵条件能加快菌丝的生长速度和多酚等次生代谢物的累积[76-77]。乙酸镁、硝普钠等外源物的加入可有效促进桑黄菌株合成多酚类化合物[78-79]。选用0.1%秋水仙素诱变桑黄原生质体,筛选出的菌株的多酚含量显著高于对照菌株[80]。也有研究利用紫外辐射、低能离子射线、常压室温等离子体、脉冲光等物理诱变手段刺激桑黄多酚物质的积累或筛选高产多酚的桑黄菌株[81-83]。
植物中的多酚类物质主要通过莽草酸途径和丙二酸途径合成[84],而在大型真菌中主要由莽草酸途径提供前体物质合成多酚类物质[85]。由莽草酸途径合成的2种芳香族氨基酸,分别为L-苯丙氨酸和L-酪氨酸,它们作为苯丙烷类代谢途径的起始物,几乎是合成一系列苯丙烷化合物的前体物质[86-87];在桑黄培养时加入苯丙氨酸、香豆素等前体添加物,可大幅提高桑黄黄酮类化合物的含量,也保持了其生物活性[88]。综合蛋白质组学和代谢组学的差异分析,发现苯丙烷类化合物的上调是多酚含量水平升高的重要原因[89]。在桑黄类真菌中是否也存在与植物相同或类似的苯丙烷类代谢途径,这需要进一步探究。全基因组与转录组信息的完善为该类真菌中次级代谢产物合成相关基因的研究提供了理论依据[90-92]。Shao等[93]通过筛选S.sanghuang基因组,寻找到7个与类黄酮合成有关的核心功能基因与假定基因,但未发现编码查尔酮合成酶(CHS)的基因,该酶是植物中针对类黄酮合成的首个酶,这表明桑黄类真菌中存在苯丙烷途径,但与植物存在差异,缺失的基因可能被亲缘关系较远的超基因家族所代替。Hispidin作为桑黄类真菌中特有的多酚类物质,被认为来自苯丙烷途径[29,89]。酚类物质的合成受苯丙烷类代谢途径中相关酶活性和相关基因表达的影响。结合S.baumii中有关类黄酮合成的靶向代谢组学和转录组学分析发现,SbPAL、Sb4CL、SbIFR基因与苯丙烷途径中类黄酮合成相关,其中SbPAL基因是起到调控作用的关键基因;通过遗传转化技术在S.baumii中过表达SbPAL基因,SbPAL的转录水平和苯丙氨酸解氨酶(PAL)活性显著增加,同时促进了S.baumii中类黄酮的积累[94]。此外,还在I.baumii和S.vaninii中获得了多酚合成相关的苯丙烷类代谢途径的2个关键基因IbPAL[95]、SvCHI[96],这有利于全面了解桑黄多酚合成过程中相关酶与基因的功能。
一氧化氮(NO)作为一种重要信号分子参与了桑黄类真菌中次级代谢物的代谢调节,添加外源NO供体硝普钠可以显著调节S.vaninii菌丝中多酚和三萜类化合物的积累,并刺激多酚物质phelligridin D的增长,改变酚类物质的组成成分[79]。在I.baumii(S.baumii)发酵液中添加AgNO3,多酚类物质相比对照增加了1.57倍[97]。桦褐孔菌(I.obliquus)与桑黄类真菌同属于锈革孔菌科,也是一类极具药用价值的大型真菌,NO在该类药用菌中的多酚调控机制研究较为深入。Zheng等[98]在没有脂氧化物质的培养基中添加多酚诱发剂和NO供体,与添加抑制剂进行比较,证明在I.obliquus中NO通过独立于脂氧化物质刺激多酚产生的信号通路,介导诱发剂促进多酚合成增加。NO的瞬时积累导致PAL、4-香豆酰-CoA连接酶(4CL)的亚硝基化,硫氧还蛋白(Trx)和硫氧还蛋白还原酶(TrxR)在亚硝基谷胺酰胺还原酶(GSNOR)受抑制的情况下脱硝基化,使PAL、4CL的催化活性增加,使得多酚积累增加。GSNOR和TrxR的活性不仅决定了PAL、4CL等参与多酚生物合成酶的活性,还改变了酚类物质组成成分,但总体来说,抑制GSNOR提高了TrxR活性,使总酚含量增加[99-101]。I.obliquus与P.morii共培养增强了编码苯丙烷途径PAL和4CL基因的表达,最终促进了苯乙烯基吡喃酮多酚的积累,原因是真菌间的相互胁迫会激发内源NO的产生,而NO可能会增强防御相关基因的表达[101-102]。这意味着NO在多酚类天然产物的合成中起关键作用,特别是在桑黄类真菌中,其潜在的调控作用机制仍有待揭示。
5 展望
目前,桑黄类真菌的研究主要集中在物种分类、化学成分和药理活性上。光谱、质谱、色谱、波谱等检测技术与提取技术的发展使得该类化合物可被精准鉴定和快速分离,是促进桑黄类真菌中多酚物质深入研究的基础。与其他大型药用菌相比,桑黄类真菌中的多酚类化合物含量相对较高,并展现了出色的药理作用,应该充分利用其天然产物在预防与治疗各种复杂性疾病的独特优势,加强桑黄多酚药用成分的筛选,并进一步明确其药理作用与活性机制。桑黄类真菌的次级代谢产物在很大程度上尚未开发,要不断加深对次级代谢衍生物的了解,利用好宝贵资源。由于桑黄类真菌的多酚合成途径与植物存在差异,且有大量基因的功能尚未注释,深入研究关键合成酶活性和缺失基因,有助于明晰桑黄类真菌多酚物质的生物合成途径。如何应用现代生物手段高产天然产物或在不同阶段稳定获取目标化合物成为人们关注的焦点,因此需要注重桑黄类真菌中多酚类物质的合成途径与代谢调控机制的研究。此外,在中药材中,质量的优劣直接影响其药用功效,优良品种的选育是提高中药材质量的有效手段[103],而传统栽培模式的优化,只能从宏观角度调节桑黄类真菌的生长状态,难以控制重要活性物质的含量,可通过分子手段结合遗传育种,推进优良品种选育,提高桑黄产业价值与药用价值,进一步提高大众对桑黄类真菌和菌物药的认知度和接受度,促进桑黄产业的发展。
