花椰菜“坐球高度”性状的主基因+多基因遗传分析
2024-04-08蔡诗怡虞慧芳王建升沈钰森顾宏辉盛小光
蔡诗怡,虞慧芳,王建升,祝 彪,沈钰森,顾宏辉,盛小光,*
(1.浙江农林大学 园艺科学学院,浙江 杭州 311300; 2.浙江省农业科学院 蔬菜研究所,浙江 杭州 310021)
花椰菜(Brassica.oleraceavar.botrytis,2n=2x=18)是芸薹属甘蓝种的重要蔬菜作物,以花序分生组织异常增殖形成的特异花球为产品器官,其柔嫩多汁、味道鲜美、粗纤维少、营养丰富,深受广大消费者的喜爱[1]。我国是世界上花椰菜种植面积最大的国家,种植区域几乎覆盖全国各地,播种面积已达约50万hm2,种植产值超过200亿元,对促进农业增效和农民增收起到重要的作用[2]。
花椰菜产业是一个劳动密集型产业,从播种、移栽、农事管理到花球采收,都需要消耗大量的人力和物力。随着农业机械化研发力度的加强,蔬菜种植过程的机械化水平在逐步提高。目前,花椰菜的播种、移栽及部分农事操作正在逐步实现机械化,但是成熟花球采收仍然几乎全部依靠人工,不仅人力、物力投入大,且生产效率极低。花椰菜成熟花球机械化采收难以实现的关键限制因素之一是其“坐球高度”(第一片真叶到花球底部的主茎长度)短。目前市场主栽的早中熟品种的坐球高度约为7~13 cm,由于花球底部至地面的距离太短,导致机械化采收异常困难[3]。因此,依据花球采收设备的实际需求,采用分子设计育种,定向、快速选育出适宜“坐球高度”的花椰菜品种,以提升花椰菜花球的机械化采收水平,对推动花椰菜产业进一步发展具有重大意义。
对于没有主球的其他作物来说,“坐球高度”在一定程度上与株高表型相似。在小麦[4]﹑水稻[5]﹑玉米[6]、高粱[7]等大田作物中,对株高性状的遗传规律,控制基因遗传定位、克隆及其功能机制分析等方面开展了大量研究,并且挖掘到了一批经典的株高控制基因:“Dn”“USn”和“Rhtn”[8]。在十字花科作物中,相关研究主要集中在油菜作物上,刘超[9]和赵波[10]分别克隆了矮秆基因ds-1和ds-3,其中ds-3参与编码赤霉素(GA)信号转导抑制因子 DELLA 蛋白,阐明了油菜株高形成的部分调控机制。对这些基因进行合理的控制与应用对实现油菜作物的稳产和丰产具有重要意义。花椰菜作物的新品种选育工作推进得较快,浙江省乃至全国的科研机构联合种业公司,育成了一大批优秀的花椰菜品种,但其相关的分子生物学研究起步较晚,研究的广度和深度还远远不够[11]。近年来,花椰菜全基因组测序和染色体级别组装工作的完成,加速了花椰菜重要农艺性状控制基因的挖掘和利用等研究[12],但是“坐球高度”性状的遗传规律分析及QTLs定位研究,未见相关报道。
本研究以主茎较高的芥蓝和主茎短的花椰菜为亲本构建的六世代群体(P1、P2、F1、F2、B1、B2)为研究材料,以主茎长度(第一片真叶到花球/蕾底部的主茎长度)和叶痕间距(主茎中间部分第一和第三个叶痕间的距离)两个指标来锚定“坐球高度”性状,在植株现球/蕾期对这两个指标进行测定和统计分析。利用主基因+多基因遗传模型的分析方法检测花椰菜“坐球高度”性状的遗传规律[13],为进一步遗传定位和挖掘克隆到控制此性状的主效基因提供研究基础。
1 材料与方法
1.1 实验材料
利用早熟(生育期约52 d)、紧实型花椰菜ZAASC4101作为母本(F7代自交系)与芥蓝ZAASJ1401为父本(F6代自交系,生育期约45 d)构建六世代群体:P1(ZAASJ1401)、P2(ZAASC4101)、F1、F2、B1(F1×ZAASJ1401)、B2(F1×ZAASC4101),试验材料由浙江省农业科学院蔬菜研究所花椰菜和青花菜育种研究室提供。
1.2 实验方法
试验在本单位海宁杨渡科研基地开展。六世代群体 P1、P2、F1、F2、B1、B2测量株数分别为 50、50、50、219、70、185,一共624株。2021年7月10日将六个世代的材料分别播种进行穴盘育苗,2021年8月7日双行定植于露地大田中。株距约 50 cm,行距约 60 cm。进行常规的田间管理。
在植株现球/蕾期,使用卷尺测量两个指标,第一是主茎高度:花球底部到第一片真叶的主茎长度,第二是叶痕间距:主茎中部第一个和第三个叶痕间的距离(图1)。

图1 主茎高度和叶痕间距性状的测量标准Fig.1 Measurement criterias for the traits of main stem height and leaf scar spacing
1.3 遗传分析
利用数量性状遗传分析 R 软件包 SEA v2.0进行“坐球高度”性状的遗传多世代联合分析以及主茎高度和叶痕间距的频率分布直方图分析[14];采用IECM算法和极大似然法对主茎高度和叶痕间距的遗传参数进行估算,根据极大对数似然值(maximum likelihood value, MLV)和最小信息量准则(Akaike’s information criterion, AIC)选出最适的几个备选模型,然后从中筛选出最佳模型;最后利用最小二乘法计算出该模型的主基因和多基因效应值和遗传参数等数据[15]。
2 结果与分析
2.1 表型数据分析
母本花椰菜(ZAASC4101,P2)和父本芥蓝(ZAASJ1401,P1)主茎平均高度分别为8.12 cm和28.68 cm,两者相差20.56 cm。F1主茎平均高度为20.98 cm,处于双亲之间并且大于双亲主茎高度的平均值(图2)。P1、P2和F1这3个群体内的植株基因型一致,群体内植株主茎高度数值的分布相对集中,标准差(standard deviation, SD)数值较低并且均小于1.0 cm;P2群体变异系数(coefficient of variation,CV)稍大,为10.15%,P1和F1群体分别为2.94%和3.79%。F2、B1和B2群体植株的主茎平均高度分别为22.95 cm、27.89 cm和20.21 cm。由于这3个群体均为遗传分离群体,植株主茎高度数值分布的离散度相对较高,其中B1群体的SD和CV值最小,分别为2.18 cm和7.83%,F2群体最大,分别为5.48 cm 和23.88%。总体来说,B1群体总体偏向于主茎高的父本ZAASJ1401。B2和F2群体的变异系数远高于亲本,而B1群体的变异系数介于两个亲本之间,说明B1群体的变异性较低,但是B2和F2群体具有较高的遗传变异(表1)。

表1 花椰菜六世代群体的主茎高度和叶痕间距性状的遗传分析Table 1 Genetic analysis of main stem height and leaf scar spacing traits in the six-generation populations of cauliflower

a,ZAASJ1401父本;b,ZAASC4101母本;c,F1植株;d~k,F2植株。a, ZAASJ1401 male parent; b, ZAASC4101 female parent; c, F1 plant, d-k: F2 plants.图2 六世代群体中的双亲、F1和F2植株Fig.2 Parents, F1 and F2 plants in the six-generation populations
对于叶痕间距性状,本研究测定了P1、P2和F2群体植株的数值。双亲的叶痕间距平均值分别为5.47 cm和2.56 cm,两者相差2.91 cm。F2群体植株的叶痕平均间距为4.00 cm,SD和CV值分别是0.98 cm和24.50%。总体来说,这三个群体植株叶痕间距数值的分布趋势和上述主茎高度一致。F2群体植株这两个性状的相关性分析也显示,主茎高度与叶痕间距呈极显著正相关,相关系数为0.652,表明主茎高度越高,叶痕间距越长。
F2群体植株的主茎高度和叶痕间距的表型变异最为丰富,变异系数分别为23.88%和24.50%。从F2群体性状的表型频率分布图中可以看出(图3),主茎高度和叶痕间距性状均呈连续性近似正态分布的特征,说明它们是典型的数量性状,在受到主效基因控制的同时也受到其他微效位点的影响。

图3 F2群体主茎高度及叶痕间距的频率分布(柱形)、拟混合分布(红线)与成分分布(黑线)Fig.3 Frequency (column), mixed (red line), and component (black line) distributions for main stem height and leaf scar spacing traits in F2 population
2.2 遗传模型
对六世代群体植株的主茎高度性状进行主基因+多基因混合遗传分析,共获得5类24种遗传模型,从中选择AIC值最小的3个模型作为备选模型,包括两对加性-显性主基因+加性-显性-上位性多基因遗传模型(MX1-AD-ADI, AIC=3 045.244)、两对加性-显性-上位性主基因+加性-显性-上位性多基因遗传模型(MX2-ADI-ADI, AIC=3 048.592)和两对连锁主基因+加性-显性-上位多基因遗传模型(PG-ADI, AIC=3 051.222)。由于这3个模型的AIC值比较接近,所以均作为备选模型(表2)。
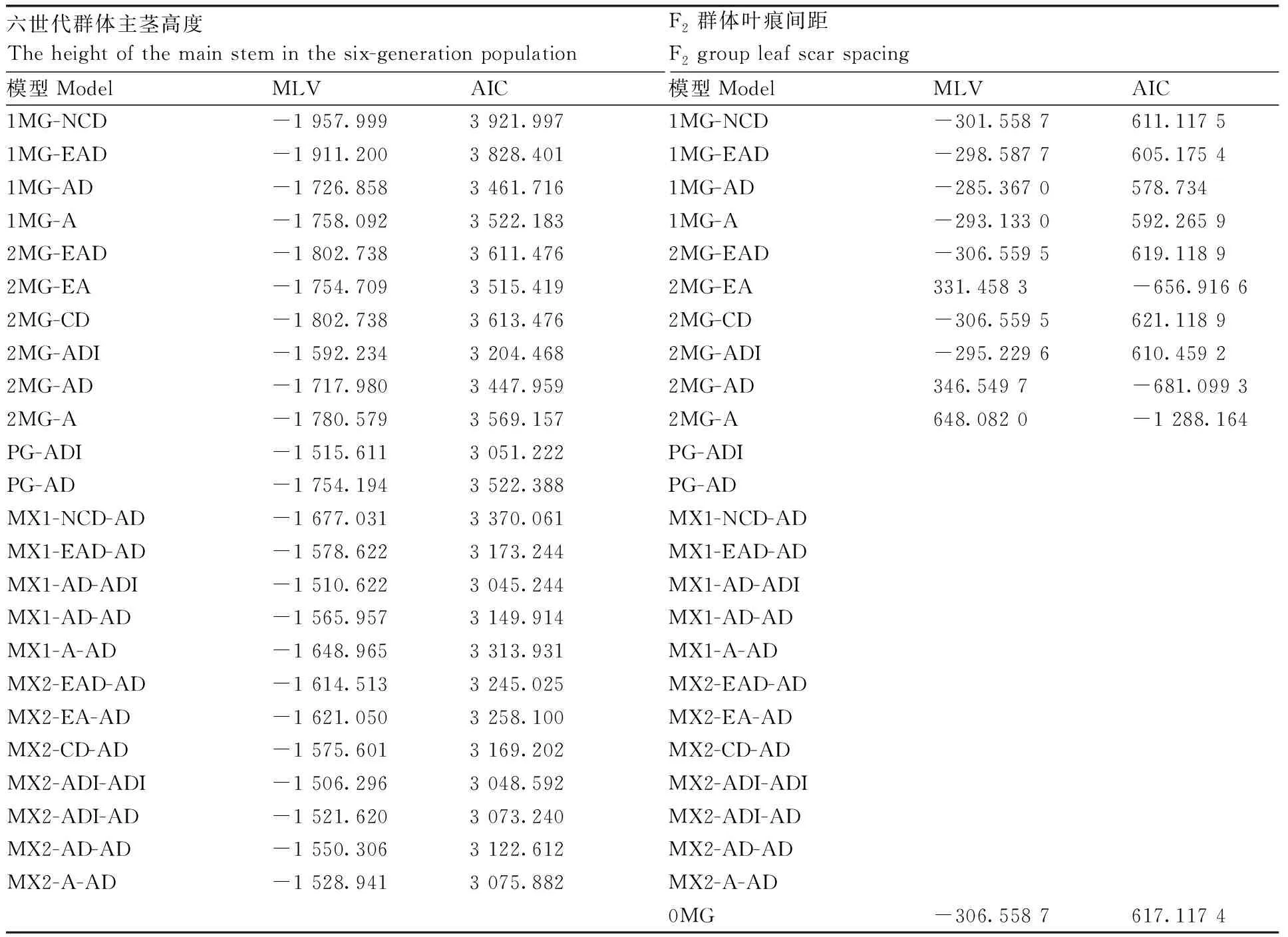
表2 六世代群体主茎高度和F2群体叶痕间距的MLV和AIC值Table 2 The estimation of MLV and AIC value of the main stem height of the six-generation populations and the leaf scar spacing of the F2 population of cauliflower
对F2群体植株的叶痕间距性状进行遗传分析,共获得3类11种遗传模型。其中两对主基因下的加性模型(2MG-A)的AIC值最小,为-1 288.164,并且与其他模型的AIC值相比差异显著,因此2MG-A被认为是最适模型。
综合主茎高度的六世代遗传分析和叶痕间距F2群体的遗传分析结果,花椰菜“坐球高度”性状的最适遗传模型是:MX2-ADI-ADI。
2.3 遗传参数的估计
根据已确定的最适宜遗传模型,计算出参数的极大似然估计值,应用最小二乘法进行一阶和二阶的遗传参数估计(表3)。

表3 F2群体最适模型遗传参数估计值Table 3 Estimated value of optimal model genetic parameters for F2 population
花椰菜“坐球高度”性状的遗传模型符合MX2-ADI-ADI。一阶参数的结果表明,该性状受两对主基因控制,其加性效应值da=1.727 6,显MG,主基因模型;MX,主基因+多基因混合模型;PG,多基因模型;A,加性效应;D,显性效应;I,上位效应;E,相等。
性效应ha=-0.399 7,为负向显性,并且da的绝对值大于ha的绝对值,说明控制花椰菜主茎高度的主基因以加性效应为主。二阶参数的估算结果表明,F2世代的主基因遗传率(h2mg)为 13.49%,多基因遗传率(h2pg)为84.35%。说明花椰菜“坐球高度”性状在F2群体中以多基因遗传为主,并同时受主基因+多基因的共同影响。
3 讨论
花椰菜“坐球高度”是决定其花球能否机械化采收的重要标准之一,明确其遗传规律是进一步挖掘控制该性状的主效基因的基础,对利用连锁分子标记进行辅助选择、快速选育出适宜机械化采收的花椰菜新品种具有重要意义。由于花椰菜的分子生物学研究相对滞后,已经报道的遗传定位的相关研究主要集中在产量、球色、荚叶等性状[16-17],针对“坐球高度”性状的遗传规律分析和遗传定位研究,至今无相关报道。本研究利用芥蓝和花椰菜杂交构建的六世代群体为研究材料,采用主茎高度和叶痕间距两个性状指标共同来锚定花椰菜“坐球高度”性状,并在群体植株现球/蕾期进行这两个指标的测定,可以明显地区分构建群体中单株“坐球高度”的差异。
本研究中F2世代分离群体的主茎高度和叶痕间距的频次分布表现为连续分布,具有明显的数量性状遗传特征。因此,可以使用盖钧镒[15]的植物数量性状遗传体系分析方法对主基因和多基因的遗传效应进行研究,明晰花椰菜“坐球高度”性状的遗传规律。该方法在多种作物的株高类性状遗传规律研究中都有成功运用,何文昭等[18]对玉米株高和穗位高性状进行了不同环境下的数量遗传分析,发现在三个环境下符合的遗传模型各不相同,这与材料的遗传背景、测试时间以及环境状况等因素的不同有关;解松峰等[19]研究表明,小麦株高性状符合两对累积作用的主基因+加性作用多基因混合遗传模型;李英双等[20]认为,甜荞株高符合两对加性-显性-上位性主基因+加性-显性多基因模型;而李军庆等[21]的研究结果表明,油菜半矮秆株高性状符合一对加性-显性主基因+加性-显性-上位性多基因模型。本研究中,结合主茎高度的六世代和叶痕间距的F2群体遗传规律分析,结果表明,花椰菜“坐球高度”性状的最适遗传模型是两对加性-显性-上位性主基因+加性-显性-上位性多基因遗传模型(MX2-ADI-ADI)。但是,植物数量性状分离分析方法推测的只是概念上的基因,而基因数目的多少和各基因对“坐球高度”性状的贡献率还需要利用后续的QTLs遗传定位结果从分子水平上进行深入解析。
遗传率是决定世代选择速度的关键因子。本研究中“坐球高度”性状的主基因+多基因遗传率达到97.84%,说明其后代主要受到遗传因素的影响,可以开发连锁分子标记在早期世代进行定向辅助选择。但是,本研究结果同时也显示,该性状的多基因遗传率大于主基因遗传率。因此,在后续的育种工作中,在考虑主基因影响的同时需要同时关注多基因微效位点带来的影响。另外,环境也是影响花椰菜“坐球高度”性状的重要因素之一,在栽培过程中需要注意。
4 结论
本研究综合了主茎高度的六世代遗传分析和叶痕间距F2群体的遗传分析结果,表明花椰菜“坐球高度”为典型的数量遗传性状,由两对加性-显性-上位性主基因+加性-显性-上位性多基因遗传模型控制。本研究结果为花椰菜“坐球高度”性状的主效基因挖掘及连锁分子标记开发打下良好的基础。
