外源激素对干旱胁迫下黑麦草生理特性的影响及抗旱性评价
2024-03-28刘亚西杨梦含刘金平苗增建
刘亚西,毛 芮,杨梦含,刘金平,苗增建,王 丹
(1.西华师范大学生命科学学院 西南野生动植物资源保护省部共建教育部重点实验室, 四川 南充 637009;2.珙县林业和竹业局,四川 珙县 644500;3.简阳市市政园林管理中心, 四川 简阳 641400;4.西宁奥来利农业开发有限公司, 青海 西宁 812100)
植物生长过程受多种环境胁迫因子的影响,干旱胁迫导致植物细胞膜的结构与功能损伤,生理代谢紊乱,影响植物正常生长发育,直至死亡,对农牧业生产造成巨大经济损失,因此,如何应用农业措施缓解干旱胁迫伤害,成为农牧业生产中所关注的重要问题。目前国内外学者针对提高植物抗旱性开展了多方面研究,如选育抗旱性品种[1]、干旱锻炼[2]、施加外源激素[3-4]等。其中施加外源激素是增强植物抗旱性的重要途径之一,在多种植物如玉米(Zea mays)[5]、小麦(Triticum aestivum)[6]、高羊茅(Festuca elata)[7]等开展了相关研究。因此,探索通过施加外源激素提高植物的耐旱性,对干旱地区农牧业的生产具有重要的意义。
植物外源激素可通过影响植物的生理特性、光合特性以及内源激素的合成来改变植物的生长过程[8]。研究表明,在逆境胁迫下施加适当浓度的外源植物激素可以缓解胁迫对植物的伤害作用。赤霉素(gibberellin,GA3)作为重要的植物生长调节激素能促进植物种子萌发与幼苗生长[9],使用200 和300 mg·L–1GA3浸种处理能够明显提高15% PEG 胁迫下多年生黑麦草种子的发芽率[10]。脱落酸(abscisic acid,ABA)因其能够积极响应干旱、高温等逆境胁迫而被称为植物的“应激激素”[11]。郭贵华等[12]的研究表明,外源ABA 在孕穗期能够有效缓解干旱胁迫对水稻(Oryza sativa)生理代谢功能的损伤。此外,有研究表明,外源施加60 μmol·L-1ABA 可通过降低猕猴桃(Actinidia deliciosa)幼苗的细胞膜透性,增加抗氧化酶活性来提高其抗旱性[13]。褪黑素(melatonin,MT)作为一种新型植物激素在提高植物在非生物胁迫下的抗逆性具有积极的作用[14]。在干旱胁迫下,外源褪黑素能通过增加黑麦草抗氧化酶活性和脯氨酸的积累来增强黑麦草的抗旱性[15]。油菜素内酯(2,4-epibrassinolide,EBR)作为一种在植物抗逆性方面发挥重要作用的新型植物激素[16],在提高植物抗旱性方面也有很多研究。王雨婷等[17]的研究表明,外源0.1 mg·L-1EBR 可有效提高葡萄(Vitis vinifera)幼苗的抗旱能力。综上,外源施用GA3、ABA、MT 和EBR 4 种激素都具有缓解植物干旱胁迫的作用,针对不同种类的植物具有深入研究的意义和实践应用前景。
多年生黑麦草(Lolium perenne)为禾本科黑麦草属草本植物,是世界上广泛栽培的冷季型草坪草和畜牧业养殖用草,因其生长速度快,根系分布较浅,在建植过程中需要灌溉用水较多,容易因缺水受到干旱胁迫的影响[18]。通过外源激素来提高多年生黑麦草的抗旱性研究具有广泛的应用价值。为此,在自然干旱胁迫下施加外源激素GA3、ABA、MT 和EBR 通过隶属函数分析法研究不同外源植物激素对干旱胁迫下多年生黑麦草的缓解效应,以期为外源植物激素在黑麦草生产中的应用提供理论依据。
1 材料与方法
1.1 试验材料
供试多年生黑麦草品种为‘雅晴’(Yatsyn),种子购自百绿国际草业有限公司(种子于2019 年生产,发芽率为90%,千粒重为1.752 5 g)。外源植物激素MT (纯度≥ 99.0%)和GA3(纯度≥ 90.0%)购自北京索莱宝科技有限公司,ABA 购自北京兰博利德商贸有限公司,EBR 购自上海源叶生物技术有限公司。
1.2 试验设计
采用盆栽试验,于2022 年4 月1 日将黑麦草种子均匀播种在育苗盆(高11 cm,上口径14.8 cm)中,待幼苗生长60 d 后,进行干旱胁迫与叶面喷施植物激素处理。共设6 个处理,对照组(CK1)、干旱组(CK2)、50 μmol·L-1MT + 干旱(T1)、0.1 μmol·L-1EBR + 干旱(T2)、10 μmol·L-1ABA + 干旱(T3)、200 mg·L–1GA3+ 干旱(T4),每个处理3 个重复,每个重复4盆,共72 盆。CK1正常浇水(每隔2 d 浇一次水,每次以浇透为准),不喷激素;CK2不浇水不喷激素;T1、T2、T3、T4喷施激素后不浇水。每天下午18:00 对植株正面叶面进行喷施,程度为叶面湿透无滴水,连续喷施3 d 后,在激素喷施结束的第2 天(定义为干旱胁迫0 d)开始对黑麦草进行干旱断水处理。自然干旱处理9 d,分别在处理的0、3、6、9 d取样。于同一时间段取植株相同位置叶片(从基部向上数第3 片叶)用液氮速冻后保存在-80 ℃冰箱中用于之后生理指标的测定。取样和测定生理指标时各处理均进行3 次重复。
1.3 测定指标与方法
1.3.1 叶片相对含水量
采用饱和称重法[19]测定叶片相对含水量,分别称取鲜重(Wf),将叶片在清水中浸泡24 h 之后吸干叶片表面水分称取饱和重(Wt),之后在烘箱中烘干至恒重称取干重(Wd)。叶片相对含水量 (leaf relative water content, LWC) = (Wf-Wd)/(Wt-Wd) × 100%。
1.3.2 叶片相对电导率
叶片相对电导率的测定参照方紫雯[20]的方法,稍有改进。在干净的试管中加入 10 mL 去离子水,测定去离子水空白电导率(E0),取待测的新鲜叶片(0.1 g),剪成相同长短的碎段放在 10 mL 去离子水中,遮光放置 12 h,测量浸泡液的电导率(E1),沸水浴30 min,冷却后,进行电导率测定(E2)。叶片相对电导率 = (E1–E0)/(E2–E0) × 100%。
1.3.3 生理生化指标
丙二醛(malondialdehyde,MDA)、脯氨酸(proline,Pro)、可溶性糖(soluble sugar,SS)、可溶性蛋白( soluble protein,SP)、超氧化物歧化酶(superoxide dismutase,SOD)、过氧化物酶(peroxidase,POD) 、过氧化氢酶(catalase,CAT)均采用苏州格锐思生物科技有限公司试剂盒进行测定。
1.3.4 内源激素
4 种内源激素玉米素核苷(zeatin riboside,ZR)、GA3、ABA、 生长素(auxin,IAA)含量采用酶联免疫法,利用北京北农天一生物技术有限公司试剂盒进行测定。
1.4 数据统计分析
采用Excel 2010 对数据进行处理和制图,应用SPSS 22.0 软件对数据进行单因素方差分析,用Duncan多重比较进行差异显著性分析(P< 0.05)。用模糊数学中的隶属函数法[21-22]对不同外源激素处理下黑麦草抗旱能力进行综合评价。隶属函数法可将每个指标值转换为 (0,1)的纯数,优化不同指标之间的可比性,对不同处理下的指标进行综合分析,最后根据各处理平均隶属度值由大到小进行排序,位次越靠前,表明处理的缓解效果越好。隶属函数法可弥补单一指标带来的偏差,使评定的结果与实际结果较为接近,是近年来抗旱性评价方法中常用的方法。
隶属函数法的计算公式为R(Xi) = (Xi-Xmin) /(Xmax-Xmin) 。式中:Xi为指标i的测定值,Xmin和Xmax分别为所有处理下某一指标的最小值和最大值;R(Xi) 为指标Xi的隶属函数值。如果所测指标与抗旱性呈负相关关系,则用反隶属函数计算其抗旱隶属度R(Xi) = 1 - (Xi-Xmin) /(Xmax-Xmin)。将各个处理下的测定指标的隶属函数值相加求平均值后进行综合评价各处理下的抗旱性强弱。
2 结果与分析
2.1 不同外源激素对干旱胁迫下黑麦草幼苗叶片相对含水量的影响
随着干旱天数的增加,黑麦草叶片相对含水量逐渐减少(图1)。与CK1相比,CK2处理下叶片相对含水量在第6 天和第9 天分别显著下降了69.47%和88.60% (P< 0.05)。外源施加植物激素可以增加叶片相对含水量,缓解干旱胁迫对黑麦草的伤害。在第6 天和第9 天,T1、T2、 T3、T4处理下与CK2相比叶片相对含水量表现为增加的趋势,其中T3和T4处理效果显著(P< 0.05)。在第6 天,T3、T4处理下相对含水量分别是CK2的1.78和2.53 倍,在第9 天分别是CK2的3.06 和4.17 倍。
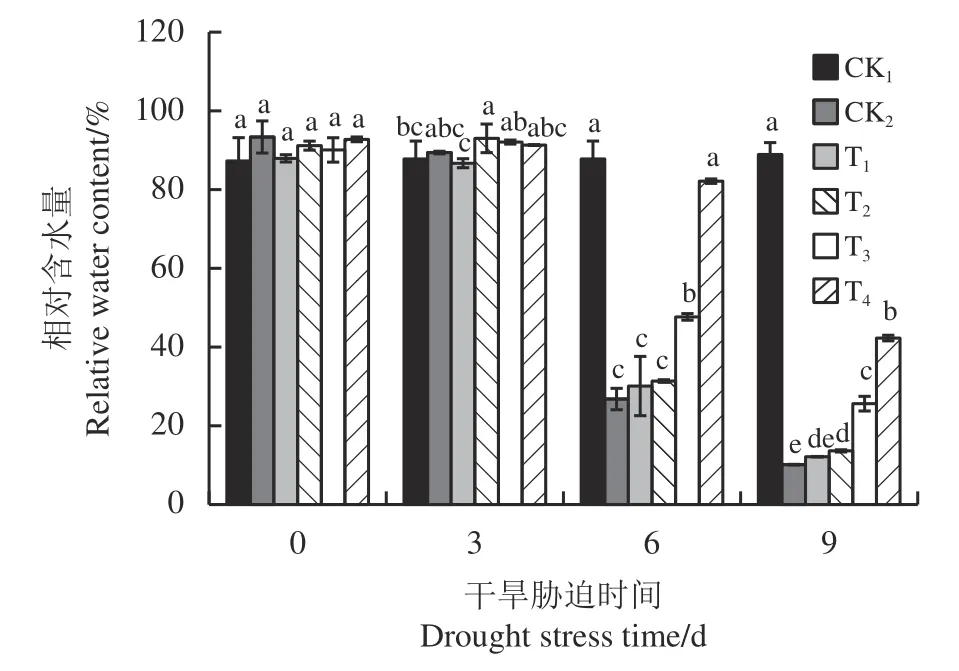
图1 不同外源激素对干旱胁迫下黑麦草幼苗叶片相对含水量的影响Figure 1 Effects of different exogenous hormones on relative water content in ryegrass seedling leaves under drought stress
2.2 不同外源激素对干旱胁迫下黑麦草幼苗叶片相对电导率的影响
在干旱胁迫的第0 天和第3 天,各处理间黑麦草相对电导率无显著差异(P> 0.05),T1处理第3 天除外(图2)。在第6 天和第9 天,CK2处理下相对电导率为CK1的13.53 和16.12 倍;4 种外源植物激素处理下相对电导率相比对照表现为下降的趋势,其中T4处理下相对电导率最低。在第6 天,与CK2相比,T1、T2、 T3和T4处理下相对电导率分别显著下降了10.60%、11.07%、70.34% 和85.69% (P< 0.05)。在第9 天, T1、T2处理下相对电导率与CK2相比没有显著差异(P> 0.05)。T3、 T4处理下分别显著下降了5.55%和73.51% (P< 0.05)。

图2 不同外源激素对干旱胁迫下黑麦草幼苗叶片相对电导率的影响Figure 2 Effects of different exogenous hormones on the relative electrical conductivity of ryegrass seedlings under drought stress
2.3 不同外源激素对干旱胁迫下黑麦草幼苗丙二醛含量的影响
随着干旱胁迫天数的增加,相比CK2,外源激素处理下MDA 含量表现出逐渐增加的趋势(图3)。与CK1相比,CK2处理下MDA 含量在第3、6 、9 天显著增加(P< 0.05),分别为CK1的3.13、2.93、2.43倍。外源激素处理可降低MDA 含量。在第3 天,各激素处理下MDA 含量虽有一定程度的降低,但与CK2相比没有显著差异。在第6 天,T4处理下MDA 含量最低,与CK2相比显著降低了23.09% (P<0.05)。在第9 天,T3和T4处理下MDA 含量与CK2相比显著降低了26.90%、39.89% (P< 0.05)。与CK2相比T1和T2处理虽有一定程度的降低,但无显著差异(P> 0.05)。

图3 不同外源激素对干旱胁迫下黑麦草幼苗丙二醛含量的影响Figure 3 Effects of different exogenous hormones on malondialdehyde content in ryegrass seedlings under drought stress
2.4 不同外源激素对干旱胁迫下黑麦草幼苗渗透调节物质的影响
Pro 作为渗透调节物质,在干旱胁迫下含量增加(表1)。在干旱胁迫第6 天和第9 天,与CK2相比,T1、 T2、T3、 T4处理下Pro 含量显著降低(P<0.05)。在第6 天,与CK2相比T4处理下Pro 含量显著降低了68.02% (P< 0.05)。T1、T2、T3处理下与CK2相比显著降低了7.74%、7.64%、7.28% (P< 0.05),但3 个处理间没有显著差异(P> 0.05)。在干旱胁迫第9 天,T3处理下Pro 含量最低,与CK2相比显著降低了41.97% (P< 0.05)。

表1 不同外源激素对干旱胁迫下黑麦草幼苗脯氨酸含量的影响Table 1 Effects of different exogenous hormones on proline content in ryegrass seedlings under drought stress mg·g-1
随着干旱天数的增加,各处理下SS 含量整体表现为先降低后升高的趋势(表2)。在第6 天和第9 天 ,CK2处理下黑麦草SS 含量最高,为CK1的5.62、2.57 倍。干旱胁迫下,外源施加植物激素可降低SS 含量。在第6 天,T3和T4处理与CK2相比显著降低了66.43% 和68.45% (P< 0.05)。在第9 天,T2、T3、T4处理下与CK2相比显著降低了26.20%、30.33%、63.77% (P< 0.05)。T1与CK2处理间均无显著差异(P> 0.05)。
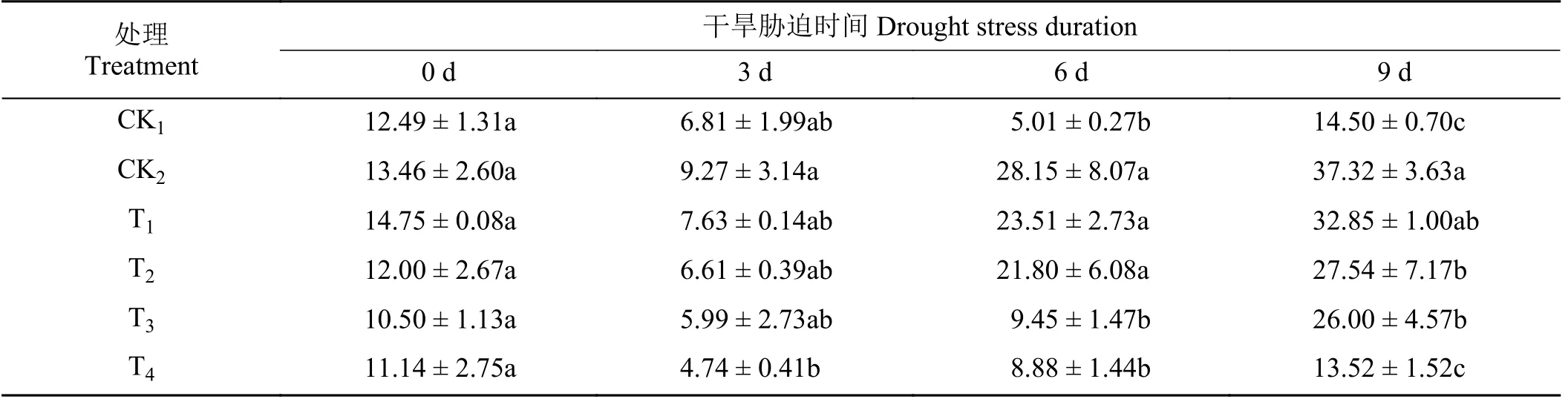
表2 不同外源激素对干旱胁迫下黑麦草幼苗可溶性糖含量的影响Table 2 Effects of different exogenous hormones on soluble sugar content in ryegrass seedlings under drought stress mg·g-1
随着干旱胁迫时间的增加,在第0、3、6 天CK2处理下SP 含量与CK1相比无显著差异(P> 0.05)(表3)。在第6 天,T3和T4处理下SP 含量与CK2相比显著降低了44.34% 和49.25% (P< 0.05),T1和T2处理与CK2相比虽有一定程度降低但无显著差异(P> 0.05)。在第9 天,与CK1相比,CK2处理下SP 含量显著降低了56.99% (P< 0.05)。各外源植物激素处理下SP 含量均明显增加,T1、T2、T3、T4处理下SP 含量分别为CK2的1.36、2.56、2.67、2.15 倍。
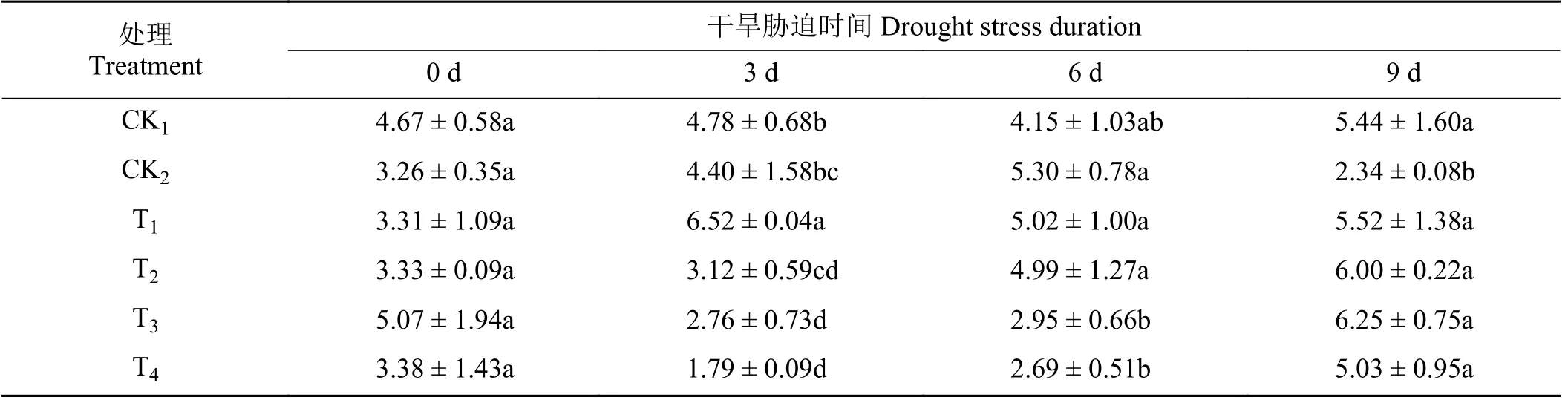
表3 不同外源激素对干旱胁迫下黑麦草幼苗可溶性蛋白含量的影响Table 3 Effects of different exogenous hormones on soluble protein content of ryegrass seedlings under drought stress mg·g-1
2.5 不同外源激素对干旱胁迫下黑麦草幼苗抗氧化酶活性的影响
随着干旱胁迫天数的增加,CK2处理下黑麦草叶片的SOD 酶活性逐渐增加(表4),在干旱胁迫第6 天和第9 天,SOD 酶活性为CK1处理的2.84、4.03倍。4 种外源植物激素处理下,除T4处理SOD 酶活性表现为先增加后减少的趋势,其余处理表现为先减少后增加的趋势。在第6 天,与CK2相比T1、T2、T3、T4处理下SOD 酶活性显著降低了39.55%、39.87%、34.52% 和22.84%(P< 0.05)。在第9 天,与CK2相比T1、T2、T3、T4处理下 SOD 酶活性显著降低(P< 0.05),其中,T4处理最低,与CK2相比降低了47.17%,T1和T2处理之间无显著差异(P> 0.05)。
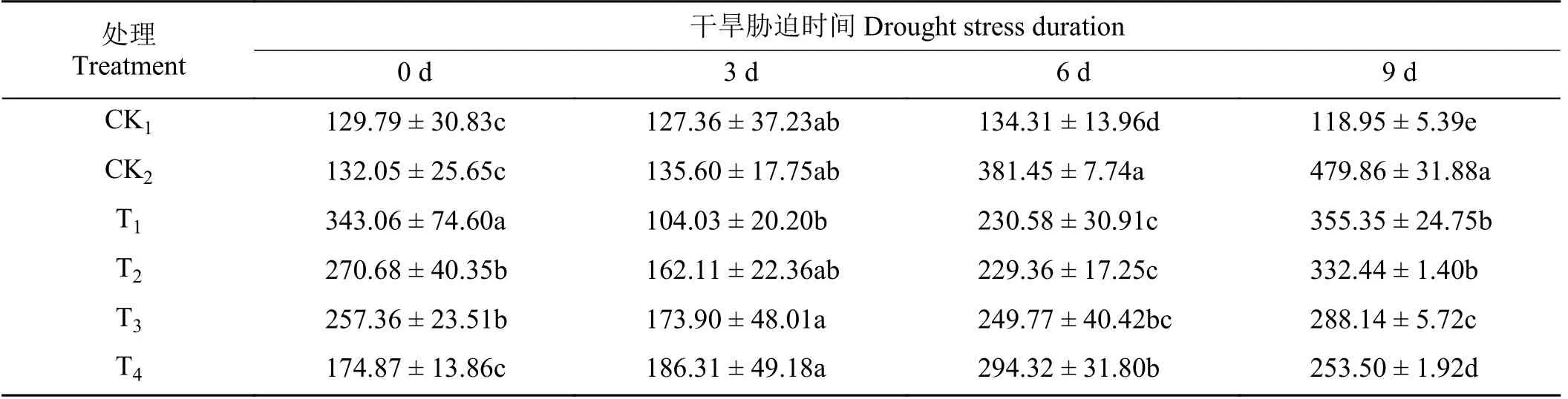
表4 不同外源激素对干旱胁迫下黑麦草幼苗超氧化物歧化酶酶活性的影响Table 4 Effects of different exogenous hormones on superoxide dismutase activity of ryegrass seedlings under drought stress U·g-1
随着干旱天数的增加,CK2处理下黑麦草叶片POD 酶活性逐渐增加(表5)。在第3、6、9 天与CK1相比显著增加(P< 0.05)。在第6 天,与CK2相比T1、T2、T4处理下,POD 酶活性显著降低了18.62%、42.42%、18.31% (P< 0.05),T3处理与CK2无显著差异(P> 0.05)。在第9 天,CK2处理下,POD 酶活性为CK1处理的3.72 倍;4 种外源植物激素处理下,T2、T3、T4处理下SOD 酶活性较CK2显著降低(P<0.05),其中T4处理下POD 酶活性最低,较CK2降低了62.71%。
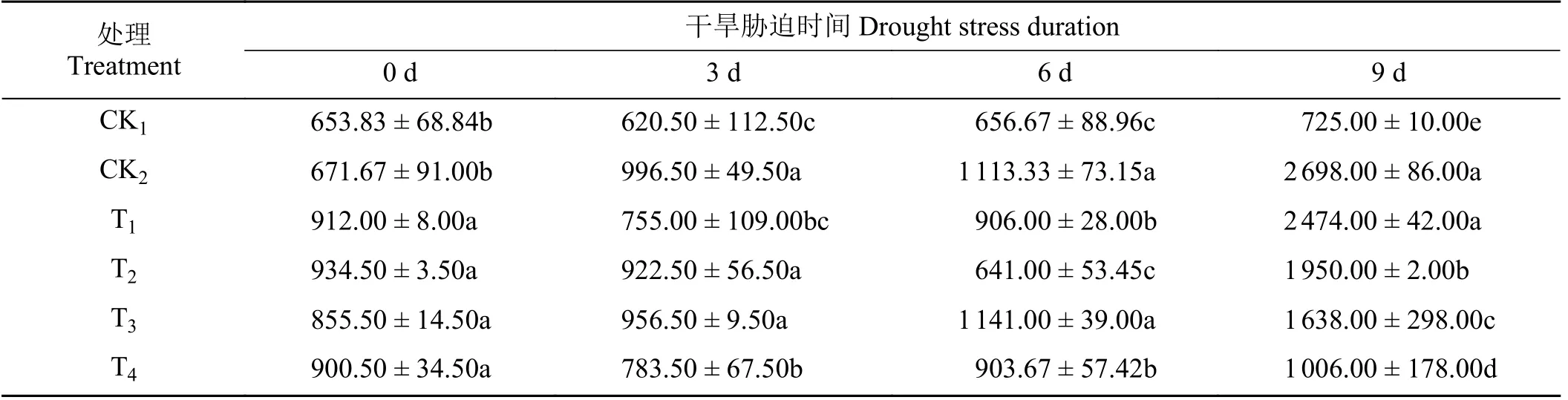
表5 不同外源激素对干旱胁迫下黑麦草幼苗过氧化物酶活性的影响Table 5 Effects of different exogenous hormones on peroxidase enzyme activity in ryegrass seedlings under drought stress U·(g·min)-1
CK2处理下黑麦草叶片CAT 酶活性随干旱胁迫天数的增加表现为先增加后减少的趋势(表6),在第6 天达到最大。在第6 天,与CK2相比T2、T3、T4处理下CAT 酶活性显著降低(P< 0.05)。其中T4处理下最低,较CK2降低了36.71%。在第9 天,与CK2相比4 种外源植物激素处理下CAT 酶活性有一定程度的增加,但各处理间无显著差异(P>0.05)。
2.6 外源激素对干旱胁迫下黑麦草叶片内源激素含量的影响
在干旱胁迫第6 天,黑麦草内源激素ZR、GA3、ABA、IAA 含量CK2较CK1显著增加(P< 0.05) (表7)。外源施加植物激素后,黑麦草内源ZR 和ABA 含量减少,GA3含量增加,IAA 含量除T2处理增加外,T2、T3、 T4处理均减少。ZR 含量在CK2处理下比CK1处理显著增加了15.32% (P< 0.05),T3处理下ZR 含量最低,与CK2相比,显著降低了32.08% (P<0.05)。与CK1相比,CK2处理下GA3含量显著增加了23.31% (P< 0.05)。T4处理下GA3含量最高,分别为CK2、T1、 T2、T3处理下的7.87、4.84、5.85、7.04 倍。干旱胁迫下,黑麦草内源激素ABA 含量增加,CK2处理为CK1的18.65 倍。T1、 T2、T3、 T4处理下黑麦草内源ABA 含量与CK2相比均显著降低(P< 0.05),其中T4处理下内源ABA 含量最低,较CK2降低了81.67%。 黑麦草内源IAA 含量在CK2处理下为CK1的2.79 倍。除T2外,其余处理下内源IAA 含量降低。T3处理下IAA含量最低,较CK2显著降低了57.08% (P< 0.05)。

表7 外源激素对干旱胁迫第6 天黑麦草叶片内源激素含量的影响Table 7 Effects of exogenous hormones on the endogenous hormone contents of ryegrass leaves on the sixth day of drought stress ng·g-1
2.7 不同外源激素处理下黑麦草的抗旱性综合评价
用隶属函数法对各处理下的多年生黑麦草的抗旱性进行了综合评价,取干旱胁迫第6 天和第9 天各指标隶属函数值的平均值,隶属函数值越大,表明抗旱性越强。如表8 所列,CK1、CK2、T1、T2、T3、 T4处理下的隶属函数值分别为0.90、0.02、0.23、0.34、0.49、0.67,抗旱性大小排序为CK1> T4>T3> T2> T1> CK2。
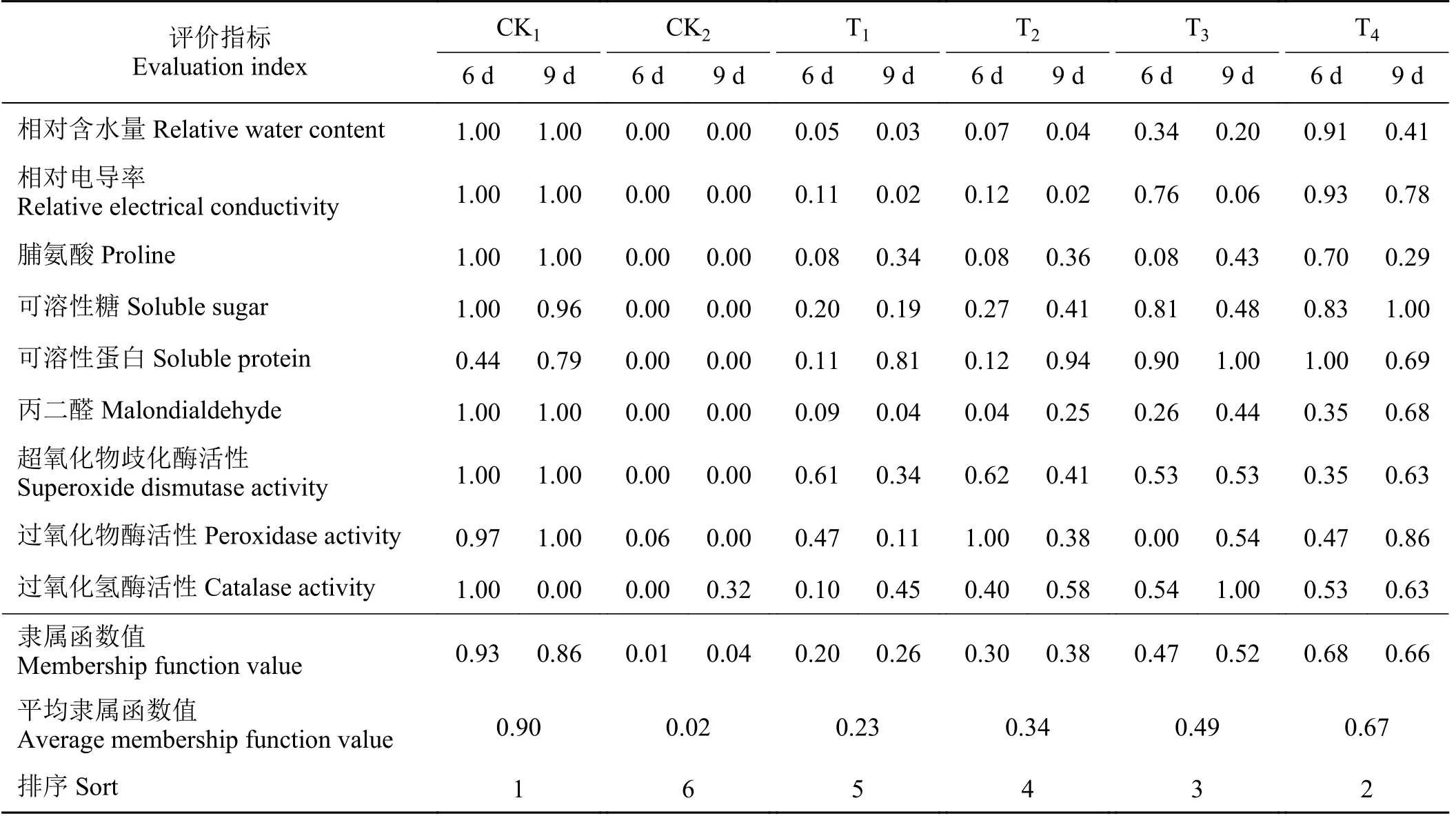
表8 不同外源激素处理下黑麦草的生理指标的隶属函数值及抗旱性排序Table 8 Membership function values and drought resistance ranking of physiological indexes of ryegrass under different exogenous plant hormone treatments
3 讨论
3.1 不同外源激素对干旱胁迫下黑麦草生理特性的影响
植物叶片相对含水量是重要的水分状况指标,随着干旱胁迫程度的加深,叶片相对含水量表现出下降的趋势[23]。本研究中随着干旱天数的增加,黑麦草叶片相对含水量下降,而施加外源植物激素后,可有效提高黑麦草叶片的相对含水量。外源施加GA3表现最好。干旱胁迫会破坏细胞膜的稳定性,导致细胞内电解质外渗,相对电导率升高。相对电导率可用来衡量植物受伤害的程度[24]。本研究中黑麦草叶片相对电导率随干旱胁迫天数的增加逐渐升高。喷施外源ABA 和GA3可显著降低黑麦草相对电导率,表明喷施外源植物激素缓解了干旱胁迫下黑麦草细胞膜受到的伤害。这与缐旭林[25]的研究结果一致。干旱胁迫下植物体内活性氧(reactive oxygen,ROS) 含量增加,膜脂发生氧化损伤,导致细胞膜结构和稳定性遭到破坏。MDA 含量的高低可以反映膜脂过氧化作用的强弱和膜结构受伤害的程度[26]。刘文瑜等[1]研究发现,4 个不同品种的藜麦(Chenopodium quinoa)幼苗的MDA 含量随着干旱胁迫程度的增加而逐渐增加,这与本研究结果一致。本研究中随着干旱胁迫天数的增加,黑麦草叶片MDA 含量逐渐增加。相关研究表明在干旱胁迫下施用不同浓度的EBR 可使得棉花(Gossypium hirsutum)成苗叶片中的MDA 含量显著降低,缓解干旱胁迫对细胞的损伤[24]。本研究中在干旱胁迫第9 天,T3和T4处理下黑麦草叶片MDA 含量显著降低(P< 0.05),表明外源施加ABA 和GA3可有效缓解干旱胁迫对黑麦草细胞的损伤。
在干旱胁迫下,植物体渗透调节物质含量增加。本研究中干旱胁迫下,黑麦草Pro、SS 含量增加,SP 含量先增加后减少。逆境下植物渗透调节物质含量的高低与植物渗透调节能力的大小密切相关[27]。在干旱胁迫第9 天,外源施加植物激素黑麦草Pro、SS 含量降低,SP 含量增加。表明外源激素缓解了干旱胁迫对黑麦草渗透调节的影响,其中外源GA3处理下效果最好。
干旱胁迫下,植物体内ROS 含量增加可作为信号物质促进植物体内抗氧化酶活性的增加,抗氧化酶活性的增加可达到消除活性氧维持植物体内活性氧动态平衡的目的[28]。本研究中随着干旱胁迫天数的增加,黑麦草SOD、POD 酶活性逐渐增加,CAT 酶活性先增加后减少。外源施加植物激素处理下黑麦草SOD、POD、CAT 酶活性在干旱胁迫第0天与CK2相比表现为增加,在干旱胁迫第9 天SOD、POD 酶活性表现为减少,CAT 活性表现为一定程度增加。许多研究表明,施加外源激素后可通过提高植物的抗氧化酶活性来缓解干旱胁迫对植物的伤害[5]。这与本研究结果有一定的差异,可能受干旱胁迫较轻,活性氧产生较少,使得黑麦草产生较少的抗氧化酶,这表明外源激素可提高黑麦草的抗旱能力,缓解干旱胁迫对黑麦草的伤害。
3.2 不同外源激素对干旱胁迫下黑麦草叶片内源激素含量的影响
植物在遭受干旱胁迫时,会引起体内内源激素含量和活性的改变,从而调控植物的生理生化过程和同化产物的运输和分配[29]。ZR 作为高等植物体内一种主要的游离态细胞分裂素,其作用主要是促进细胞的分裂和生长。LI 等[30]研究表明,细胞分裂素参与了植物对干旱胁迫的响应。本研究中,在干旱胁迫第6 天,CK2处理比CK1处理ZR 含量显著增加(P< 0.05),而外源施加植物激素后,黑麦草ZR 含量都呈下降的趋势。表明黑麦草在干旱胁迫下可能通过增加ZR 含量以延缓叶片衰老。这与张世英等[31]对中华金叶榆(Ulmus pumila)盆栽苗的研究结果一致。而外源植物激素可能通过降低ZR 含量来减慢黑麦草的生长速度,减少对水分的需求,以更好地适应干旱[32]。ABA 作为一种胁迫应激激素,参与多种非生物胁迫[33]。研究表明干旱胁迫下,植物体内ABA 含量显著升高[34],这与本研究结果一致。T1、T2、T3、T4处理下黑麦草叶片中内源ABA含量显著降低(P< 0.05),表明外源施加植物激素有效缓解了干旱胁迫下黑麦草植株的衰老,T4处理下效果最好。内源GA3不仅能促进细胞伸长而生长,还能改变植物对水分的利用、促进气孔关闭减少蒸腾。有研究表明,内源GA 的含量会随着干旱胁迫的增加而降低[35],但也有研究表明干旱胁迫可增加植物内源GA3含量[36]。本研究中干旱胁迫下内源GA3含量显著增加(P< 0.05),T1、T2、T4处理下黑麦草内源GA3含量增加,表明外源施加植物激素可通过增加内源GA3含量来提高黑麦草的抗旱能力。干旱胁迫对植物体内源IAA 含量的影响研究结果各异。大多研究表明干旱胁迫下,植物内源IAA 含量降低[35-36];也有研究表明干旱胁迫下内源IAA 含量增加[37]。本研究中干旱胁迫下黑麦草内源IAA 含量增加,而外源激素处理下除T2外黑麦草内源IAA 含量降低。Cox 等[38]的研究表明IAA 含量的下降有利于植物气孔的关闭,因此在干旱胁迫下IAA含量降低有利于植物更好的适应干旱胁迫。
3.3 不同外源激素对干旱胁迫下黑麦草抗旱性的影响
本研究用隶属函数法对不同外源激素处理下黑麦草抗旱性大小的排序结果为CK1> T4> T3>T2> T1> CK2,表明外源喷施植物激素有效提高了黑麦草的抗旱能力,缓解了干旱胁迫对黑麦草的伤害。效果最好的是外源GA3处理,其总的隶属函数值在4 种外源激素处理中最高。干旱胁迫下,黑麦草相对含水量降低,相对电导率、Pro 含量、SS含量、MDA 含量和SOD、POD 酶活性增加,SP 含量和CAT 酶活性先增加后减少, 黑麦草内源激素ZR、GA3、ABA、 IAA 含量都增加。干旱胁迫下,外源施加激素可通过提高黑麦草相对含水量,降低黑麦草相对电导率和MDA 含量来维持细胞膜结构稳定性,并通过调节体内渗透调节物质含量、抗氧化酶活性以及内源激素含量来更好地适应干旱胁迫。
4 结论
自然干旱胁迫下,外源施加激素处理后都可在一定程度上效缓解干旱胁迫对黑麦草的伤害,其中效果最好的是200 mg·L-1GA3处理。外源GA3处理主要通过提高黑麦草的相对含水量,降低叶片相对电导率、MDA 含量,增加内源GA3含量以及降低内源ZR、ABA、IAA 含量来提高黑麦草的抗旱性。本研究结果可为揭示黑麦草响应干旱胁迫的机理及促进其抗旱栽培提供理论与实践参考。
