遮阴条件下两种短命植物的叶绿素荧光特性及适应性差异
2024-03-28付慧蓉景玉婷赵恒明张城铭刘红玲
付慧蓉,李 嫚,景玉婷,赵恒明,伍 勇,张城铭,刘红玲,庄 丽
(1.石河子大学生命科学学院, 新疆 石河子 832000;2.成都师范学院化学与生命科学学院 / 特色园艺生物资源开发与利用四川省高校重点实验室, 四川 成都 611130)
光照作为植物光合作用的主要能源,是影响植物生理生态的重要环境因子,过强光照会引起植物的光抑制,造成光损伤,而过弱光强会使其养分供应不足,限制光合速率,导致植物萎蔫或死亡。近年来,极端气候事件频繁发生,荒漠太阳辐射增强,植物生理功能和群落稳定性受到严重威胁[1]。因此,研究荒漠植物对光照的光合适应性,提高植物适应能力,对荒漠化防治及生态环境建设有着重要意义。
古尔班通古特沙漠是中国第二大沙漠,植被覆盖率约50%,生长有多年生灌木及一年生草本植物,其中,短命植物占植物总数的37.1%,绿色产量占群落总产量的60%以上,为春季主要草本植物类群[2]。它们常利用冬季降雪融水于每年3 月-5 月迅速完成生命周期[3],整个生命期内均具有较高的光能利用率。然而,此特性极易受水分、温度和光照等外界环境的影响。叶绿素荧光技术作为植物光合作用的无损探针,其参数变化能够准确反映植物在各种环境下光能吸收、利用、传递及耗散过程[4]。在干旱高温复合胁迫下,油菜(Brassica campestris)的初始荧光产量(Fo)和最大荧光产量(Fm)增加,最大光合效率(Fv/Fm) 和电子传递速率降低,光系统Ⅱ反应中心受损,光合能力降低[5];黄条金刚竹(Pleioblastus kongosanensis)随着干旱程度增加光合效率、非光化学淬灭系数(NPQ)和环式电子传递流量子产量Y(CEF)呈先升后降趋势,在轻度干旱时启动了光保护机制以消耗过剩光能,而重度干旱时光合系统结构受损,光保护能力减弱[6];采用叶绿素荧光技术对13 种刚竹(Phyllostachys viridis)品种进行光合效率的测定,发现其中6 种光合效率较高,生长良好,是种植的优选竹种[7]。叶绿素荧光技术在评估植物受胁迫程度、选育优良植物品种中发挥了作用。然而,由于短命植物长期生长于荒漠环境中,其叶绿素荧光方面研究较少,对其光照的适应性尚不清楚。植物光合日变化依据光照强度的增加或减弱表现出“单峰型”和“双峰型”变化趋势,其“双峰型”植物在午间(12:00-15:00)光强最高时会发生明显的光抑制[8]。短命植物东方旱麦草(Eremopyrum orientale)和狭果鹤虱(Lappula semiglabra)的光合日变化均呈“双峰型”曲线,并存在明显午间光抑制[9],然而,卷果涩芥(Malcolmia scorpioides)和硬萼软紫草(Arnebia decumbens)作为古尔班通古特沙漠南缘的优势短命植物[10],是否也存在相同的午间光抑制,需进一步研究证实。
因此,本研究以卷果涩芥和硬萼软紫草为研究对象,通过对自然环境和遮阴低光条件下两种短命植物叶绿素荧光参数日变化、叶绿素含量和生物量的测定和分析,探索其叶绿素荧光日动态变化规律,找出不同短命植物物种光合适应性差异,揭示其光合生理机制,进而为短命植物资源保护和荒漠植被恢复提供理论依据和数据支撑。
1 材料与方法
1.1 研究区概况
研究区位于古尔班通古特沙漠南缘莫索湾研究站(86.06° E,45.03° N,海拔364~359 m),该地夏季干旱少雨,冬季寒冬少雪,是典型的温带大陆性干旱半干旱气候,光热资源丰富,年平均日照时间达3 100~3 200 h,平均温度为4~6 ℃,昼夜温差极大,年均降水量为160 mm,其中本研究进行期内(2022 年5 月1 日-15 日)的白天平均温度为31 ℃,夜间平均温度为16.4 ℃,降水量为1.8 mm,平均光照强度日变化呈抛物线型(图1)。土壤类型以风沙土和灰漠土为主。植物群落由梭梭(Haloxylon ammodendron)、沙拐枣(Calligonum mongolicum)等灌木和尖喙牻牛儿苗(Erodium oxyrhinchum)、沙冬青(Ammopiptanthus mongolicus)等一年生植物组成。
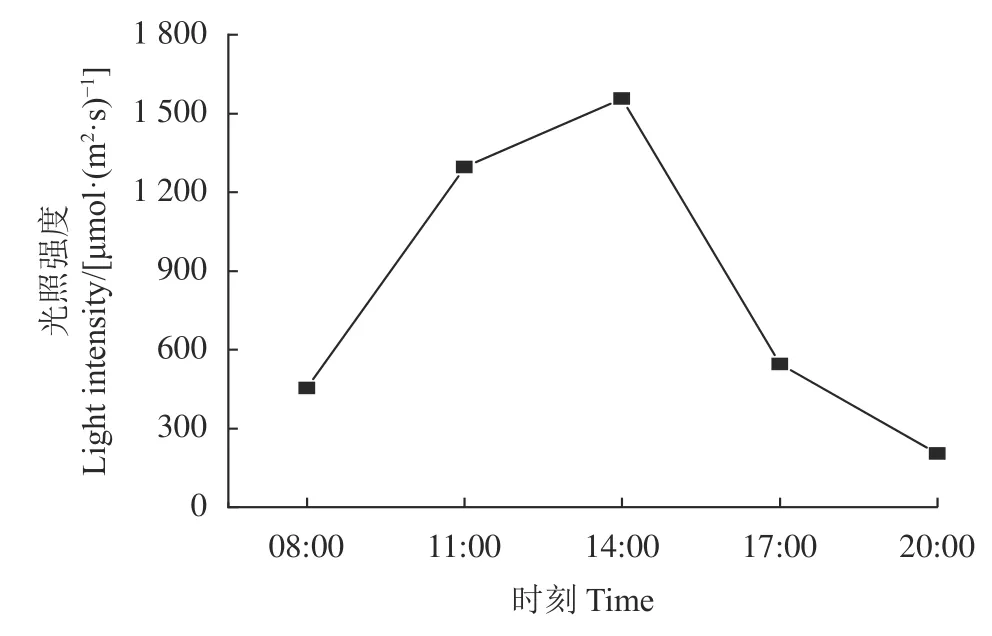
图1 试验期内(5 月1 日-15 日)莫索湾地区平均光照强度日变化Figure 1 Daily variation in average light intensity in the Mosuowan area during the test period (May 1 to 15)
1.2 试验设计
于2022 年5 月在莫索湾研究站附近寻找一块卷果涩芥和硬萼软紫草分布较为密集的样地,密度分别为39.7%和30.6%,将此样地分为4 块5 m × 5 m的小样地,分别对应试验设置的4 个遮阴时间梯度(0、1、7、14 d)。对应0 d 的样地无需进行任何处理,其他3 块样地均用遮阴率为75% 的遮阴网于2022 年5 月1 日进行遮阴,遮阴高度为1.5 m,遮阴完成后用手持海博特光谱仪测定光强,确保遮阴率达到遮阴要求。其后在相应天数对应的样方中选择生长良好、叶片完整的植株测定两种短命植物的叶绿素荧光参数日变化,测定时间为0 d (5 月1 日)、1 d(5 月2 日)、7 d (5 月8 日)和14 d (5 月15 日)。这4 d的测定均从08:00 至20:00,每3 h 测定一次,共5 次,每次进行3 个重复(n= 3),每一重复均选取不同植株中部、完全展开的叶片进行测定,并在测定完成后用红绳进行标记。待14 d 的数据测定完成后,在14 d 对应的遮阴样地和附近的未遮阴样地中摘取叶片用于叶绿素含量的测定,并挖取完整的卷果涩芥和硬萼软紫草植株,各25 株,制作为3 个重复,用于生物量的测定,将未遮阴样地中的材料作为对照。
1.3 叶绿素荧光参数的测定
参考Kramer 等[11]的方法,利用双通道调制叶绿素荧光仪Dual-PAM-100 (Walz,Germany)测量PSⅠ和PSⅡ的叶绿素荧光参数。首先,将植株叶片暗适应30 min,于低光强[30 μmol·(m2·s)-1]下测定初始荧光(Fo),并施加长约300 ms 的饱和脉冲光[10 000 μmol·(m2·s)-1]测定最大荧光(Fm),其后使用远红光代替光化光照射10 s,测定饱和脉冲光下的最大P700 信号(Pm),以确定P700 的氧化还原状态。最后,打开光化光持续4~5 min,每隔20 s 给予一个持续长约300 ms 的饱和脉冲光,待慢诱导曲线平稳后,记录最大荧光信号(Fm′) 和最大P700 信号(Pm′),关闭光化光,通过计算机PAM 软件自动计算相应的叶绿素荧光参数,计算公式如下。
PSⅡ非调节性能量耗散的量子产量[quantum yield of non-regulated energy dissipation in PSⅡ,Y(NO)] =Fs/Fm;
PSⅠ供体侧限制引起的非光化学能量耗散的量子产量[quantum yield caused by PSⅠ donor-side limitation,Y(ND)] =P/Pm;
PSⅠ受体侧限制引起的非光化学能量耗散的量子产量[quantum yield caused by PSⅠ acceptor-side limitation,Y(NA)] = (Pm-Pm′)/Pm;
式中:0.84 为吸收率(叶子吸收的入射光的比例),0.5 为到达PSⅠ或PSⅡ的吸收光的比例,PPFD为光合有效辐射强度。
1.4 叶绿素含量和生物量的测量
采用紫外光光度法在遮阴14 d 时摘取适当植株叶片,剪碎,称取0.10 g,用5 mL 的80% 丙酮避光浸提48 h,使用Unic7200 型分光光度计在470、646、652 和663 nm 的波长下测定吸光度,根据Lichtenthaler 和Wellburn[12]的方法计算叶绿素含量。
将挖取的植株用蒸馏水清洗后吸干水分,称取鲜重,其后置于105 ℃的烘箱中,杀青1 h,再将温度调至80 ℃,烘至恒重,称取干重。
1.5 午间光抑制的主成分分析和逐步回归分析
由于本研究中叶绿素荧光参数数量较多且各参数日变化复杂,参数之间存在高度相关性,严重增加了数据的分析难度,因此,采用主成分分析法(principal component analysis,PCA) 将多变量数据进行降维处理,以进一步探索午间(14:00) 自然强光(对照)和遮阴低光下两种短命植物叶片光合机构和功能的变化规律,并通过逐步回归分析方法将光合活性及光损伤情况进行量化处理。
1.6 数据处理
用Excel 2019 整理数据;用SPSS 26.0进行两种短命植物叶绿素荧光参数、叶绿素含量和生物量的差异明显性分析(P< 0.05),用平均值±标准误表示结果;运用SPSS 26.0 回归分析法进行逐步回归分析;运用Origin 2021 作图和PCA 聚类分析。
2 结果
2.1 遮阴对两种短命植物叶绿素荧光参数的影响
0 d,两种短命植物Fv/Fm日变化呈“V”型(图2),在14:00-17:00 明显下降,且硬萼软紫草下降程度较卷果涩芥大;而对于Y(Ⅰ),卷果涩芥趋于平稳,硬萼软紫草呈“双峰型”,11:00 上升,14:00 明显下降,下降率达51.7%;Y(Ⅱ)和qP 日变化均呈“双峰”型,14:00 明显下降。经遮阴处理后,两种短命植物的Fv/Fm日变化幅度较小,除卷果涩芥Fv/Fm在14 d的14:00 下降9.3%外,其他时间点Fv/Fm均介于0.80~0.85;而Y(Ⅰ)、Y(Ⅱ)和qP 日变化幅度较大,1 和7 d时,两种短命植物Y(Ⅰ)、Y(Ⅱ) 和qP 日变化均呈“单峰型”,在14:00-17:00 上升至峰值,14 d 时,硬萼软紫草仍保持“单峰型”,卷果涩芥变为“双峰型”,14:00 明显下降,表明遮阴可改变两种短命植物光合效率日变化规律,在未遮阴环境的强光下,14:00 光合效率下降,而遮阴后达到峰值。进一步比较各遮阴天数14:00 的Y(Ⅰ)、Y(Ⅱ) 和qP,可发现相较于0 d,卷果涩芥Y(Ⅰ)在1 和7 d 时上升了19%和4.5%,14 d 时下降47% (图2C),Y(Ⅱ)在1 d 时上升55.7%,7 与0 d 无明显差异,14 d 下降52.0%;而硬萼软紫草,14 d 内的遮阴环境下午间Y(Ⅰ)、Y(Ⅱ)和qP 均高于0 d,表明随遮阴时间的延长两种短命植物PSⅠ和PSⅡ光合效率逐渐下降,相较于未遮阴环境(0 d),1~7 d 的遮阴使午间光合效率有了明显升高,但当遮阴至14 d 时,反而不利于卷果涩芥光合效率的提升。

图2 遮阴对两种短命植物光合效率和光化学淬灭系数的影响Figure 2 Effects of shading on photosynthetic efficiency and the photochemical quenching coefficient of two ephemeral plants
遮阴与未遮阴环境下Y(ND) 的日变化趋势存在较大差异(图3),未遮阴环境(0 d)下两种短命植物Y(ND) 在11:00 时明显下降,14:00 和17:00 上升;遮阴后,除卷果涩芥在14 d 时呈“倒M”型外,其他天数中Y(ND) 日变化均呈“V”型,14:00-17:00时降至最低,并随遮阴时间的延长而上升;且硬萼软紫草下降程度较卷果涩芥大,表明在未遮阴的午间(14:00-17:00) 强光下,由PSⅠ供体侧限制引起的量子产量增加,遮阴后午间PSⅠ供体侧不受限制,但日变化总趋势随遮阴时间的延长而上升。Y(NA) 较Y(ND) 平稳,两种短命植物Y(NA) 均表现为0 d > 1 d > 7 d 和14 d,但其中,卷果涩芥14 d的午间14:00 明显上升,较11:00 升高553.0%,其值正好与0 d 的Y(NA)相近,表明随着遮阴时间的延长由PSⅠ受体端限制引起的量子产量逐渐下降,但当遮阴时间延长至14 d 时卷果涩芥午间PSⅠ受体端限制引起的量子产量再次上升。Y(NO) 在0 d 的14:00 和卷果涩芥14 d 时的14:00 明显上升外,其他时间趋于平稳,表明在0 d 和14 d 的午间硬萼软紫草和卷果涩芥的PSⅡ非调节性能量耗散量子产量增加,植物面临光损伤。
遮阴处理使两种短命植物的Y(CEF) 不同程度的升高(图4);0 d 时,卷果涩芥Y(CEF) 11:00 和17:00 下降,14:00 明显升高,硬萼软紫草Y(CEF)除11:00 明显下降外,其他时间点较平稳;经过遮阴处理后,仅卷果涩芥在遮阴14 d 时呈“双峰型”,其他天数中Y(CEF) 日变化均呈“单峰型”,在14:00-17:00 升高至峰值,表明自然午间强光下,卷果涩芥环式电子传递速率增加,硬萼软紫草变化不明显;遮阴后,1~7 d 午间两种短命植物环式电子传递速率均增加,但遮阴延长至14 d 时卷果涩芥午间的环式电子速率下降。遮阴对NPQ 日变化影响较大,0 d时两种植物NPQ 均在午间14:00-17:00 明显升高;遮阴后,NPQ 随遮阴时间的延长而上升,日变化均呈“双峰型”,11:00 和17:00 上升至峰值,14:00 明显下降,表明在自然午间强光下,两种短命植物的热耗散效率增加,遮阴下,随着遮阴时间的延长,耗散效率逐渐下降。
2.2 遮阴对两种短命植物叶绿素含量和生物量的影响
硬萼软紫草的叶绿素含量较卷果涩芥高,生物量较低(表1)。经过遮阴后,除卷果涩芥类胡萝卜素含量和硬萼软紫草生物量无明显差异外,其他均明显下降,其中,卷果涩芥生物量下降率最高,达68.6%,叶绿素b 含量次之;硬萼软紫草叶绿素b 含量下降率达31.6%,类胡萝卜素含量下降率最低,为13.8%。进一步比较两种短命植物遮阴后叶绿素含量和生物量的下降率可发现,卷果涩芥总叶绿素含量和生物量的下降率明显高于硬萼软紫草,表明75%的遮阴率不利于两种短命植物叶绿素和生物量的积累,且卷果涩芥受遮阴影响较硬萼软紫草大。
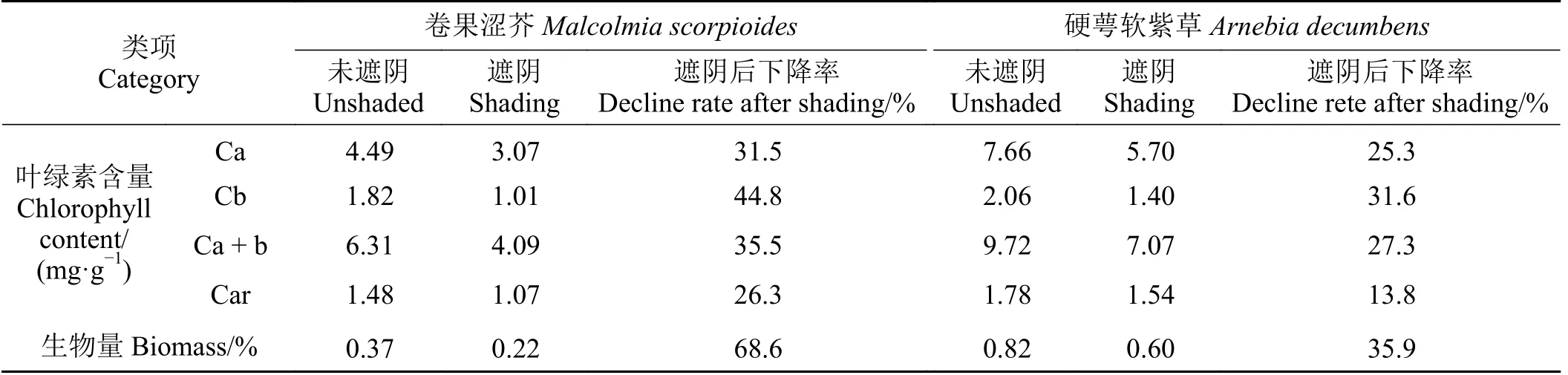
表1 遮阴对两种短命植物叶绿素含量和生物量的影响Table 1 Effects of shading on chlorophyll content and biomass of two ephemeral plants
2.3 两种短命植物午间叶绿素荧光参数的PCA聚类分析
卷果涩芥的PC1和PC2贡献率为55.4%和18.1%(图5),PC1主要分布有簇1 和簇3,包括Y(Ⅰ)、qP、rETR(Ⅱ) 和Y(ND) 等参数,簇2 分布于PC2上,包括NPQ 和Y(NA)。结合各参数的生理意义,簇1 主要代表PSⅠ和PSⅡ的光合活性,簇2 和簇3 代表热耗散效率和由PSⅡ供受体侧限制引起的量子产量。根据遮阴与未遮阴的午间(14:00) 荧光参数聚类结果可发现,遮阴组主要位于第1、3 和4 象限的簇2 和簇3 下,未遮阴组位于第1 象限的簇1 和簇2 之间,表明未遮阴环境下卷果涩芥的PSⅠ和PSⅡ活性较高,而遮阴环境使热耗散和光损伤程度升高。硬萼软紫草的PC1和PC2贡献率为75.5%和10.3%,荧光参数主要形成了两簇,且两簇均位于PC1 上,簇1 包括参数与卷果涩芥簇1 基本相同,簇2 包括Y(ND)和NPQ。另外,PC2上分布有Y(NO)和Y(NA),簇1 代表着光合活性,簇2 与Y(NO)和Y(NA)代表着热耗散效率和由PSⅠ供受体侧限制引起的量子产量。根据聚类结果可知,未遮阴组位于第2、3 象限的簇1 和PC2上的两个参数之间,而遮阴组位于第1、4 象限的簇2 下,表明硬萼软紫草与卷果涩芥相反,遮阴环境提高了硬萼软紫草光合活性,但未遮阴环境下光合活性较弱,且热耗散效率增加。另外,需特别注意NPQ 和Y(CEF)的变化,卷果涩芥Y(CEF)和NPQ 分布于簇1 和簇2 中,位于未遮阴组下,而遮阴组下仅有NPQ 分布;硬萼软紫草Y(CEF)位于遮阴组下,NPQ 位于未遮阴组下,表明Y(CEF)与两种短命植物光合活性呈正相关关系。

图5 两种短命植物午间叶绿素荧光参数的PCA 聚类分析Figure 5 PCA cluster analysis of chlorophyll fluorescence parameters of two ephemeral plants at noon
2.4 基于两种短命植物叶绿素荧光参数PCA 分析的逐步回归分析
为进一步了解遮阴对两种短命植物午间叶绿素荧光参数的影响程度,用PCA 结果中的簇1 (X1)代表光合活性,簇2 (X2)和簇3 (X3)代表热耗散效率和光损伤,以各处理下PSⅠ的实际光合效率Y(Ⅰ)为变量Y,进行逐步回归分析。结果表明,与未遮阴相比,遮阴使卷果涩芥的光合活性下降95.3%,热耗散效率提升25.0%,由PSⅠ供体侧限制引起的量子产量和PSⅡ非调节性热耗散量子产量增加95.0%;而硬萼软紫草与卷果涩芥的结果相反,遮阴使光合活性上升4.0%,热耗散效率下降18.6%,由PSⅠ受体端限制引起的量子产量和PSⅡ非调节性热耗散量子产量上升52.2% (表2)。
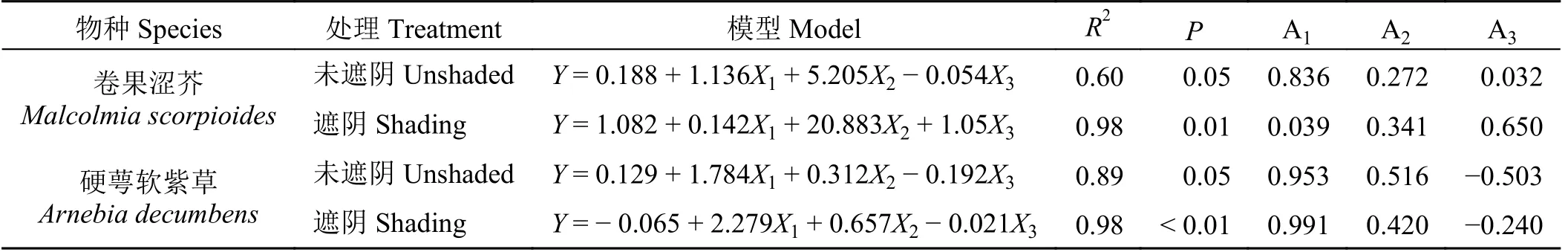
表2 基于两种短命植物叶绿素荧光参数PCA 分析的逐步回归分析Table 2 Stepwise regression analysis based on PCA analysis of chlorophyll fluorescence parameters of two ephemeral plants
3 讨论
3.1 两种短命植物存在午间光抑制
光照是植物光合作用的主要驱动力,过强或过弱会降低光能利用效率,即光抑制现象[12-14]。Fv/Fm是衡量植物光抑制的典型指标,常介于0.80~0.85,遇到胁迫环境时会明显降低[15]。在本研究中,两种短命植物Fv/Fm在午间14:00 明显下降,光合效率[Y(Ⅱ)、Y(Ⅰ)和qP]也随之降低,说明存在午间光抑制,然而,有研究表示,当植物处于午间高温、强光环境时,植物会因避免自身光合机构损伤和叶片水分过度流失而启动保护机制,即光合午休现象,此现象发生时也会表现为光合效率的降低[16]。因此,为明确本研究中两种短命植物光合效率下降的具体原因,对光合系统激发能分配的变化进行分析,发现当午间光合效率Y(Ⅱ) 和Y(Ⅰ) 下降时,Y(NA)和Y(NO)升高,说明两种短命植物吸收的光能多以非调节性热耗散的方式进行释放且光系统已受损,许多研究者将Y(NA)和Y(NO)升高作为植物光损伤指标[17],因此判断两种短命植物光合效率下降主要由光系统损伤所致。遮阴后,除卷果涩芥Fv/Fm在14 d 的午间下降外,其他天数中两种短命植物日变化均趋于平稳,且光合效率在14:00-17:00 上升至峰值,两种短命植物午间光抑制得到明显改善。这与汤正辉等[14]的研究结果一致,在遮阴环境下,凤丹白牡丹(Paeonia suffruticosa)的午间光抑制被改善,且遮阴明显提高了该物种的光合速率。不同植物对光照的需求及适应性不同,卷果涩芥和硬萼软紫草对遮阴的适应性也存在差异。随着遮阴时间的延长,两种短命植物的午间光合效率均表现为1 d >7 d > 14 d,午间光抑制的改善效果随遮阴时间的延长而逐渐减弱;而与未遮阴环境(0 d)相比,卷果涩芥仅1 和7 d 的午间光合效率高于0 d,硬萼软紫草14 d 内均高于0 d,说明遮阴时长超过7 d 时卷果涩芥的光合活性受到抑制,而硬萼软紫草14 d 内午间光抑制均被改善。
有研究者称,光抑制程度取决于光系统损伤程度和植物光保护能力[18]。在本研究中,环式电子传递流对提升光合活性起到了重要作用,1~7 d 两种短命植物午间光合活性随着Y(CEF) 的增加而提高,且光损伤参数Y(NO)和Y(NA)维持较低水平[17],14 d 时卷果涩芥午间Y(CEF) 下降,光合活性也随之下降,光损伤参数升高,说明环式电子传递流在维持光系统供受体侧电子传递速率、避免光损伤中起到了重要作用;另外,Y(CEF)与两种短命植物午间光合活性呈正相关关系、与光损伤参数呈负相关也体现了这一点。环式电子传递流的光保护作用已在众多研究中得到证实[19-20]。Huang 等[21]指出高活性的环式电子传递流可促进类囊体腔酸化,建立高的质子梯度(ΔPH),稳定放氧复合体(oxygenevolving complex, OEC)结构,耗散过余光能[22];并且可增加P700 的氧化率,调节Y(ND)和Y(NA)的平衡,保持P700 较大的电子流出速率,避免供受体侧电子积累造成光损伤[23]。
3.2 两种短命植物对遮阴环境的适应能力
根据植物对光照强度的需求,可将植物分为阳性植物、阴性植物和中性植物三大类[24]。卷果涩芥和硬萼软紫草长期生长于光照强度较高的荒漠中,因此大量研究者将其归类为阳性植物,但在本研究中两种短命植物对未遮阴强光和遮阴低光的适应性存在明显差异,未遮阴的午间强光下(0 d)卷果涩芥光合效率、电子传递速率和Y(CEF)均较高,遮阴后Y(ND)、Y(NA) 上升,光损伤程度增加95.0%。硬萼软紫草与其相反,未遮阴的午间强光下光损伤程度增加52.2%,经遮阴后,光合效率升高4.0%,热耗散能量降低18.6%,说明卷果涩芥较硬萼软紫草耐强光,硬萼软紫草比卷果涩芥耐阴。在遮阴环境下硬萼软紫草类囊体ΔPH 较高,吸收的光能多用于CO2固定和光呼吸,具有较高的光能利用率[25],而卷果涩芥对强光、高盐和干旱等环境的强抗逆性在樊丛照等[10]的研究中也被证实,这也许正是莫索湾地区卷果涩芥物种丰富度高,硬萼软紫草常生长于草甸下的原因之一[26]。另外,硬萼软紫草的强耐阴性在遮阴后两种短命植物叶绿素含量和生物量下降率中也可得到体现。叶绿素荧光参数作为叶绿素含量变化的指针,其高低反映着植物光合作用的强弱及干物质的积累情况[27]。经过遮阴后硬萼软紫草的午间光合活性明显高于卷果涩芥,并且硬萼软紫草的叶绿素含量和生物量下降率明显低于卷果涩芥,说明遮阴对午间光抑制的改善效果在维持硬萼软紫草叶绿素含量和生物量中起到了重要作用。
另外值得注意,经过遮阴后两种短命植物午间光合活性虽上升,但叶绿素含量和生物量均明显下降,Smethurst 和Shabala[28]认为Fv/Fm可作为的特征指数,反映叶绿素含量变化状态,与叶绿素含量和生物量呈正相关关系,而本研究结果与此相悖,其原因与试验处理中遮阴率太高有关。两种短命植物光合效率Y(Ⅱ)和Y(Ⅰ)的日变化趋势中可发现,75%的遮阴率虽可改善两种短命植物的午间光抑制,但除午间14:00-17:00 以外的其他时间点光合效率均明显低于自然环境,说明此遮阴率不宜作为最适遮阴率,对两种植物的整体光合情况具有不利影响,更加适当的遮阴率还需进一步研究探索。
4 结论
综上,在未遮阴环境的午间强光下两种短命植物由于PSⅠ受体侧受损和PSⅡ非调节性热耗散增加使光能利用受到限制,发生明显的光抑制;而在遮阴环境下两种短命植物的环式电子传递链被高度激活,避免了PSⅠ供受体侧电子的过度积累,使午间光抑制得到有效缓解,但其缓解效果随遮阴时间延长而逐渐减小。另外,两种短命植物的光合适应性存在明显不同,卷果涩芥在强光下具有较强的光保护能力,适于荒漠强光环境,可大面积种植扩繁;而硬萼软紫草可在遮阴环境下保持较高的类囊体腔质子梯度,具有高的光能利用率,适于弱光下生长,可采用适当遮阴的方式进行种植。
