鸟岛湿地土壤产甲烷菌群落对不同降水梯度的响应
2024-03-28陈克龙杜岩功车子涵王恒生
王 霞,陈克龙,杜岩功,章 妮,车子涵,暴 涵,王恒生
(1.青海师范大学生命科学学院, 青海 西宁 810008;2.青海省自然地理与环境过程重点实验室, 青海 西宁 810008;3.青海师范大学青藏高原地表过程与生态保育教育部重点实验室, 青海 西宁 810008;4.青海青海湖湿地生态系统国家定位观测研究站,青海 西宁 810008;5.高原科学与可持续发展研究院, 青海 西宁 810008;6.中国科学院西北高原生物研究所,青海 西宁 810000;7.合肥师范学院生命科学学院, 安徽 合肥 230000)
青藏高原平均海拔超过4 000 m,是气候变化的敏感区域,在水源供应、气候调节、生物多样性保护等诸多方面承担着生态安全屏障的重要作用[1]。其湿地总面积约为11.56 × 104km2[2],占中国湿地总面积的30%[3]。近几十年来,青藏高原温度明显升高,年降水量也在以每10 年10.2 mm 的增速上升[4]。气候变暖加之降水格局的改变,从一定程度上提高了湿地植物的固碳能力,但也促进了温室气体的排放,从而进一步影响湿地生态系统的碳源汇功能[5]。甲烷(methane,CH4) 是第二大温室气体,百年尺度内温室效应是CO2的28 倍[6]。CH4的产生与消耗是陆地碳循环过程的重要环节,其减排机制是当前科学研究的难点与热点任务[7]。自然湿地是目前已知的最大天然CH4排放源,贡献了所有天然CH4排放源的70%[8]。降水变化直接影响土壤养分有效性[9]和生物化学循环过程[10],可能导致表层及地下水位的波动,引起湿地的退化、扩张、消亡或新生[11],进而对CH4的产生与消耗造成较大的影响。已有研究发现,持续干旱导致CH4排放量减少,而降水增加则可以促进碳固存,增加CH4排放量[12]。
土壤产甲烷菌是CH4产生与湿地碳循环过程的重要功能菌群,其种类、丰度与CH4通量密切相关,通过能量代谢过程产生大量CH4[13-15],在湿地生态系统的物质循环和能量流动中发挥着重要作用[16]。此外,产甲烷菌对环境变化的反馈明显,其群落组成及多样性可用于预测土壤环境的演化过程[17]。降水通过改变土壤含水量来影响产甲烷菌活性与群落结构、微量气体产量、底物可用性、碳氮循环速率等,从而对土壤中CH4产生的速率造成影响[18-19]。青海湖鸟岛地处青藏高原东北部,是国际水鸟繁殖和迁徙的重要节点,在阻止荒漠化蔓延、维护生态安全等方面发挥着重要作用[20]。降水格局的改变,导致青海湖面积持续扩大,鸟岛区域的蛋岛、鸬鹚岛完全被湖水包围,进而导致鸟岛区域湿地生境发生变化[20]。目前,有关我国青海湖鸟岛区域湿地生境的研究相对较少,降水变化下鸟岛土壤产甲烷菌群落结构、多样性以及如何影响湿地生态系统结构和功能,都有待进一步研究。因此,本研究采用高通量测序技术,探讨降水变化对鸟岛土壤产甲烷菌多样性及群落结构的影响,以期为更深入地认识鸟岛CH4产生机制及其在湿地碳循环中发挥的作用提供基础数据。
1 材料与方法
1.1 研究区域概况
研究区位于青海省海北藏族自治州刚察县的青海湖湿地生态系统国家定位观测研究站——鸟岛站,地理位置为99°54′ E,37°04′ N,海拔3 194~3 226 m,属湖滨湿地[21]。气候类型为高原半干旱高寒气候,降水少、大风多、太阳辐射强、温度日较差显著[22]。年平均气温为-0.7 ℃,极端气温最高为28 ℃,最低为-31 ℃;年均降水量为322.7 mm,主要集中在6 月-8 月,低于年蒸发量[22]。土壤类型为干润砂质新成土[23],草地植被以青甘韭(Allium przewalskianum)、披针叶黄华(Thermopsis lanceolala)、赖草(Leymus secalinus)、藏龙蒿(Artemisia waltonii)、早熟禾(Poa annua)为主。
1.2 试验设计与方法
试验样地设在鸟岛站样地东北部(36.98° N,99.87° E),选择地势平坦、植被均匀的地段(40 m ×40 m)作为试验场地[24],设置自然对照(Nck)、增雨25%处理(NZa)、增雨50%处理(NZb)、减雨25%处理(NJa)、减雨50% 处理(NJb) 共5 个降水处理梯度,每种处理设置3 次重复,布局为完全随机分布。样地于2018 年6 月开始运行,降水装置[25](图1)通过改变透明U 型塑料板的数量来达到不同的减雨水平,同时将截留的雨水收集到卧箱内,采用滴管将卧箱内收集的雨水滴灌至增雨小区内达到相应的增雨水平。为防止地表径流,每个降水小区周围布设20 cm 的防径流带,并将铁皮深埋于地下25 cm。

图1 青海湖鸟岛降水装置Figure 1 Precipitation device used on Bird Island in Qinghai Lake
本研究选择在2020 年生长季初期(6 月上旬),进行土壤样品的采集与处理。利用土钻(直径为4.5 cm)在每个处理样方按照五点取样法收集0-10 cm 土壤样品。每个处理采集的土壤样品经过人工挑出石块、植物根系、凋落物后,混匀过2 mm筛。筛分后的土壤样品装入10 mL 的EP 管保存,分别标记为Nck1、Nck2、Nck3、NZa1、NZa2、NZa3、NZb1、NZb2、NZb3、NJa1、NJa2、NJa3、NJb1、NJb2、NJb3,置于液氮罐中,用于土壤产甲烷菌的高通量测序。
1.3 土壤DNA 提取与高通量测序
参照制造商的使用说明,利用Powersoil DNA Isolation Kit 试剂盒进行土壤样品DNA 的提取。通过NanoDrop 2000 分光光度计测量OD (光密度)值,并计算260 nm/280 nm 和260 nm/230 nm 吸光度比率以评估提取DNA 的浓度和纯度[26]。使用凝胶电泳(1%琼脂糖)检测DNA 的提取质量,将质检合格的DNA于-80 ℃保存待用[27]。
使用正向引物5′-GGTGGTGTMGGATTCACACA RTAYGCWACAGC-3′和反向引物5′-TTCATTGCRT AGTTWGGRTAGTT-3′扩增产甲烷菌mcrA 基因序列[28]。PCR 扩增由ABI GeneAmp® 9700 进行,在95 ℃预变性5 min 后,在95 ℃ 30 s、55 ℃ 30 s、72 ℃ 45 s进行30 个循环的PCR 反应,最后在72 ℃延伸10 min,获得的PCR 产物于10 ℃环境保存[29]。对PCR 产物进行纯化回收、检测定量,完成文库构建[30]。文库质检合格的扩增产物使用Illumina PE 150 平台进行测序。
测序数据上传至NCBI 数据库,登录号为SRR22075793、SRR22075794、SRR22075795、SRR22 075796、SRR22075797、SRR22075798、SRR22075799、SRR22075800、SRR22075801、SRR22075802、SRR22 075803、SRR22075804、SRR22075805、SRR22075806、SRR22075807。
1.4 数据处理与分析
采用软件FLASH v1.2.7、Trimmomatic v0.33 和UCHIME v4.2 对原始测序序列进行拼接、质控过滤和去除嵌合体后,得到高质量序列[30]。利用UCLUST序列比对工具,将高质量序列聚类得到操作分类单元(operational taxonomic units, OTU)数目以及韦恩图。在不同水平上对OTU 序列进行物种注释,得到相应的物种丰度表,使用R for Windows v4.0.3 中的ggplot 2 包绘制属分类水平下产甲烷菌群落的相对丰度图。运用Mothur v1.45.3 软件做稀释性分析,同时进行Observe OTUs 指数、Chao1 指数、Shannon-Wiener 指数、Simpson 指数等α 多样性指数的计算分析,采用SPSS 21.0 进行ANOVA 方差分析检验样品的群落多样性指数的显著性。使用Qiime 软件计算Unifrac 距离,R 包绘制PCoA 图,利用SPSS 21.0 进行ANOSIM分析来检验微生物群落组间差异是否显著。利用Galaxy 平台[31]对不同水平菌群进行线性判别分析(linear discriminant analysis effect size, LEfSe)以寻找降水处理与自然状态土壤中具有统计学差异的Biomaker[32]。
2 结果
2.1 鸟岛土壤产甲烷菌OTU 分布
利用mcrA 扩增子测序技术对Nck、NJa、NJb、NZa、NZb 土壤产甲烷菌群落进行检测,通过与Silva 数据库比对,将高质量测序序列基于97%的相似性阈值下进行聚类操作[33],共得到1 475 个可操作分类单元(OTU)。不同处理间土壤样品的OTU数目存在差异(图2),Nck、NJa、NJb、NZa、NZb 分别含1 305、1 311、1 321、1 329 和1 316 个OTU,5 组样本共有的OTU 有1 144 个,而Nck、NJa、NJb、NZa、NZb 特有OTU 数分别为11、13、18、25、21 个。
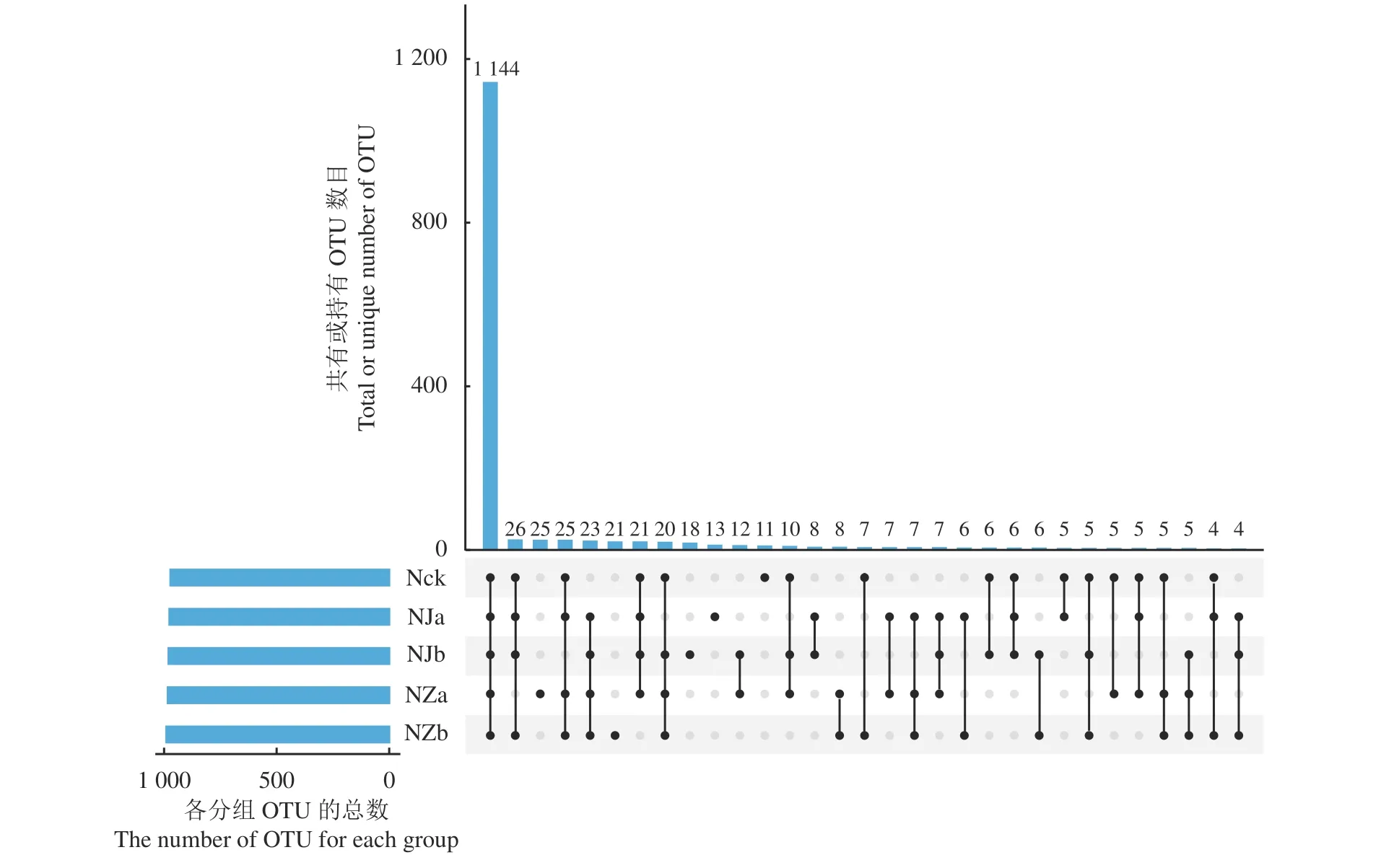
图2 不同降水梯度下鸟岛土壤产甲烷菌群落韦恩图Figure 2 Venn diagram of methanogenic community in Bird Island soil under different precipitation gradients
2.2 鸟岛土壤产甲烷菌群落α 多样性
对土壤样品中产甲烷菌的序列信息进行随机抽样,用抽取的序列数与物种多样性来构建稀释性曲线(图3A),从而说明样品测序量的准确性与合理性。稀释性曲线随样品数目的增加逐渐趋于平坦,再增加测序量也不会明显改变样本的多样性指标,表明样品序列充分,可以反映样本中绝大部分的产甲烷菌多样性信息,足以进行后续数据分析[34]。
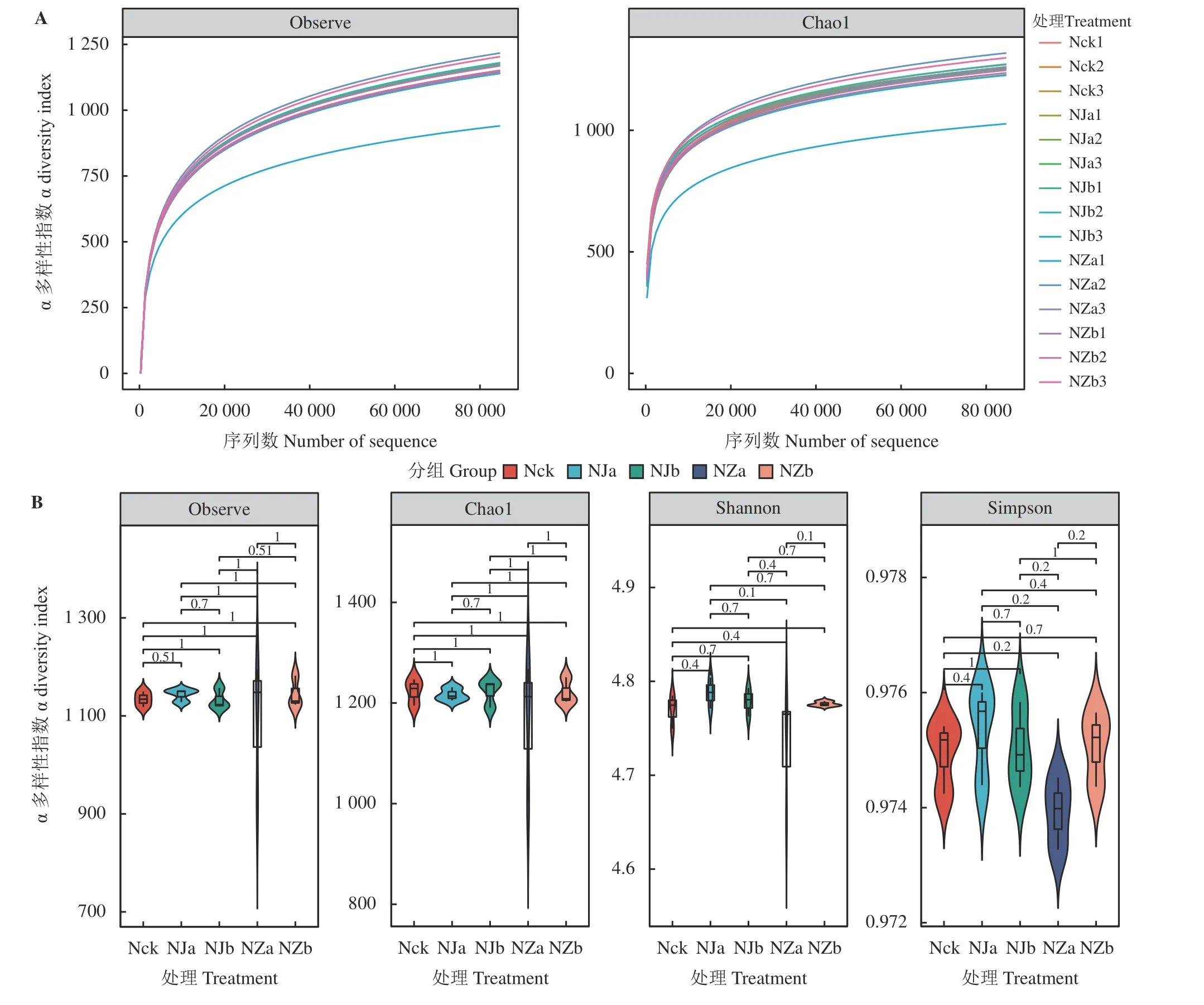
图3 鸟岛土壤产甲烷菌稀释性曲线(A)及α 多样性指数(B)Figure 3 Dilution curve (A) and alpha diversity index (B) of methanogens in Bird Island soil
α 多样性是反映微生物群落丰富度和均匀度的综合指标(图3B)。本研究采用Observe OTUs 指数和Chao1 指数来表征样品中微生物的丰富度,采用Shanno-Wiener 指数和Simpson 指数表征微生物的多样性。结果显示,产甲烷菌群落的丰富度指数表现为NZb > NZa > Nck > NJa > NJb,多样性指数表现为NJa > NJb > Nck > NZb > NZa,表明降水变化对鸟岛土壤产甲烷菌群落的α 多样性具有一定程度的影响,但其影响未达到显著水平(P> 0.05)。
2.3 鸟岛土壤产甲烷菌群落β 多样性
在群落生态学中,β 多样性是反映样本间差异的指标。本研究以检测到的所有属水平的产甲烷菌为指标,通过主坐标分析法(PCoA)对鸟岛土壤产甲烷菌群落进行分析(图4)。鸟岛土壤产甲烷菌PCoA1 的解释度为27%,PCoA2 的解释度为14%,总解释度达到41%。5 个分组中,NJb 和NZb 的样点与其余3 个组的距离较远,表明减雨50%处理和增雨50%处理对土壤产甲烷菌群落的影响可能更为显著。为进一步判断分组有效性,使用R 语言进行ANOSIM 分析,P= 0.02,说明不同分组之间差异显著(P< 0.05),达到预期分组目标。

图4 鸟岛土壤产甲烷菌群落PERMANOVA 和PCoA 分析Figure 4 PERMANOVA and PCoA analysis of soil methanogens on Bird Island
2.4 鸟岛土壤产甲烷菌群落结构组成
在门分类水平上,鸟岛土壤产甲烷菌群落均为广古菌门(Euryarchaeota,100%)。目分类水平上,甲烷微菌目(Methanomicrobiales)的相对丰度最高,为63.82%~65.74%; 其次是甲烷八叠球菌目(Methanosarcinales),相对丰度为15.29%~16.04%;甲烷杆菌目(Methanobacteriales),相对丰度为10.46%~11.01%;第七产甲烷古菌目(Methanomassiliicoccales)[35],相对丰度为5.80%~6.22%;甲烷胞菌目(Methanocellales),相对丰度为0.50%~0.61%;另有相对丰度为2.21%~2.40%的未分类菌群。
在属水平上(图5),主要微生物菌属(相对丰度 >0.1%),依次为甲烷丝菌属(Methanothrix),相对丰度为14.13%~14.85%;甲烷杆菌属(Methanobacterium),相对丰度为10.14%~10.61%;Methanomassiliicoccus属,相对丰度为4.21%~4.54%;Candidatus Methanogranum属,相对丰度为1.50%~1.70%;甲烷八叠球菌属(Methanosarcina),相对丰度为0.78%~0.81%;甲酸甲烷规则菌属(Methanoregula)[36],相对丰度为0.67%~0.73%;甲烷胞菌属(Methanocella),相对丰度为0.39%~0.49%;甲烷泡菌属(Methanofollis),相对丰度为0.12%~0.14%;Methermicoccus属,相对丰度为0.10%~0.14%。

图5 鸟岛土壤样品属水平产甲烷菌群落组成Figure 5 Community composition of methanogens at the genus level in Bird Island soil samples
2.5 鸟岛土壤产甲烷菌菌群LEfSe 分析
通过对5 种生境中不同水平产甲烷菌菌群进行LEfSe 分析,可以找到组间具有统计学差异的Biomarker[32]。共鉴定出31 个差异菌群(图6),目水平差异菌群为甲烷胞菌目,科水平差异菌群为甲烷胞菌科(Methanocellaceae)、甲烷螺菌科(Methanospirillaceae)、Candidatus Methanoperedenaceae 科,其余均为属水平及种水平菌群。属水平差异菌群数为10 个,相对丰度大于1% 的有Methanomassiliicoccus属、Candidatus Methanogranum属;种水平差异菌群数为17 个,相对丰度大于1%的Methanomassiliicoccus属未分类种和Candidatus Methanogranum属未分类种。在NZa、NZb、NJa、NJb 等降水处理下,相对丰度变化最显著的产甲烷菌菌属分别为甲烷螺菌属(Methanospirillum)、Methanomassiliicoccus属、Candidatus Methanogranum属、Methanosphaerula属。

图6 鸟岛土壤产甲烷菌LEfSe 分析Figure 6 LEfSe analysis of soil methanogens from Bird Island
2.6 鸟岛土壤产甲烷菌群落FAPROTAX 功能
将鸟岛土壤产甲烷菌群落与FAPROTAX 数据库进行比对,共鉴定出11 个功能基团(图7),主要包括甲烷生成(methanogenesis)、氢营养型甲烷化(hydrogenotrophic methanogenesis)、通过CO2与H2还原产生甲烷(methanogenesis by CO2reduction with H2)、暗氢氧化(dark hydrogen oxidation)、化能异养(chemoheterotrophy)、通过氢还原甲基化合物生成甲烷(methanogenesis by reduction of methyl compounds with H2)、甲基营养(methylotrophy) 等功能基团。上述功能基团占所测全部功能基团的75%以上,表明大部分鸟岛土壤产甲烷菌的代谢类型可能是以CO2/H2为底物的氢营养型。

图7 鸟岛土壤产甲烷菌功能预测Figure 7 Lefse analysis of soil methanogens from Bird Island
3 讨论
3.1 降水变化下鸟岛土壤的CH4 产生途径
许多研究表明,自然湿地土壤产甲烷菌的优势目为甲烷微菌目、甲烷八叠球菌目和甲烷杆菌目[37-40],本研究中,鸟岛5 种降水处理土壤中这3 类优势菌目的占比均达到产甲烷菌总丰度的90%以上,与前人的研究结果相一致。自然界中CH4的生成主要包括氢营养、乙酸营养、甲基营养3 种途径,其中以CO2/H2为底物的氢营养型产CH4途径广泛存在于自然环境中,除少量产甲烷菌无法利用氢营养型产CH4途径外,绝大部分产甲烷菌可以利用该途径生成CH4[41]。本研究中检测到的3 类优势菌目中大部分产甲烷菌属为氢营养型产甲烷菌,这与FAPROTAX数据库比对鉴定出的结果相一致,推测鸟岛土壤CH4产生途径主要为CO2+ 4H2→CH4+ 2H2O。乙酸营养途径是通过产甲烷菌裂解乙酸,将羧基氧化为CO2,甲基还原为CH4。自然环境中约70% 的生物产CH4来自于乙酸营养途径,但只有甲烷八叠球菌属和甲烷丝菌属可以通过该途径生成CH4[42],它们的相对丰度在本研究中经过不同降水处理后并未产生显著性变化。甲基营养途径有两种代谢模式,一种是H2依赖型,它利用H2为电子供体,将甲基还原为CH4[43-44];另一种是严格的甲基营养型,通过氧化甲基化合物生成CO2和还原当量,再进一步将甲基化合物中的甲基基团还原为CH4,这也是大部分甲基营养型产甲烷菌所采用的产CH4途径[45]。自然环境中只有甲烷八叠球菌目和甲烷球形菌属(Methanosphaera)可以通过甲基营养途径生成CH4[42],它们的相对丰度在本研究中经过不同降水处理后并未产生显著性变化。综上所述,鸟岛土壤CH4产生途径未在降水变化下出现明显改变,产甲烷菌在鸟岛自然土壤和降水处理土壤中主导的产CH4途径均为氢营养型途径。
3.2 降水变化对鸟岛土壤产甲烷菌群落多样性的影响
降水变化对土壤产甲烷菌的影响主要是通过引起土壤理化性质和植物生长发育过程的改变来影响产甲烷菌的多样性及其群落结构的稳定性。研究表明,高降水量区域的土壤微生物具有较高的群落多样性[46],而本研究与Dijkstra 等[47]的研究区年降水量均小于400 mm,这导致微生物受到水分限制而生长缓慢。本研究对鸟岛土壤进行增雨25%、增雨50%处理后,土壤水分状况得到改善,缺氧区域有所增加,所以土壤产甲烷菌的丰富度指数升高。同时,部分产甲烷菌由于无法适应土壤中的水势变化而死亡,从而导致产甲烷菌的多样性指数下降。杨阳等[24]研究表明,减雨降低了土壤全碳含量、全氮含量以及铵态氮含量,由此推测,减雨25%、减雨50%处理可能会降低土壤碳氮含量以及植物净初级生产力、抑制产甲烷菌生长,从而导致产甲烷菌丰富度指数下降。此外,减雨处理会降低土壤水分含量、增加氧气含量,这可能会导致革兰氏阴性菌死亡而促进适应能力更强的革兰氏阳性菌的生长繁殖,进而导致产甲烷菌的多样性指数升高[48]。整体而言,降水变化对鸟岛土壤产甲烷菌群落的α 多样性具有一定程度的影响,但其影响未达到显著性水平(P> 0.05),这与王楠楠等[49]、王誉陶等[50]的研究结果相一致,可能是由于土壤中的产甲烷菌对不同降水处理下的土壤环境和植物变化产生了一定的适应能力。
3.3 降水变化下鸟岛土壤产甲烷菌群落结构变化
Wang 等[51]的研究表明,微生物丰度和群落结构与降水变化有关。本研究对不同降水梯度的产甲烷菌菌群进行LEfSe 分析,共鉴定出31 个差异菌群,属水平差异菌群数为10 个,在增雨25%、增雨50%、减雨25%、减雨50% 等降水处理下,相对丰度变化最显著的产甲烷菌菌属分别为甲烷螺菌属(Methanospirillum) 、Methanomassiliicoccus属、Candidatus Methanogranum属、Methanosphaerula属,说明上述菌属可作为不同降水水平下鸟岛土壤产甲烷菌群落结构变化的指示物种。有研究表明,甲烷螺菌属是氢营养型产甲烷菌,细胞染色为革兰氏阴性,其对增雨25%较为敏感可能是由于产甲烷菌属于严格厌氧菌,降水增加导致土壤含水率升高,厌氧程度增大,提高了菌群活性,因此甲烷螺菌属的相对丰度增加[52-53]。Methanomassiliicoccus属和Candidatus Methanogranum属同为第七产甲烷古菌目下的菌属,缺少将CO2还原为甲基辅酶M 的完整路径,需额外添加H2才能生长,两者对降水变化的敏感程度可能是由于土壤养分有效性存在差异[54]。Methanosphaerula属也是氢营养型产甲烷菌,细胞染色为革兰氏阳性,可能是由于其细胞壁较厚,对环境变化的调整适应力强,更能适应干旱环境,因此在减雨50% 处理下相对丰度显著增加[55]。总体而言,增雨处理增加了产甲烷菌的相对丰度,减雨处理降低了菌群的相对丰度,这一结果进一步证实了鸟岛土壤产甲烷菌群落的丰度和群落结构受降水变化的影响。
4 结论
青海湖鸟岛土壤产甲烷菌群落特征对不同降水梯度的响应研究表明:1)鸟岛土壤产甲烷菌群落的优势菌目为甲烷微菌目、甲烷八叠球菌目和甲烷杆菌目;2)不同降水梯度下的鸟岛土壤均以氢营养型产甲烷菌占优势地位,由此推测产甲烷菌在鸟岛土壤中主导的产CH4途径为氢营养型;3)降水变化对土壤产甲烷菌群落多样性的影响不显著(P> 0.05);4)鸟岛土壤产甲烷菌群落结构对降水变化的响应较为敏感,尤以增雨50% 处理下差异菌群数量最多。研究鸟岛湖滨湿地土壤产甲烷菌群落结构、多样性并进行差异分析和功能预测是了解CH4产生机制的关键环节,为进一步研究降水变化背景下高寒湿地碳循环提供了数据支撑和参考依据。
